
Abstract
Abstract
A study was conducted using zebrafish as a model of hypoxic brain injury to investigate the potential neuroprotective effects of zinc (Zn2+) chelation. The accumulation of intracellular Zn2+ is a significant causal factor of the neuronal injury, and has been implicated in cell death followed by ischemic stroke. In this study, the zebrafish was placed in the hypoxia chamber with an extremely low level of dissolved oxygen (less than 0.8 mg/L), which is similar to the conditions in a complete global ischemic stroke. Approximately 50% of zebrafish died after a short period (≈11 min) of hypoxic treatment, suggesting that this is a responsive model system for use in evaluating treatments for hypoxic brain damage. The application of DEDTC reduced intracellular Zn2+ accumulation and produced a concentration-dependent effect by increasing the survival rate of zebrafish. Zn2+ chelation also enhanced zebrafish tolerance for hypoxia. When the brain damages were evaluated with TTC staining, the zebrafish that were treated with DEDTC in hypoxic treatment yielded the improvement of TTC staining that was similar to the healthy zebrafish brain. The results support that rising intracellular Zn2+ plays a critical role in the neuronal damages, and demonstrate the protective effects of Zn2+ chelation in hypoxic-ischemic brain injury in zebrafish.
Introduction
The accumulation of endogenous zinc (Zn2+) in neurons following ischemic stroke is well recognized.9–12 The staining of affected brain regions with Zn2+ indicators and cell viability probes revealed a striking correlation or co-localization between Zn2+ accumulation in neurons and cell death.13–15 While Zn2+ is found abundant in a great number of proteins and organelles (such as synaptic vesicles), Zn2+ homeostasis is tightly controlled so that the intracellular and extracellular free Zn2+ concentration is considerably low comparing to other divalent ions. 12 Therefore, an excess of Zn2+ accumulation is a critical and detrimental factor to the neuron after stroke. The treatment of rodent models of cerebral ischemia with a Zn2+ chelator reduces both infarct volume or neuronal death, and Zn2+ accumulation in affected neurons.16–19 Additionally, the idea of endogenous Zn2+ toxicity as a contributing mechanism has been investigated and shown to be valid in other injury models, including epilepsy, brain trauma, and Alzheimer's diseases.9,10
In the present study, we used zebrafish as a model of hypoxic brain injury, to study the potential protective effects of Zn2+ chelation in the brain injury. The zebrafish was placed in the hypoxia chamber with an extremely low level of dissolved oxygen (DO) (less than 0.6–0.8 mg/L), which is similar to the conditions in a complete global ischemic stroke. Zebrafish circulation is a single circuit: heart–gills–brain (body)–heart. Zebrafish can easily take dissolved chemicals through its gills merely by breathing. Therefore, chemicals such as a chelator are typically added to the aquatic environment, allowing for easy experimental manipulation and observation. Diethyldithiocarbamate (DEDTC) is a membrane-permeable Zn2+ chelator. It has been shown to chelate histochemical reactive or free Zn2+ in the brain effectively.20,21 The present study shows that, after the hypoxic treatment, the application of DEDTC produced a concentration-dependent effect by increasing zebrafish survival rate. The application of DEDTC also enhanced the hypoxia tolerance of zebrafish. Quantified analysis revealed that DEDTC improved the neuronal viability after hypoxic treatment. Our results suggest that Zn2+ chelation has protective effects in zebrafish that are subject to hypoxic brain injury.
Materials and Methods
Animals
All work in this study was conducted according to Ohio University Institutional Animal Care and Use Committee (IACUC) guidelines. Adult zebrafish (Danio rerio) were purchased from a local supplier and kept under 12-h light/12-h dark cycle with tank water which contained half deionized water plus half dechlorinated water at room temperature. The dechlorinated water was made by fresh tap water with 10–15 mg/L sodium thiosulfate. 22 Zebrafish were fed daily by commercial food flake. The average size of zebrafish used in this study was 3.82±0.36 cm (mean±SD).
Hypoxic treatment
As previously described, 8 the zebrafish was individually treated with hypoxia in the hypoxic chamber until it reached the endpoint. The endpoint was defined as zebrafish staying motionless for 1 minute. In the experiment of hypoxia tolerance, sodium diethyldithiocarbamate trihydrate (DEDTC) was added in the hypoxic chamber. After they reached the endpoint, the zebrafish was transferred to tank water to recover. The process of recovery was recorded by a camera (Nikon D300s) for 2 hours. Software A4 image video converter was used to convert the video into image sequences.
TTC staining and measurement
The 2,3,5-triphenyltetrazolium chloride (TTC) staining method was adopted and modified from Preston et al. 23 The staining was performed 24 hours after the hypoxic treatment. Slices were taken as 1 mm thickness, from the middle region of the optic lobe, which was 300 μm away from the beginning of the optic lobe, and were placed in TTC solution (2% by weight) in darkness. For staining only, the incubation time was 40 minutes. Slices were then placed in 10% formalin overnight and images taken on the next day. For staining and then extracting, the incubation time was extended to 90 min. After staining, the TTC solution was discarded and slices were gently rinsed by 2 drops of DMSO/ethanol (1:1) solution, and then placed individually in a cuvet with 1 mL DMSO/ethanol (1:1) solution in darkness for extraction overnight. A spectrophotometer was used to measure absorbance of each cuvet. Slices were taken out from cuvets before absorbance values were tested. The area of the brain slices was measured by software Motic plus 3.0. The hypoxic treatment in experiments of the TTC staining was 8 min.
Fluorescent Zn2+ imaging and brain slice preparation
Adult zebrafish were anesthetized with MS-222 and decapitated. The brain was removed and placed in ice-cold artificial cerebral spinal fluid (ACSF) and cut coronally into one 200 μm thick slice by a Vibratome at 300 μm away from the beginning of the optic lobe. ACSF was made by the following (in mM): 121 NaCl, 1.75 KCl, 5 MgCl2, 1.25 KH2PO4, 26 NaHCO3, and 10 glucose, gassed with 95%O2/5% CO2. 14 Slices were incubated with 5 μM membrane impermeable Newport Green, a fluorescent Zn2+ indicator, for 5 min, and then rinsed with ASCF twice before taking images.
Chemicals and reagents
Sodium thiosulfate, 2,3,5-triphenyltetrazolium chloride (TTC), and sodium diethyldithiocarbamate trihydrate (DEDTC) were purchased from Sigma Aldrich (St Louis, MO). Newport Green DCF (cell impermeant) was purchased from Invitrogen (Life Technologies Corporation, Grand Island, NY).
Statistical analysis
Values of standard deviation were calculated by Microsoft Excel. Statistical analysis of percentage of death under different DEDTC concentrations was performed using Fisher's exact test under R program. Statistical analysis of hypoxia tolerance and TTC measurement were performed using one-tailed Student t test under Microsoft Excel. P<0.05 was considered significant.
Results
As described previously, 8 the zebrafish were transferred individually into the hypoxia chamber when DO was below 0.8 mg/L (Fig. 1B). The zebrafish was hypoxia sensitive as erratic behaviors, beginning with agitated-like fast swimming, were readily observed usually within 1 min after being placed in the chamber. In a few minutes, the uncoordinated swimming movement became obvious and recurrent, followed by loss of equilibrium, with periods of quiescence, and quickly falling to the bottom of the hypoxia chamber. The endpoint of the time in the hypoxia chamber (when the zebrafish was taken to the recovery beaker) was determined as when zebrafish was lying motionless on one side at the bottom of the chamber for a complete 1 minute (except for continual opercular movement). Once this endpoint was reached, the zebrafish was then transferred to the recovery beaker for recovery and behavioral observation. The mean time to endpoint (TTE) of zebrafish in hypoxia chamber was 679.52±90 seconds. There was no difference in average TTE between surviving and dead fish. Essentially, about 52% of zebrafish died and 48% survived following the hypoxic treatment, suggesting that this was a responsive animal model system for use in evaluating pharmacological interventions for the hypoxic brain injury. If the zebrafish was allowed to lie motionless at the bottom for continuous 2 min before transferring to recovery beaker, it caused the 100% death rate.
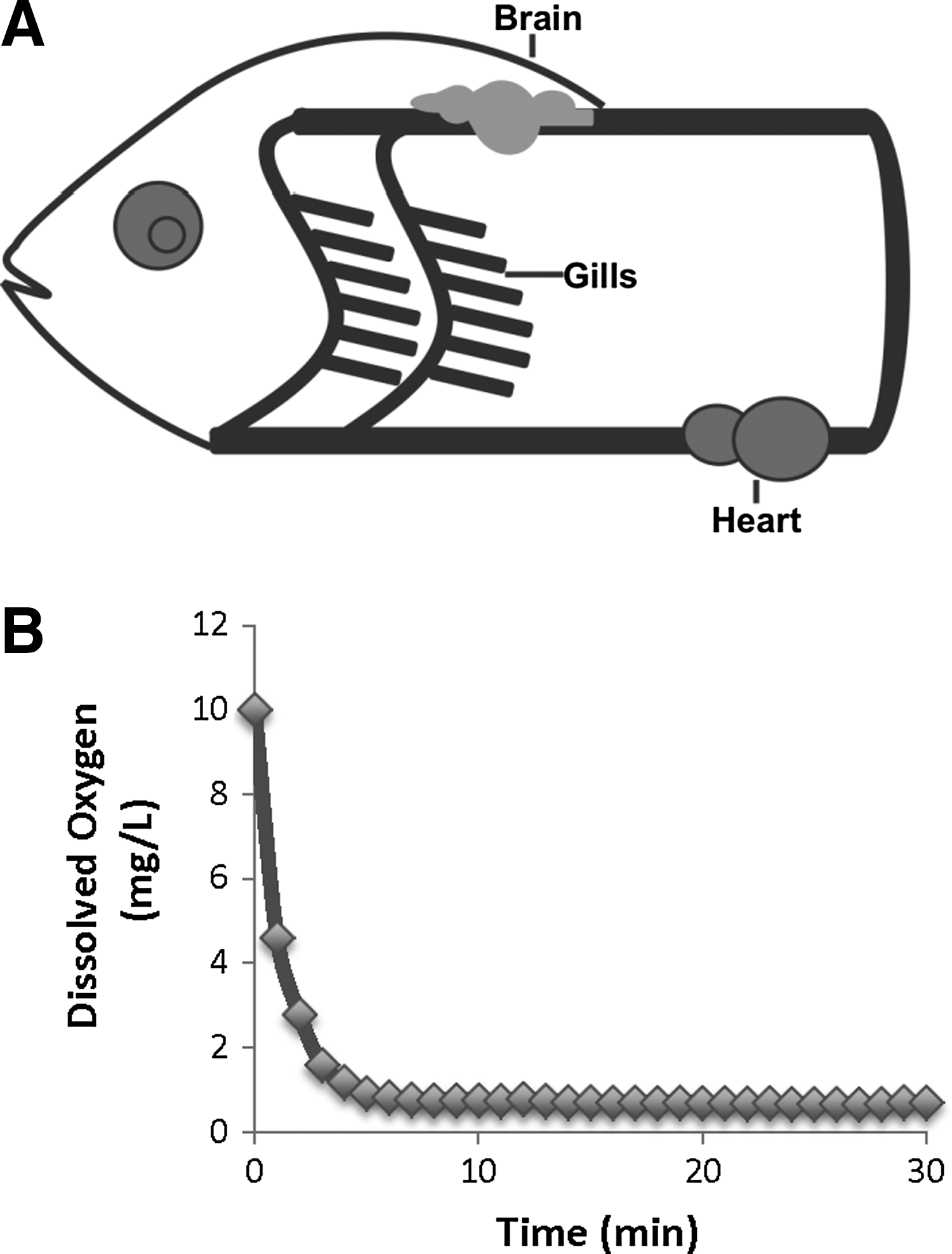
Zebrafish hypoxic model.
Zinc chelator improved the zebrafish survival after hypoxic treatment
In the control experiments, Zn2+ chelator DEDTC, when it was added in the water bathing normal zebrafish, did not alter zebrafish swimming behavior. The application of DEDTC increased survival rates of hypoxic-treated zebrafish in a concentration-dependent manner (Fig. 2). In this set of tests, DEDTC was included in the recovery chamber (containing tank water). The maximum concentration of DEDTC applied was 250 μM, which yielded 83% survival rate of hypoxic-treated zebrafish. This improved survival rate was significantly higher than that in controls (48%) (Fig. 2). Hypoxic treatment could induce a significant increase in intracellular Zn2+ and neuronal injury/death.13,14,24 The staining of affected brain regions with Zn2+ indicators and cell viability probes revealed a striking correlation or co-localization between Zn2+ accumulation in neurons and cell death.13–15 In this experiment, fluorescent Zn2+ indicator Newport Green was used to measure Zn2+ accumulation in hypoxic-damaged cells. In the hypoxia-treated zebrafish, the increases in intracellular Zn2+ were detected throughout the brain regions (Fig. 3). When DEDTC was applied during the zebrafish recovery following hypoxic treatment, the fluorescence intensity of Zn2+ staining was significantly reduced in the zebrafish brain.

DEDTC increased the survival rates. The concentration-dependent improvement of survival rate happened in DEDTC-treated zebrafish. DEDTC concentration range of 5–250 μM was tested. DEDTC was presented in recovery chamber. The numbers of animals tested: 31 zebrafish in control (0 μM DEDTC), 14 in 5 μM, 26 in 50 μM, 14 in 100 μM, and 24 in 250 μM DEDTC treatment. *p<0.05, indicating the significant difference between the control and the zebrafish treated in 250 μM DEDTC.
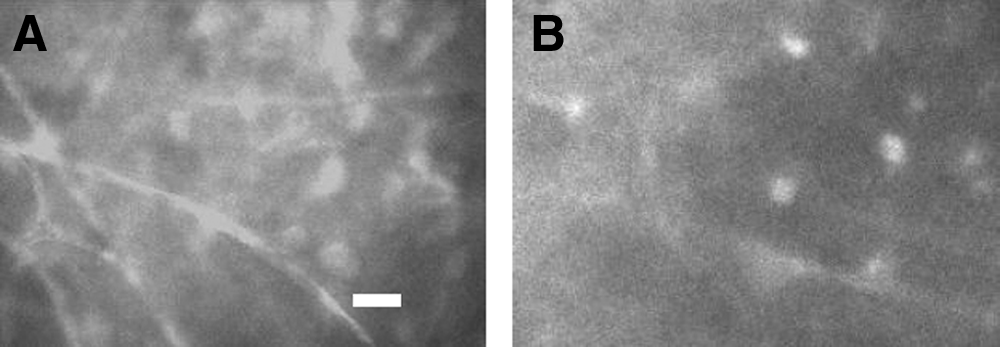
DEDTC decreased intracellular Zn2+ accumulation. The images are sections of the optic lobe of the zebrafish brain stained with a fluorescent Zn2+ indicator, Newport Green.
We recorded the movements and behaviors of the tested zebrafish once they were transferred to the recovery beaker. Through recovery following hypoxic treatment, all surviving zebrafish in either control or DEDTC application experienced initial twitching, followed by unbalanced or erratic movements before balanced movement was regained. Unbalanced movement was defined as swimming associated with erratic behaviors such as circling, rotating, and upright swimming. 8 Circling and rotating were both observed on the same zebrafish, but did not necessarily have to happen at the same time. Sometimes, zebrafish circled first, proceed by a couple of seconds of rotating, and then they were circling again. The surviving fish began to show balanced movement after about 15 min of recovery, which was when the zebrafish regained normal and coordinated swimming, indicating sufficient recovery in their swimming ability.
DEDTC enhanced the hypoxia tolerance in zebrafish
The effect of DEDTC on hypoxia tolerance in zebrafish was tested by applying DEDTC to the hypoxia chamber (not recovery chamber in the experiments described above). The hypoxia tolerance or tolerance time was defined as the period from the initiation of zebrafish hypoxia to the endpoint (or the time to endpoint, TTE) during the hypoxic treatment of each zebrafish (Fig. 4). The presence of DEDTC during the hypoxic treatment significantly increased TTE of zebrafish (TTE
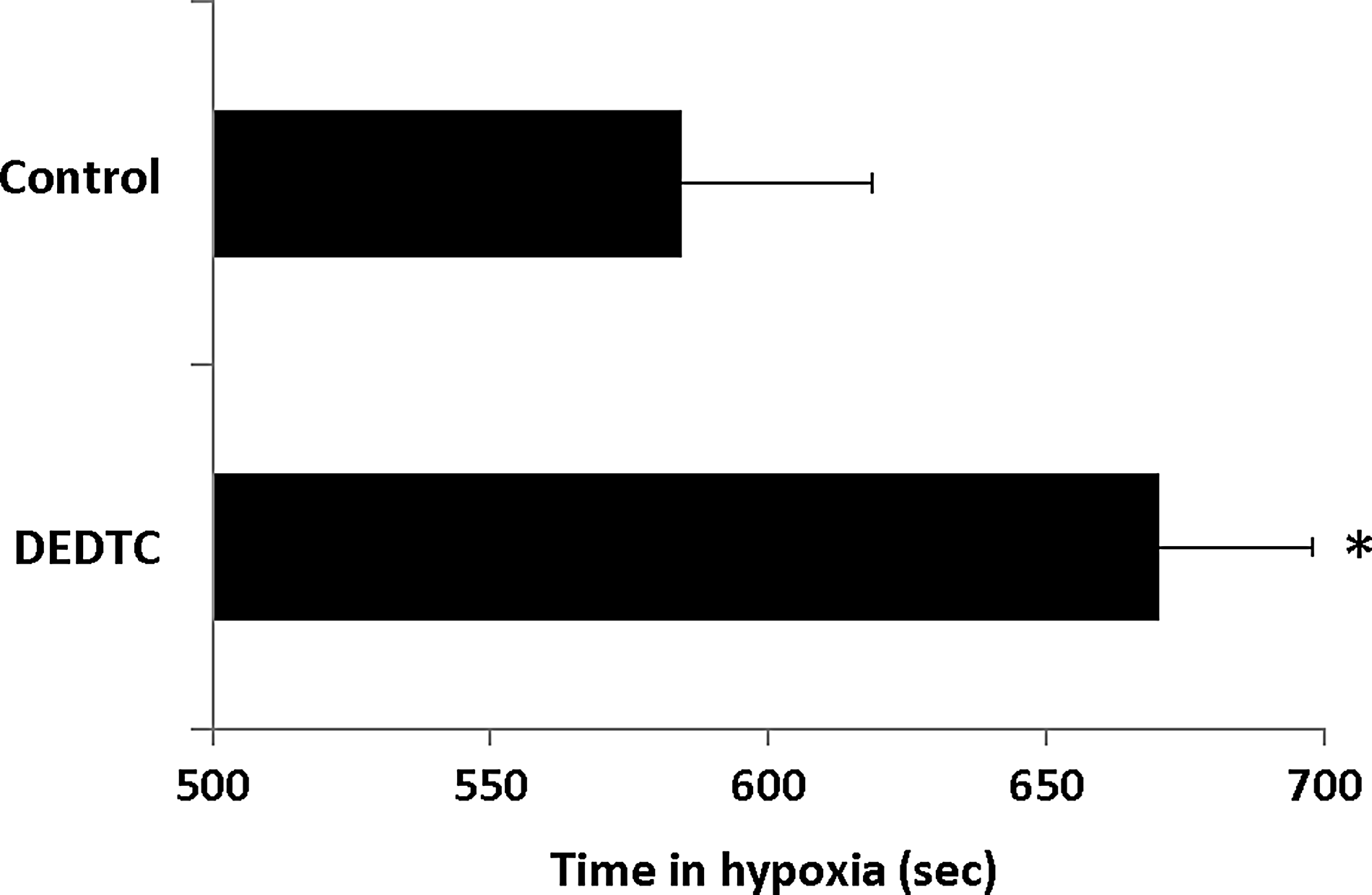
DEDTC enhanced hypoxia tolerance in zebrafish. The bar chart shows TTE (time to endpoint) of zebrafish in hypoxic treatment. DEDTC treatments (50 μM) significantly increased TTE when comparing to control (no DEDTC application). Mean±SD, N=9; *p<0.05.
DEDTC improved TTC staining
The zebrafish brain damage by hypoxic treatment was evaluated post-mortem by TTC (2,3,5-triphenyltetrazolium chloride) staining, which is the widely used method to measure hypoxic-ischemic brain damages. 25 The brains of healthy zebrafish were stained deep red (Fig. 5, also see Ref. 8). In the zebrafish that was treated with hypoxia, bilateral, moderate to complete TTC decoloration or demarcation of the infarct (pale or unstained areas versus deep red colored brain tissue) was clearly visible in the tectum of optic lobes. In some zebrafish, TTC decoloration expanded to the deep structure of the optic lobe. DEDTC when applied in hypoxia chamber significantly improved TTC staining in zebrafish (Fig. 5A). To quantify TTC staining and the brain damage, the TTC stain in the zebrafish brain was extracted and the absorbance of the extraction was measured using spectrophotometric assay following standard procedures. There were significant increases in TTC absorbance in the DEDTC-treated group, indicating more TTC stained tissue, which was 1.373±0.163 mm^-3 (mean±SD) in comparing to 0.829±0.211 mm^-3 (mean±SD) in the hypoxia group without DEDTC treatment. Also, DEDTC was proved not chemically react with TTC to increase the TTC staining by comparing the healthy group (shown as “control” in Fig. 5B) with the healthy plus DEDTC-treated group (shown as “DEDTC control” in Fig. 5B). This result further supported that the DEDTC application reduced the hypoxic brain damage.

Zebrafish brain injury detected by TTC staining.
Discussion
The major finding of this study is that Zn2+ chelation with DEDTC increases zebrafish survival rate after hypoxic treatment and enhances the tolerance of zebrafish for hypoxia. The results support previous observations that rising intracellular Zn2+ during hypoxic-ischemic treatment plays a critical role in neuronal damage and brain function.10,26 Reducing intracellular Zn2+ significantly increased neuronal survival.11,14,19 Zn2+ has been found to be released from metalloproteins and other storages during oxidative stress.27–29 Zn2+ itself is a strong inducer of oxidative stress by promoting mitochondrial and extra-mitochondrial production of reactive oxygen species, and promoting neuronal death.28,30 The application of DEDTC in the present study reduced intracellular Zn2+ accumulation and increased the zebrafish survival. The latter was probably achieved by reducing oxidative stress that was triggered by Zn2+. Therefore, this study supports that the increased level of intracellular Zn2+ is a critical factor in brain damage caused by hypoxic stress.
In this study, the brain damage was evaluated with TTC staining, a common method used in other mammal models. The results of TTC staining resembled those observed in other animal models. Hypoxia-induced brain injury or infarction was clearly visible in the cross section of the zebrafish brain. The zebrafish that were treated with DEDTC in hypoxic treatment yielded the improvement of TTC staining that was similar to the control. Due to the small size (about 1 mm in diameter) of the zebrafish brain, the density of the staining was quantified using spectrophotometry. Hypoxic treatment significantly reduced TTC contents. The reduction in TTC staining was reversed when DEDTC was applied (Fig. 5). The elevated intracellular Zn2+ is correlated with mitochondrial dysfunction, loss of mitochondrial defenses, and increased production of reactive oxygen species.30–32 TTC is enzymatically reduced to a red formazan product by dehydrogenases, which are most abundant in mitochondria. Zn2+ chelation, by removing excessive Zn2+ during hypoxia, may minimize mitochondrial dysfunction, and consequently preserved TTC reduction to a red formazan.
There are growing efforts to use (to develop) zebrafish as a model system in neurological studies ranging from genetic to behavioral evaluations.33–35 Using adult zebrafish as a model to study brain damage in hypoxia has not been extensively explored at the present. 8 How can the zebrafish model help the research of hypoxic-ischemic brain damages and ultimately contribute to the advancement of therapeutic drug development? First of all, the present study indicates that zebrafish are sensitive to hypoxic attack. As shown here, hypoxia-induced brain damage and death occurred in a considerably short time. Second, this is a novel animal model for global ischemia. The zebrafish was placed in the air-proof hypoxia chamber, which is similar to the conditions in a complete global ischemic stroke. In this setting, the extent of hypoxia can be regulated by the level of oxygen deprivation to achieve partial or complete hypoxia (anoxia) and by the length of time in the hypoxia chamber (transient or permanent). Therefore, this model potentially allows for the study of global brain damage caused in a variety of hypoxic conditions.
Besides its simplicity and susceptibility to ischemic brain damage, the zebrafish offers several additional advantages for use as the model system. The zebrafish can easily take dissolved chemicals with water into its body through its gills merely by breathing. Zebrafish circulation is a single circuit: heart—gills–body–heart. Chemicals are typically added to the aquatic environment in which they live, allowing for easy experimental manipulation and observation. Chemicals are absorbed directly into the circulation system without the need for nociceptive and time-consuming injection or invasive operations. There is no stress or tissue damage that is associated with rodent stroke models. It is a noninvasive animal model. Another advantage is that the zebrafish has a well-studied set of genes and fully-sequenced genome. Since there is availability of well-characterized mutants, it is practical and convenient to use gene mutations and transgenic for further study if needed. The zebrafish has increasingly been used as a whole organism model in which to perform functional high-throughput drug screens, and numerous chemical screens have been performed in zebrafish.36–40 Compared to other animal models of hypoxia/ischemia, zebrafish models are also cost effective.
The application of a Zn2+ chelator to prevent damages caused by Zn2+ overload is under active investigation as a treatment for stroke.19,41,42 In this study, the application of the Zn2+ chelator DEDTC improved the survival and tolerance under the hypoxic-ischemic treatment in a newly developed in vivo zebrafish model.8,12 As Zn2+ is required for normal biological processes, the advantage for using a moderate chelator is to maintain the level of Zn2+ in the optimal range, thereby preventing the brain damage by excessive Zn2+ while avoiding an unwanted Zn2+ deficiency. 26 At present, despite clear demonstrations of numerous agents that can prevent the cascade of events leading to ischemic neuronal death in animal models, it is not known whether any particular neuroprotectant is more effective than others to improve the outcome of a stroke in humans conclusively43–46 (also see Report of the Stroke Progress Review Group, NIH, 2002 & 2006). Therefore, the effective strategy may be a combined treatment that targets the multiple events involved in Zn2+ dyshomeostasis.
Footnotes
Disclosure Statement
No competing financial interests exist