
Abstract
Abstract
Experimental studies in the zebrafish have greatly facilitated understanding of genetic regulation of the early developmental events in the pancreas. Various approaches using forward and reverse genetics, chemical genetics, and transgenesis in zebrafish have demonstrated generally conserved regulatory roles of mammalian genes and discovered novel genetic pathways in exocrine pancreatic development. Accumulating evidence has supported the use of zebrafish as a model of human malignant diseases, including pancreatic cancer. Studies have shown that the genetic regulators of exocrine pancreatic development in zebrafish can be translated into potential clinical biomarkers and therapeutic targets in human pancreatic adenocarcinoma. Transgenic zebrafish expressing oncogenic K-ras and zebrafish tumor xenograft model have emerged as valuable tools for dissecting the pathogenetic mechanisms of pancreatic cancer and for drug discovery and toxicology. Future analysis of the pancreas in zebrafish will continue to advance understanding of the genetic regulation and biological mechanisms during organogenesis. Results of those studies are expected to provide new insights into how aberrant developmental pathways contribute to formation and growth of pancreatic neoplasia, and hopefully generate valid biomarkers and targets as well as effective and safe therapeutics in pancreatic cancer.
Introduction
Understanding the mechanisms that regulate pancreatic development has been facilitated by studies in model organisms, such as mouse, rat, frog, chick, and fish. Zebrafish (Danio rerio) is an established model of vertebrate biology as well as human diseases. 5 With its evolutionarily conserved features and its unique properties like small size, fecundity, external fertilization, and transparency, zebrafish offers advantages for morphologic, genetic, and biochemical studies. Indeed, the zebrafish system is complimentary to the rodent models for genetic analyses of the developmental processes. These include mapping of endodermal specification towards pancreatic cell fate and differentiation 6 as well as studies related to the mechanisms of human diseases.5,7 The complimentarity of the zebrafish system and mouse model is further exemplified by the recent studies that utilize forward genetic screens and genome editing using transcription activator-like effector nucleases (TALENs) in combination with live real-time imaging of development in zebrafish. Not only do the results of these studies highlight the conserved developmental processes between zebrafish and mice, but they also show that these two animal models complement each other as disease models to study organogenesis and diseases.8–10 The development of pancreas, both endocrine and exocrine, in mammals and zebrafish has previously been reviewed.1,2,11,12 A focused review of development of the endocrine pancreas in zebrafish has recently been reported. 13 The descriptive and experimental studies indicate that the developmental processes in the pancreas of the zebrafish and the associated genetic regulation are mostly conserved as those in mammals.
The unique properties of the zebrafish system have enabled the discovery of novel genetic elements and their functional roles in development of exocrine pancreas. In this article, we focus on the development of exocrine pancreas in zebrafish and update with findings from recently published literature, followed by discussion of translation of discovery in the zebrafish system into various aspects of human pancreatic cancer. First, we will critically review the developmental biology and genetics of exocrine pancreas, including formation of anlage, cell fate specification, morphogenesis, proliferation, cytodifferentiation, and growth, and compare with those in human where appropriate. Next, the potential of translating the zebrafish studies into clinical biomarkers and therapeutic targets in human pancreatic cancer will be discussed. Lastly, we will assess the zebrafish models of pancreatic cancer and how the models can be exploited for understanding pathogenesis of pancreatic neoplasia and for drug discovery.
Development of Exocrine Pancreas in Zebrafish
In adult zebrafish, the pancreas is diffusely dispersed in the mesentery and within the intestinal loops.1,14 The anatomy of adult zebrafish exocrine pancreas is similarly shown in transgenic zebrafish expressing green fluorescent protein under control of the elastase A promoter.2,15 For comparison, the adult human pancreas develops as a solitary organ within the duodenal loops. However, the microscopic structure of adult pancreas in zebrafish is very similar to that of adult human (Fig. 1). The exocrine pancreas in adult zebrafish consists of pancreatic ducts and acini with the endocrine islets dispersed within the exocrine pancreatic tissue1,16 (Fig. 1A). These morphological features are essentially the same as in adult humans (Fig. 1B). Further examination of exocrine pancreas in both larval and adult zebrafish under a transmission electron microscope shows that the pancreatic acini are composed of columnar epithelia. In the acinar cells, the nuclei are located in the basal cytoplasm and zymogen granules in the apical cytoplasm, with the centroacinar cell situated in the acinar lumen. 16 This evidence indicates that the basic structure of exocrine pancreas is mostly conserved in zebrafish and humans even at the ultrastructural level. A detailed comparison of the anatomy of pancreas in adult zebrafish and humans as well as during organogenesis is listed in Table 1. The overall structural similarities of pancreas in zebrafish and humans suggest highly conserved developmental biology and genetics as well as functions of the exocrine and endocrine pancreas in these organisms. Moreover, the morphogenetic processes during pancreatic organogenesis in zebrafish and human embryos are generally conserved at various developmental stages (Table 2). Given the anatomy and developmental processes of exocrine pancreas in zebrafish and human are highly similar, it is reasonable to speculate that the genetic regulators and biological mechanisms are shared between these organisms. These data provide support for zebrafish as a vertebrate model organism to dissect the molecular mechanisms that control pancreatic development and diseases.

Comparison of the microscopic anatomy of pancreatic tissues in adult zebrafish and human. Histological sections of
During early embryogenesis in zebrafish (2 d.p.f. to at least 7 d.p.f.), the pancreas develops as a solitary organ like the human pancreas. Unlike human, the zebrafish endocrine pancreas begins morphogenesis as a single islet that proliferates to form multiple islets as observed in human. The pancreas of adult zebrafish and human differs in the gross anatomy; while the human pancreas continues to be solitary as in the embryo; the zebrafish pancreas grows into a diffused structure and is distributed throughout the mesentery. However, microscopically and at the ultrastructural level, the pancreas of zebrafish and human remain highly similar.
h.p.f., hours-post-fertilization; d.p.f., days-post-fertilization.
References of the cited genes: available in this review and at the ZFIN database at www.zfin.org.
All emerging gene expressions are in zebrafish; where known human expressions are from ZFIN. 54
Some of the data in this table are partly adapted and modified from Tiso et al. 11
Carnegie Stage (process start stage).
not a complete list; additional and updated details available at www.zfin.org.
[w]=weeks post ovulation.
In zebrafish, an orchestrated series of signaling events coordinately lead to the formation of a pancreas with exocrine and endocrine components. These signaling events (nodal, sequential, parallel, activating, or repressing) dictate the initial endoderm induction and continue through development of a fully functional pancreas. The early steps in the development of exocrine pancreas in zebrafish include (1) induction and patterning of endoderm and formation of anlage, (2) specification of cell fate, proliferation and cytodifferentiation of progenitors, (3) morphogenesis of ducts and acini, and (4) growth of the organ. Each of these developmental steps is characterized by distinct morphological features and genetic requirements.
The endocrine and exocrine components of the zebrafish pancreas have distinct spatiotemporal origins that start off as first posterodorsal and second anteroventral buds (Fig. 2A). In zebrafish, as in other vertebrate organisms, both the endocrine and exocrine pancreas are derived from the endoderm. For induction of endoderm, cascades of signaling events are set in motion involving transforming growth factor-β, bone morphogenetic protein (Bmp), fibroblast growth factor(s) (Fgfs), and nodal-related 1 (Ndr1), and Ndr2. 6 These regulators induce a common endomesodermal territory via a concentration gradient and act through activin receptor 1b (Acvr1b). As a consequence, the transcription factors, such as Bon and Gata5 are activated and they in-turn regulate expression of Sox32, which mediates patterning of endoderm destined for pancreatic development. 11
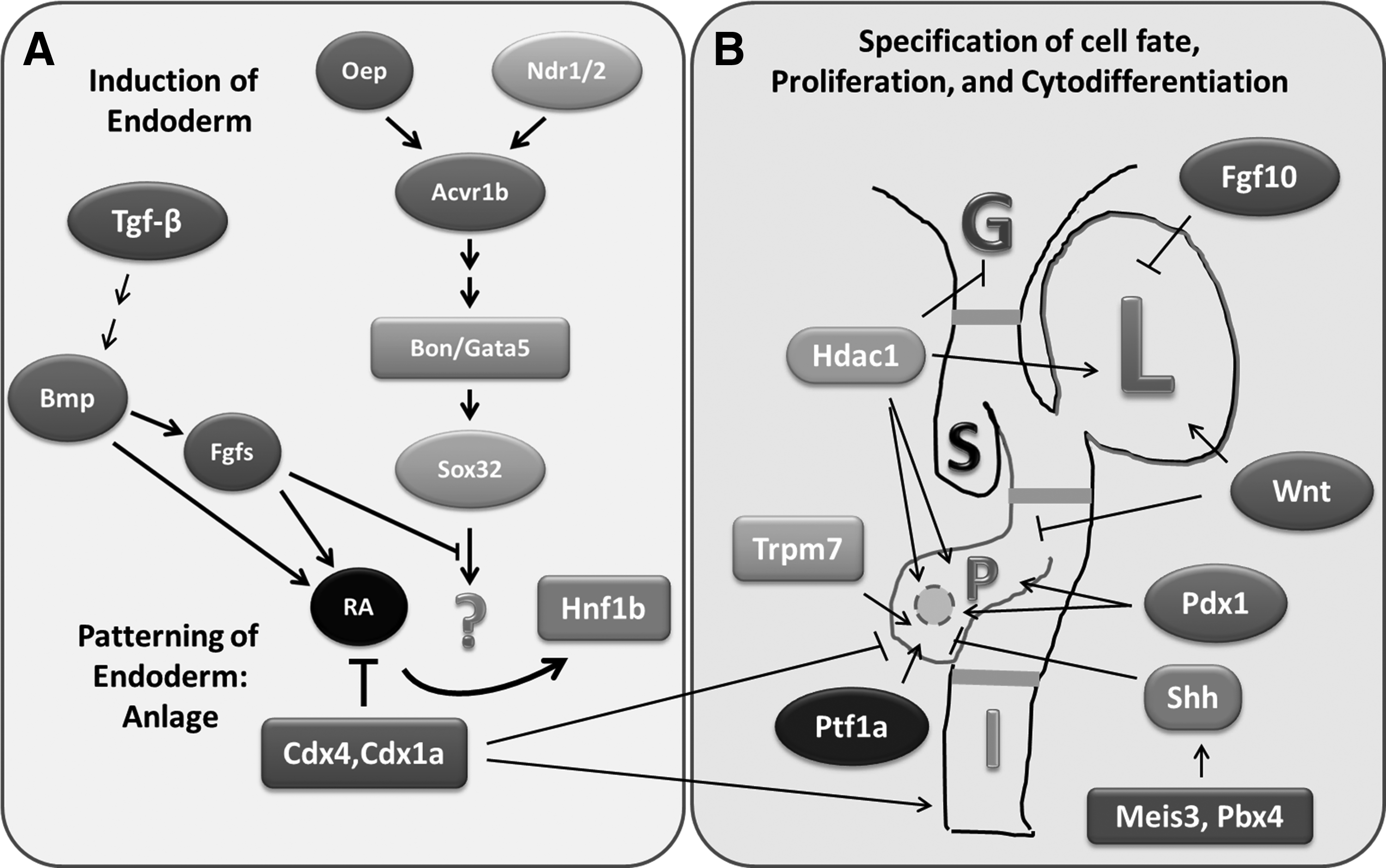
Genetic and epigenetic regulators of early pancreatic development in zebrafish. Schematic representation showing the signal transducers and transcription factors involved in regulating
After endodermal specification, the next step in pancreatic development is anteroposterior regionalization of the endoderm that will become the digestive tract (Fig. 2B). The gut patterning events are regulated by the activity of multiple signaling pathways and transcription factors. These include Fgf(s), wingless-type mouse mammary tumor virus integration site family members (Wnt), Bmp, histone deacetylase 1 (Hdac1), pancreatic and duodenal homeobox 1 (Pdx1), sonic hedgehog (Shh), and pancreas-specific transcription factor 1a (Ptf1a). Wnt signaling has been shown to play an important role in cell fate, proliferation, and differentiation; all of which influence tissue patterning. 17 Expression of Wnt inhibits pancreatic growth while promoting hepatogenesis. Similarly, retinoic acid (RA) is important for patterning of anteroposterior endoderm and formation of pancreas. Fgf10 and Fgf24 are required for specification of the ventral pancreas. The TALE homeodomain genes Meis3 and Pbx4 18 act upstream of Shh (another key patterning gene in mammals) to inhibit endocrine development (in particular insulin expression).
The exocrine component of the zebrafish pancreas arises from endodermal progenitor cells that migrate from the primitive intestine to form the pancreatic anlage.16,19 This process requires the activities of the evolutionarily conserved transcriptional factors, Pdx1 and Ptf1a.20–23 These factors are also important for pancreatic development in humans and mice, and they are expressed in pancreatic progenitors. Both Ptf1a and Pdx1 are necessary and sufficient for pancreas development. 24 Loss of Ptf1a function in zebrafish (and in humans) results in complete absence of acinar cells22,23 and significant reduction in insulin expression. 25 Similarly, lack of functional Pdx1 in humans results in pancreatic agenesis. 26 In zebrafish, the level of Ptf1a appears to be critical for determining the cell fate of pancreatic progenitors that will become exocrine and endocrine epithelia. 27 Fgf10 and Notch activate Ptf1a and Pdx1, which regulate differentiation of the exocrine pancreatic progenitors into acinar and ductal cells.16,20,22,23,27–29 Besides, Hdac1 promotes exocrine pancreas specification and cytodifferentiation as well as endocrine islet morphogenesis. 30
Proliferation of pancreatic progenitors and epithelia is controlled by various ligands and factors. Notch is required for proliferation of pancreatic progenitors, and expression of Ptf1a is dependent on the activity of Notch signaling.16,31 Optimal activity of RNA polymerase III is critical for normal epithelial proliferation that is coordinately regulated with acinar and ductal morphogenesis.2,16,32 The exocrine differentiation and proliferation factor (exdpf), a target gene of Ptf1a and a regulator downstream of RA, plays an important role in specification and proliferation of exocrine pancreatic epithelia. 33 The transient receptor potential melastatin-subfamily member 7 (Trpm7) ion channel regulates proliferation of exocrine pancreatic progenitors and epithelia through the Mg2+-sensitive pathways that involve Socs3a. 34 Several highly conserved genes involved in ribosomal biogenesis, including the causal gene for Shwachman-Diamond syndrome (sbds), ribosomal protein L3 (rpl3), and pescadillo (pes), have recently been shown to play important roles in pancreatic growth through the expansion of pancreatic progenitor cells. 7 The functional relationship among these genetic regulators of pancreatic progenitors and epithelia remains unexplored and it will be important to determine.
Besides, epigenetic mechanisms that modulate histone acetylation and DNA methylation have been shown to regulate various aspects of pancreatic development in zebrafish. Analysis of zebrafish larvae with loss-of-function in Hdac1 indicates that Hdac1 is critically required for the proliferation of exocrine pancreatic progenitors and epithelia. 35 The germline mutation hdac1hi1618 causes hyperacetylation of histones H3 and H4, and this is associated with upregulation as well as downregulation of a number of genes, including p21cdkn1a. 35 The functional roles of these targets genes of Hdac1 in development of exocrine pancreas are mostly unexplored. Moreover, the zebrafish germline mutations in DNA methyltransferase 1 (Dnmt1) exhibit degeneration of exocrine pancreas and apoptosis of acinar cells without affecting pancreatic ducts and endocrine cells. 36 These findings suggest that DNA methylation mediated by Dnmt1 is required for survival of pancreatic acinar cells during development, and the target genes remain identified. However, the complex relationship among the various ligands, molecular regulators, and the associated signaling pathways has yet to be defined. The mechanisms that control the proliferation and cytodifferentiation of pancreatic progenitors and exocrine epithelia require further investigation.
During morphogenesis of acini and ducts, the exocrine pancreatic progenitors continue to proliferate and differentiate (Fig. 3). The acinar epithelia containing zymogen granules form acini (Fig. 3A–E), and the cytokeratin-expressing ductal epithelia develop into the highly branched ductal system (Fig. 3F–J). Experimental evidence indicates that exocrine progenitors migrate and aggregate to form pancreatic ducts, rather than by budding from the exocrine anlage. 16 In agreement with this, “unpolarized” ductal morphogenesis was demonstrated by using the transgenic zebrafish expressing green fluorescent protein under control of the nkx2.2a enhancer. 37 Several mutations impair ductal branching and acinar morphogenesis, probably due to primary defects in the proliferation of pancreatic epithelia. For instance, the polr3bslj mutation, which reduces the levels of tRNAs and the proliferative capacity of pancreatic epithelia, produces hypomorphic pancreatic ducts, and acini.2,16,32,38 Analysis of the pancreatic phenotypes of Notch-deficient and Notch-activated larvae establishes a link between ductal branching and acinar morphogenesis. 16 The receptor of Shh, smoothened, is required for proper morphogenesis of exocrine pancreas, as indicated by the phenotype of the smu mutation. 15 Besides, the mutations affecting trpm7 ion channel, including the trpm7sweetbread (trpm7swd) and trpm7touchtone (trpm7tct) mutations, diminish pancreatic epithelial cell cycle progression and cell growth, impair ductal branching morphogenesis, and produce small acini. 34 Epigenetic mechanism is also involved, and this is supported by the evidence that the hdac1hi1618 mutation reduces pancreatic epithelial proliferation, and diminishes acinar and ductal morphogenesis. 35 Furthermore, the zebrafish mutations, including elysflotte lotte (elysflo), achyducttrip (achydtp), piebald (pie), mitomess (mms), and ductjam (djm) disrupt ductal branching and acinar morphogenesis to various extents; and the functional roles of the affected genes remain to be identified. 16 Continued research efforts are indicated to determine the genetic regulation of morphogenesis of exocrine pancreas. Moreover, attempt to define the relationship between acinar morphogenesis and ductal branching is expected to generate data that may shed new light into the cell of origin in human pancreatic adenocarcinoma.

Morphogenesis of pancreatic acinar glands and ducts in wild-type zebrafish.
As the pancreas continues to grow, pancreatic ducts further develop and branch, while acini enlarge and mature. The organ size is primarily determined by the amount of exocrine tissues, which is largely dependent on the number and size of acinar cells that express digestive enzymes (Fig. 4). During this stage, the acinar epithelia increase in volume, contain increased number of organelles, such as zymogen granules, rough endoplasmic reticulum, and mitochondria (Fig. 5). Mutations that impair pancreatic epithelial proliferation result in development of relatively small pancreas, as indicated by the mutations that affect polr3b, 32 exdpf, 33 trpm7, 34 and hdac1. 35 The mutations, the affected genes, and their functional roles in exocrine pancreatic development are summarized in Table 3. Further characterization of the mutations affecting exocrine pancreas, as well as the associated signaling pathways, is expected to generate new data about the mechanisms that control the growth and thus, the size of the pancreas.

Growth of exocrine pancreas during morphogenesis in zebrafish
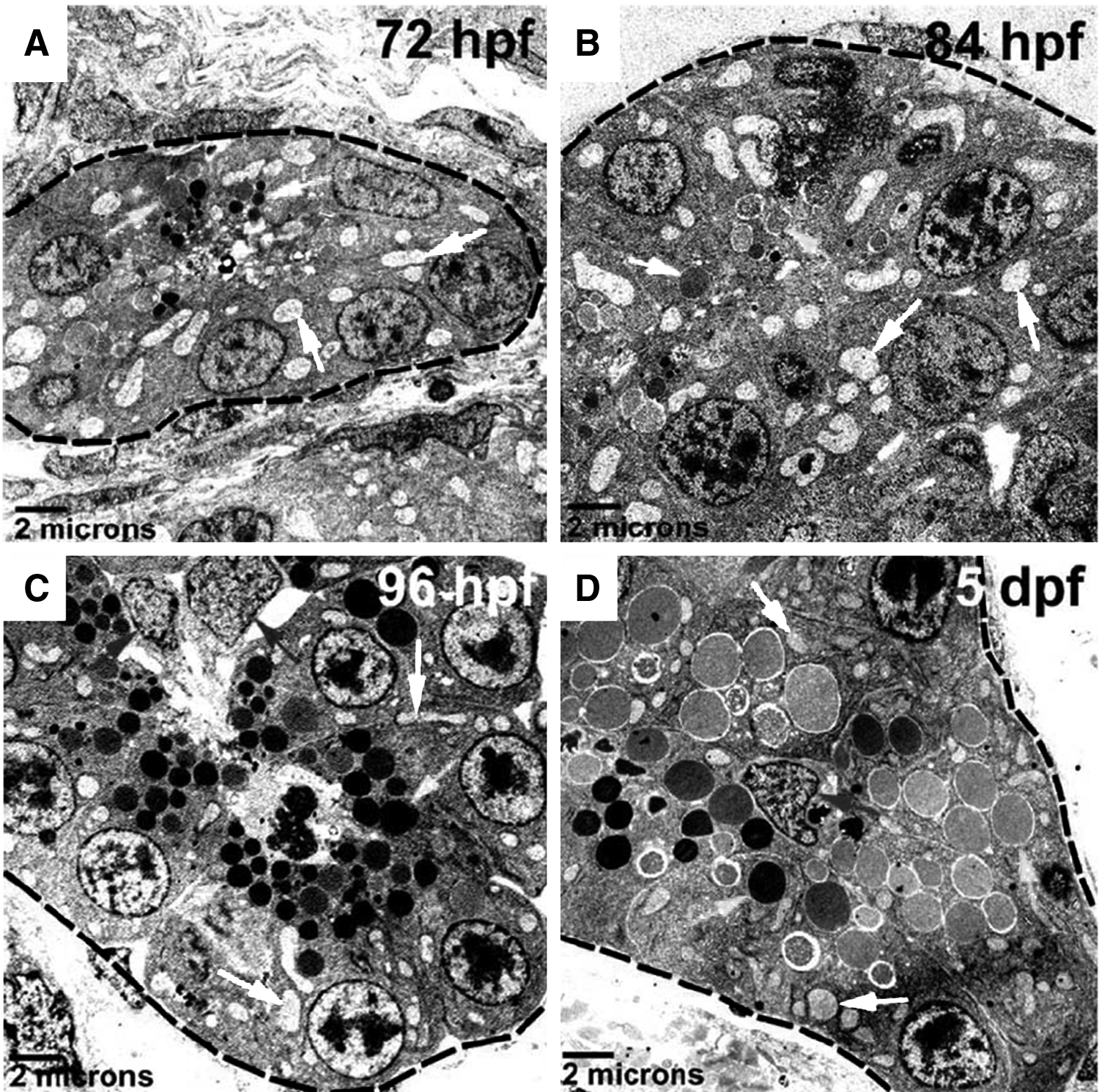
Ultrastructure of acinar epithelia in exocrine pancreas during morphogenesis
A brief homology and ontology, along with known functions of these developmental regulators in zebrafish, are presented in Table 4. These data indicate that the developmental regulators are structurally conserved between zebrafish and human on the basis of amino acid sequences and protein domains/motifs. Essentially, they suggest that the functions of these developmental regulators characterized in the zebrafish model are potentially translated to human development and diseases. We anticipate that continued research focusing on the regulatory mechanism of pancreatic development and growth in zebrafish may help understand how the aberrant control mechanism in human pancreatic epithelia and the microenvironment leads to cancerous growth and metastasis.
Identical position=primary structure of protein and identity of amino acids conserved among human, zebrafish, mouse, and rat.
Similar position=conserved amino acid property for example, may alter between L and I amino acid.
Shared synteny describes preserved co-localization of genes on chromosomes of human, zebrafish, and mouse Ortholog and paralog information obtained from Ensemble at useast.ensembl.org.
Unknown; protein IDs are listed as in zebrafish nomenclature.
Ontologies source Homologene www.ncbi.nlm.nih.gov/homologene/ and zfin.org
Developmental Regulators As Biomarkers and Targets of Pancreatic Cancer
It has been well established that the common genetic pathways are shared between embryonic development and cancer that arises in various organs—in particular, the pancreas. The developmental studies help facilitate the understanding of the pathogenetic mechanism underlying the multistep carcinogenesis. Besides, the genetic regulators of organogenesis may be exploited to develop biomarkers and targets in various human malignancies, including pancreatic cancer.
The potential of targeting the epigenetic and genetic regulators of exocrine pancreas development as a therapeutic approach in pancreatic cancer has been demonstrated in preclinical and clinical studies (Table 5). Chemical inhibitors of the epigenetic regulators of exocrine pancreas, particularly those controlling histone deacetylation and DNA methylation, have been investigated as therapeutic agents in pancreatic cancer. For instance, Hdac1 regulates gene transcription by modulating the acetylation status of nucleosomal histones and other proteins, and it is implicated in normal physiology and disease states. 39 The germ-line mutation in hdac1 or antisense oligos-mediated disruption of hdac1 expression causes reduced growth of exocrine pancreas by impairing epithelial proliferation.30,35 Treatment of zebrafish larvae with chemical inhibitors of HDACs, such as trichostatin A (TSA), impairs epithelial proliferation in the exocrine pancreas. 40 Preclinical studies indicate that the chemical inhibitor of HDACs exerts antiproliferative effects on human pancreatic adenocarcinoma, both in culture and in mouse xenograft model. 41 Clinical trials have been conducted to investigate the therapeutic efficacy of the combination of the HDAC inhibitor (suberoylanilide hydroxamic acid, SAHA) and the standard chemotherapeutic drug gemcitabine with pending result. 41
Combination of HDAC inhibitor with chemical modulators of other developmental regulators of exocrine pancreas has been shown to produce enhanced cytotoxicity in human pancreatic adenocarcinoma cells. We and others have demonstrated that HDACs and POLR3 play critical roles in the signaling mechanisms that control embryonic and neoplastic development.32,35,39,42 It was unknown if POLR3 and HDACs regulate growth of exocrine pancreas in a coordinated fashion. Whether targeting POLR3 can enhance the antitumor activities of HDAC inhibitors has not been reported. We have recently provided evidence that the combination of HDAC inhibitors (TSA or SAHA) and the POLR3 inhibitor (ML-60218) produces synergistic suppression of epithelial proliferation in the exocrine pancreas of zebrafish (Fig. 6A). This finding in zebrafish could be translated to human pancreatic adenocarcinoma, in which the combination of SAHA and ML-60218 produced enhanced antiproliferative and proapoptotic effects (Fig. 6B). Our data provide a proof of principle for enhancement of the antitumor activity of HDAC inhibitors by counteracting their “pro-oncogenic” side effects, as SAHA-induced upregulation of tRNAs was repressed by ML-60218. 40

Combined targeting Polr3 and Hdacs produces synergistic suppression of growth suppression of exocrine pancreas in zebrafish and enhanced cytotoxicity in human pancreatic adenocarcinoma.
Moreover, we have demonstrated that a combination of chemical inhibitors of HDACs and the receptor of SHH (Smoothened) produced enhanced cytotoxicity in human pancreatic adenocarcinoma. In this study, the combination of SAHA with the small molecule antagonist of Smoothened (SANT-1) produced supra-additive suppression of proliferation and induction of apoptotic death in human pancreatic cancer cells in a soft agar assay and in culture.41,43 The enhanced cell cycle arrest was associated with upregulated expression of the cyclin-dependent kinase inhibitors p21CDKN1A and p27CDKN1B, and downregulation of cyclin D1. 43 The proapoptotic effect was associated with nuclear localization of survivin, increased expression of BAX, and activation of caspases 3 and 7. 43 The potentiated cytotoxicity by the combination of SAHA and SANT-1 in pancreatic cancer cells may involve cooperative suppression of the hedgehog pathway, as shown by SAHA-induced upregulation of HHIP and repression of PTC-1 mRNAs, 43 and possibly acetylation of GLI proteins. 44
Furthermore, translation of the recent discovery of the Trpm7 ion channel in the growth of exocrine pancreas in zebrafish has led to the identification of the human ortholog TRPM7, and also its subfamily member TRPM8 as potential tissue biomarkers and molecular targets in human pancreatic adenocarcinoma (Table 5). The Trpm7 ion channel is required for normal growth of exocrine pancreas through regulation of cell cycle progression and epithelial growth 34 (Fig. 7A, B). Based on these study results, we discovered that TRPM7 and its subfamily member TRPM8 are aberrantly over-expressed in human pancreatic adenocarcinoma34,45–47 (Fig. 7C, D). Small interfering RNA-mediated silencing of TRPM7 or TRPM8 reduces cellular proliferation, impairs cell cycle progression, and induces replicative senescence in the pancreatic cancer cells.34,45–48 These data further demonstrate the potential of human orthologs to developmental regulators of exocrine pancreas in zebrafish as clinical biomarkers and therapeutic targets in human pancreatic cancer. 47
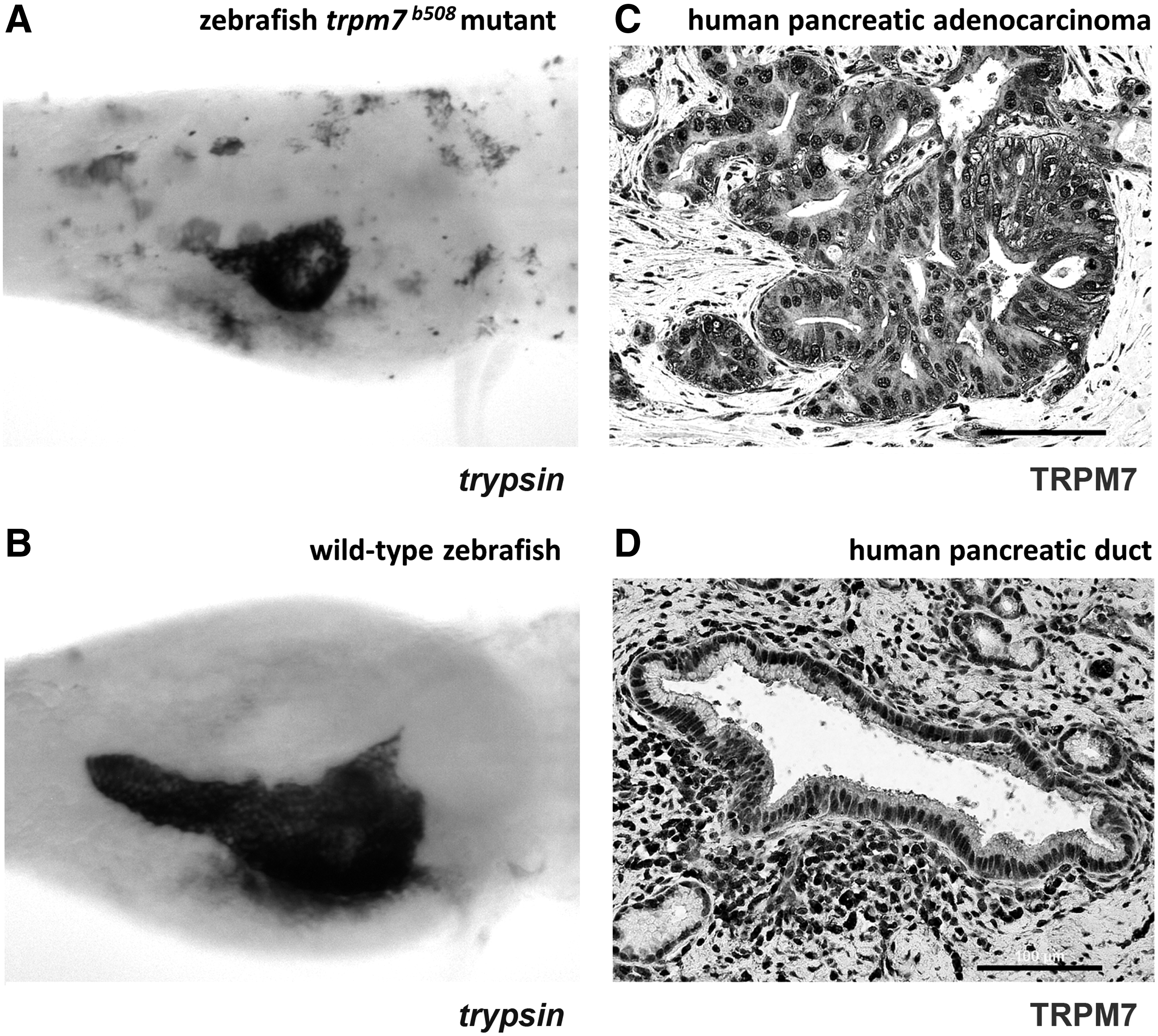
Trpm7 is critically required for growth of exocrine pancreas in zebrafish, and TRPM7 is over-expressed in human pancreatic adenocarcinoma.
Zebrafish Models for Studying Pathogenesis and Drug Discovery in Human Pancreatic Cancer
The histopathology of pancreatic adenocarcinoma in zebrafish resembles that in human (Fig. 8), suggesting similar pathogenetic mechanisms for the development of pancreatic tumor in both organisms. Zebrafish models for exocrine pancreatic tumors have been generated using germ-line mutation of oncogenic K-ras 49 and by microinjection of human pancreatic adenocarcinoma cells. 2 These zebrafish models of pancreatic cancer will be useful tools for dissecting the signaling pathways that mediate the multistep pancreatic carcinogenesis. They can be exploited for discovery and development of antitumor therapeutics in human pancreatic cancer.
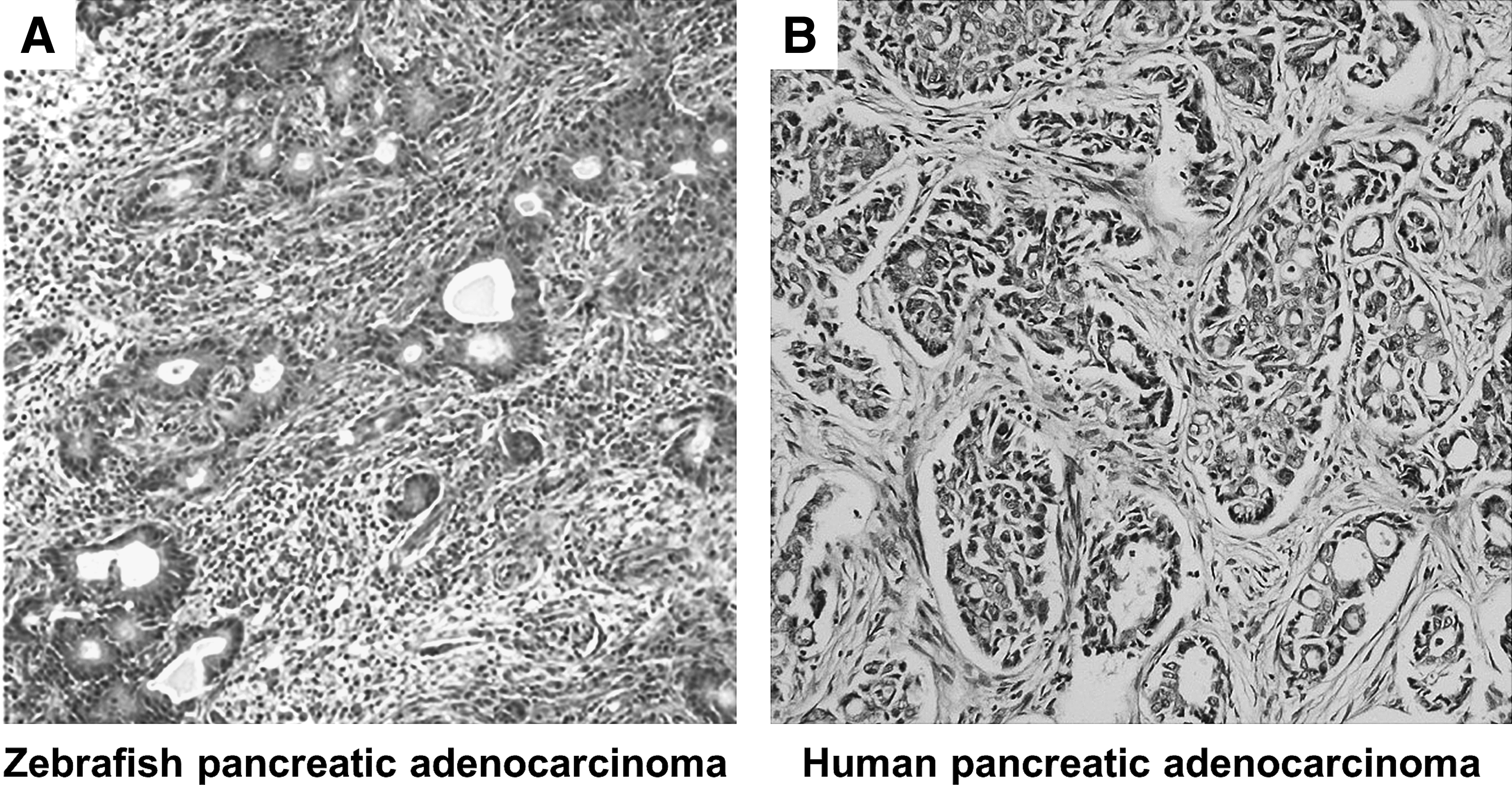
Histopathology of pancreatic adenocarcinoma in zebrafish resembles that in human.
A transgenic zebrafish model (ptf1a:eGFP-KrasG12V) was established by expressing oncogenic K-ras under control of ptf1a promoter. 49 In the oncogenic K-Ras expressing zebrafish, there is abnormal persistence of pancreatic progenitor cells which fail to differentiate in larvae, and they subsequently develop invasive carcinoma with acinar and/or ductal histological features. The pancreatic tumors in the transgenic adult zebrafish exhibit abnormally activated hedgehog signaling, which has become a therapeutic target of human pancreatic adenocarcinoma 43 (Table 5). The oncogenic K-ras expressing transgenic zebrafish model provides support for conserved histopathology and genetic pathways in pancreatic cancer between zebrafish and human. With generation of gene expression data from the human pancreatic cancer genome projects,50,51 the zebrafish model can be further utilized to advance understanding of the pathogenetic mechanisms of pancreatic adenocarcinoma. This can be accomplished by modulating the candidate genes in the transgenic zebrafish, and determining how they influence initiation, development, and progression of pancreatic tumor.
Complementary to the transgenic zebrafish model, tumor xenografts have been developed by microscopic injection of human pancreatic cancer cells into zebrafish embryos. 2 We have microinjected fluorescently labeled human pancreatic adenocarcinoma cells into the yolk sac of wild-type zebrafish larvae on 2 days-post-fertilization (d.p.f.), and tumor growth was monitored every 12 h. Between 2 and 5 d.p.f., the injected cancer cells remained as a solitary mass (Fig. 9). On 6 d.p.f., two smaller cell masses were observed; and by 7 d.p.f., at least three discrete cell masses are evident, indicating segregation of the tumor cells that resembles tumor invasion and metastasis (Fig. 9). Generation of such zebrafish xenograft model of human pancreatic adenocarcinoma should be feasible with pancreatic cancer stem cells or cell lines derived from freshly resected surgical specimens from human patients.

Zebrafish xenograft of pancreatic cancer generated with human pancreatic adenocarcinoma cells PANC-1. A suspension of 50 dil-CM labeled PANC-1 cells in 50 nL culture medium was injected under a stereo-dissecting microscope (MZ16F; Leica) into the yolk sac of zebrafish larvae on 2 d.p.f. The larvae were incubated in embryo medium at 35°C. The individual larva was kept in each well of a 24-well tissue culture plate. The tumor xenograft (arrows) in each zebrafish larvae was examined every 12 h under a stereo-dissecting microscope with fluorescent illumination at 568 nm (MZ16F; Leica). Images of the xenograft in the same larva were captured at the indicated time intervals. D, dorsal; V, ventral; A, anterior; P, posterior. Reprinted with permission from Yee 2 .
While mouse models of pancreatic cancer have accelerated our understanding of its pathogenetic mechanism and allowed preclinical testing of drug efficacy, 52 transgenesis and tumor xenografts in zebrafish offer a unique opportunity to model pancreatic cancer in a complementary fashion. In particular, the intrinsic features of zebrafish (small size, fecundity, optical transparency) enable chemical-genetic interrogation and real-time visual monitoring of pancreatic tumor growth and metastasis in vivo. 2 Moreover, these zebrafish models provide new platforms for investigation of the complex interaction between the external factors (such as environmental pollutants and chemical carcinogens) and genetic constituency in transformation of pancreatic epithelia into premalignant neoplasia and eventually invasive carcinoma. By application of chemical genetics and small molecules screening to the zebrafish models, we can improve our capability of developing effective and safe antitumor therapeutics toward the goal of prevention and treatment of human pancreatic cancer.
Conclusion and Future Perspectives
In summary, descriptive and experimental studies in zebrafish have generated new knowledge about genetic and epigenetic regulation of exocrine pancreas during development. Emerging evidence has demonstrated the potential of translating the developmental regulators of exocrine pancreas in zebrafish into clinical biomarkers and therapeutic targets in human pancreatic adenocarcinoma. Future studies using the zebrafish models of pancreatic cancer are expected to advance our understanding of the molecular mechanisms underlying tumorigenesis in a developmental context, as well as identifying and validating candidate therapeutic agents. Application of technologies, such as zinc finger nucleases, TALENs, and micro-computed tomography, in combination with inducible pancreas-specific gene expression, will further enhance the power of the zebrafish model for dissecting the mechanisms that mediate pancreatic development and cancer. These concerted efforts are expected to facilitate maximal utility of the zebrafish model, along with the mammalian models, toward the goal of personalized therapy in pancreatic cancer by targeting the molecular phenotype of tumor and its associated stroma in the individual patient. 41
Footnotes
Acknowledgments
N.S.Y. is supported by the Physician Scientist Stimulus Package from the Pennsylvania State University College of Medicine and the Penn State Hershey Cancer Institute. The research work in the author's laboratory has been supported by the Penn State Hershey Cancer Institute, the Penn State College of Medicine, the University of Iowa Carver College of Medicine, Holden Comprehensive Cancer Center at the University of Iowa, National Institutes of Health, American Cancer Society, and Fraternal Orders of Eagles. The Zebrafish International Resource Center is supported by grant P40 RR12546 from NIH-NCRR.
Disclosure Statement
No competing financial interests exist.