
Abstract
Abstract
We report a new open access database, the zebrafish CreZoo (http://crezoo.crt-dresden.de), which contains novel CreERT2-driver lines that express Cre fused to the mutated human ligand-binding domain of the estrogen receptor (CreERT2) in several tissues. Recently, the conditional Cre/loxP technology has been added to the toolbox for the precise manipulation of the zebrafish genome, but currently the number of CreERT2-driver lines is limited. To enlarge the pool of existing CreERT2-driver lines, we conducted a genome-wide screen using a gene trap cassette comprising a splice acceptor and an mCherry-tagged variant of CreERT2. All molecular and expression data obtained in this screen are summarized in the CreZoo database, which currently comprises an inventory of about 47 Cre-driver lines expressing CreERT2 in a cell- and tissue-specific manner during development and adulthood. Combined with other Cre-dependent effector lines, the CreZoo will be a great tool to manipulate the zebrafish genome.
Introduction
Recently, we and others have shown that Cre and Flp are also highly efficient in the developing and adult zebrafish, a vertebrate model system with many forward and reverse genetic advantages to study development and disease.4–8 To exploit the novel potential of site-specific recombinases in zebrafish, Clark et al. conducted a genome-wide in vivo protein trap mutagenesis screen that yielded expression-tagged, revertible mutants.9,10 To this aim, a gene-breaking transposon was used, resulting in a protein trap that efficiently disrupts gene expression and simultaneously reports protein expression of the affected locus. Subsequently, using this system, alleles were created that could restore wild-type gene function by Cre-mediated recombination. In another approach, a multifunctional gene trap vector was used to generate conditional alleles. 11 Upon integration, the vector creates a nonmutagenic full-length fluorescent fusion with the endogenous gene that can be converted into a mutant allele in a Cre-dependent manner. However, functionality of the before-mentioned approaches is dependent on Cre-mediated recombination, and currently the number of available cell- and tissue-specific Cre-driver lines is limited. To expand the existing pool of CreERT2-driver lines in zebrafish, we conducted a gene trap screen using a vector comprising of a splice acceptor (SA) and a mCherry-tagged variant of CreERT2.4,12 So far, 42 transgenic lines expressing CreERT2 in various tissues have been generated with this approach. In an attempt to categorize existing and newly generated CreERT2-driver lines from our lab, we designed an easy-to-handle database, the zebrafish CreZoo (http://crezoo.crt-dresden.de).4,12
Generation of mCherry-Tagged CreERT2-Driver Lines
To obtain a wide variety of CreERT2 insertions, a gene trap approach was performed using a vector containing an SA and an mCherry-tagged variant of CreERT2 (consisting of a single open reading frame coding for mCherry and CreERT2 separated by the viral T2A peptide sequence) followed by a polyadenylation signal 12 (Fig. 1). To avoid any SA site-specific integration bias, three different trapping vectors were generated comprising of different SA sites: SA1, rabbit β-globin 13 ; SA2, zebrafish bcl2 14 ; and SA3, zebrafish gata6 (unpublished). Using Tol2-mediated transgenesis, the various trapping vectors were injected into one-cell-stage embryos. The injected animals (F0) were raised to adulthood and crossed with wild-type fish, and the resulting embryos (F1) were screened for mCherry expression under a fluorescent microscope. In total, 1479 fish were screened so far (SA1: 1034 fish; SA2: 177 fish; SA3: 268 fish), yielding similar trapping rates (SA1: 8.7%; SA2: 15.1%; SA3: 11.6%). Embryos with detectable mCherry expression were kept and further analyzed (e.g., in situ hybridization to determine the CreERT2 expression profile, 5′RACE to identify the trapped genes). We observed a variety of mCherry expression patterns in transgenic F1 embryos, indicating that the mCherry-T2a-CreERT2 cassette was inserted in various loci in the genome and is expressed in various anatomic regions (Fig. 2).
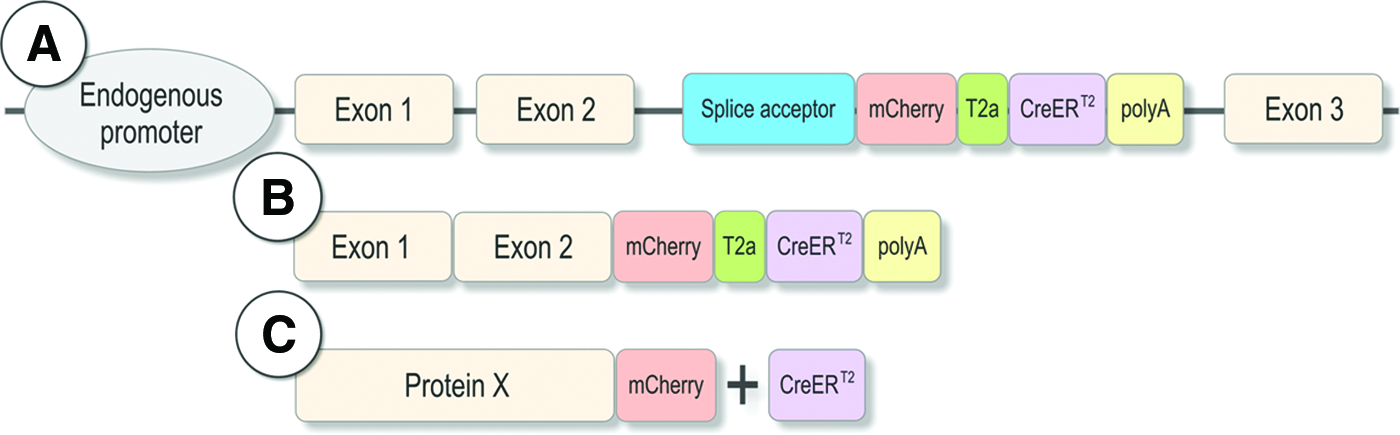
Scheme of the mCherry-T2A-CreERT2 gene trap construct, containing a splice acceptor and a mCherry-tagged variant of CreERT2 followed by a polyadenylation signal inserted into an intron. Upon transcriptional activation of the trapped gene, a fusion transcript is generated in which the exons upstream of the insertion site are spliced to the mCherry-T2a-CreERT2 cassette. DNA level

Expression patterns with respect to anatomic regions. Color images available online at www.liebertpub.com/zeb
CreZoo Database Construction
The CreZoo website has been implemented using several Open-Source technologies. The database scheme, covering the experimental data, is stored within a MySql-database server instance (www.mysql.com/). The public interface (front-end) of the database is provided by a modern web application. A separate, nonpublic web application (back-end) is used to allow involved scientists to manage the underlying data in a comfortable way. Both web applications have been developed using Grails (www.grails.org/), which itself integrates several Open-Source technologies such as Spring (www.springsource.org/) and Hibernate (www.hibernate.org/). Furthermore, Lucene (http://lucene.apache.org) has been used to provide full-text search capabilities. The user interface's client side is also powered by jQuery (http://jquery.com/). Both web applications are running on an Apache Tomcat application server (http://tomcat.apache.org/).
Structure of the CreZoo Database
The CreZoo database is designed to be a multiuser database. An introductory page provides general information about the Cre/loxP technology, the conducted gene trap screen, as well as useful links to other Cre/loxP-based approaches in zebrafish (Fig. 3). Currently, the database comprises 47 transgenic CreERT2-driver lines: 42 obtained via the described gene trap approach and additional 5 lines that have been generated using various promoter constructs.4,12 To access specific information on each transgenic line, one starts by selecting “transgenic lines” in the CreZoo database. At the moment, we have 25 of 42 transgenic insertion sites mapped (Table 1), which one can browse by selecting “genes.”

CreZoo webpage: http://crezoo.crt-dresden.de. CreZoo entries can be selected via transgenic lines
To find out the expression of a specific line, one can select “anatomy/region” for the CreERT2 expression pattern or perform a “Search” to retrieve lines by a keyword (Fig. 3A–D). Each transgenic line has its own digital notebook comprising general information, insertion site, native fluorescent mCherry and CreERT2 expression profiles, the transactivation paradigm, and miscellaneous data (Figs. 4 and 5).

Transactivation paradigm. Respective CreERT2-driver lines are combined with the Cre-dependent reporter line Tg(hsp70l:loxP-DsRed-loxP-EGFP), which expresses DsRed2 under the control of the ubiquitous, temperature-inducible hsp70l promoter, but switches permanently to EGFP after a successful recombination event. The progeny of this cross are exposed to 5 μM tamoxifen from 6 to 24 hpf to elicit recombination, heat shocked at 24 hpf for 1 h to activate reporter expression, and analyzed at 28 hpf. Color images available online at www.liebertpub.com/zeb
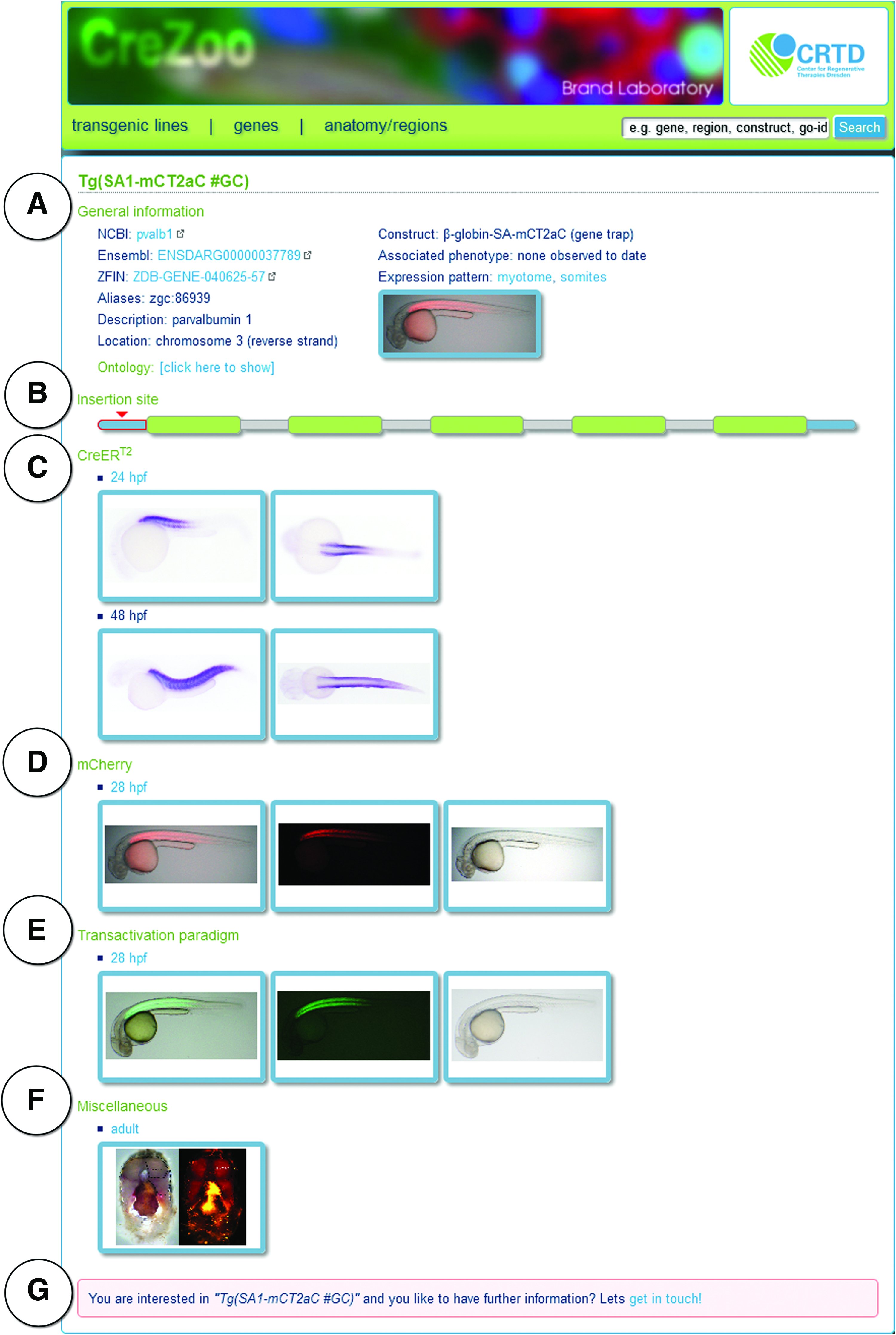
Each line has its own digital notebook comprising general information
The “General information” (Fig. 5A) summarizes common information such as gene annotation, gene ontology, and the utilized gene trap vector. Furthermore, information about “associated phenotypes” is included in this section. A schematic drawing illustrates the insertion site within the exon–intron structure of the affected gene (Fig. 5B). Embryonic CreERT2 expression analysis was done by in situ hybridization at 24 and 48 hpf (Fig. 5C). In addition, native fluorescent mCherry profiles are displayed based on developmental stages (Fig. 5D).
To demonstrate the functionality of each CreERT2-driver line, we apply a standardized transactivation paradigm (Figs. 4 and 5E). In this paradigm the respective CreERT2-driver line is combined with the Cre-dependent reporter line Tg(hsp70l:loxP-DsRed-loxP-EGFP), which expresses DsRed2 under the control of the ubiquitous, temperature-inducible hsp70l promoter, but switches permanently to enhanced green fluorescent protein (EGFP) after a successful recombination event. 6 The progeny of these crosses are exposed to 5 μM tamoxifen from 6 to 24 hpf to elicit recombination, heat shocked at 24 hpf to activate reporter expression, and analyzed for EGFP expression at 28 hpf. Driver lines with an onset of CreERT2 beyond 24 hpf are exposed to 5 μM tamoxifen from 36 to 48 hpf, heat shocked at 48 hpf, and analyzed at 52 hpf.4,12 Additional expression data can be found in miscellaneous (Fig. 5F), including in situ hybridization, immunostaining, or native fluorescent mCherry images of adult tissue.
All lines are available upon request with eventual distribution through the European Zebrafish Resource Center. For further information or to request a line, we have created a “get in touch” button, which can be found at the end of the digital notebook for each line (Fig. 5G).
Future Plans and Potential Applications
On the basis of the response that we received during the 10th International Conference on Zebrafish Development and Genetics in Madison in 2012, there is a significant need to increase the existing pool of CreERT2-driver lines. We therefore plan to expand the described gene trap approach and will integrate novel CreERT2-driver lines into the CreZoo database. We aim to identify all trapped genes using 5′RACE as well as inverse PCR and will continue to characterize the expression profile of the novel CreERT2-driver lines beyond the 24 and 48 hpf stages. The CreZoo database is designed to provide useful Cre-driver lines within the zebrafish community and thereby highly appreciates input and annotations about expression data. Combined with other Cre/loxP-based effector lines, for example, revertible mutants 9 or conditional alleles, 11 the CreZoo will be a great tool to manipulate the zebrafish genome. Moreover, in addition to these genome-wide approaches, ZCre (www.zcre.org.uk/Welcome.html) has been launched to create multilox effector lines that will further broaden the genetic tool kit using site-specific recombinases in zebrafish.
Footnotes
Acknowledgments
This project is supported by the Deutsche Forschungsgemeinschaft (BR 1746/3-1), ZF health (242048), and the SFB 655. The authors thank Sven Schreiber and the group of applied bioinformatics (Biotechnology Center, Dresden) for the implementation of the web page as well as Daniel Wehner, Oliver Bludau, Claudia Selck, and Irene Riera Tur for contributing stainings to the CreZoo database. Furthermore, we thank Dr. Anming Meng for providing the T2BGS vector.
Author Disclosure Statement
No competing financial interests exist.