
Abstract
Abstract
Background:
Although maternal smoking during pregnancy disrupts offspring development, it is not clear whether smoking before pregnancy has any effect on the next generation. Given that nicotine, the major psychoactive component in cigarettes, is toxic to many organs, we hypothesized that maternal smoking even before a pregnancy affects offspring development. Myelin is an important structure in the nervous system, and deficits in myelin are related to many psychiatric disorders and drug addiction. We therefore examined the effect of maternal exposure to nicotine on the expression of myelin genes in the offspring using zebrafish as a model.
Methods:
Female adult zebrafish were exposed to nicotine through water at a concentration of 1, 5, 10, 15, 20, 25, or 30 μM (nicotine base) for either 1 h or a continuous 24 h each day for 4 months. The nicotine-treated females were then bred with drug-naive males, and the embryos and larvae were grown in a nicotine-free environment. Maternal survival rates were calculated. Larvae of those exposed to nicotine at a dose of 1, 5, 10, 15, or 20 μM for 24 h each day were collected at 4, 7, or 14 days postfertilization (dpf). The mRNA expression of myelin-related genes was examined using quantitative RT-PCR.
Results:
The mRNA expression of most genes encoding myelin major proteins increased with age. These genes were generally downregulated by maternal nicotine exposure in 4 dpf larvae, whereas they were upregulated in 14 dpf larvae. The expression of myelin-related transcription regulators correlated well with that of myelin major proteins.
Conclusions:
Prepregnancy nicotine exposure altered myelin gene expression in the offspring, implying that maternal smoking before pregnancy affects the next generation.
Introduction
S
Regarding the effects of nicotine on the nervous system, previous studies mostly focused on drug addiction and dependence, anxiolytic effects, cognitive function, or the dopamine neurotransmitter system.4–9 Nicotine stimulates the release of the neurotransmitter dopamine in the nucleus accumbens by direct activation of nicotinic acetylcholine receptors on the neurons of the mesolimbic dopaminergic pathway. 10 Nicotine produces significant activation of cognitive functions directly or indirectly on cortical neurons. It also produces positive effects such as neuroprotection, neuroplasticity, and better performance of synaptic circuits. On the other hand, nicotine induces apoptosis and oxidative stress, and high doses of nicotine can impair memory. 11 In zebrafish, nicotine induces axon path-finding errors in embryonic development. 12 Considering that the expression of neuronal nicotinic acetylcholine receptors can be detected in oligodendrocyte progenitor cells 13 and the number of glia cells is increased in the brain of rats with prenatal nicotine exposure, 14 nicotine appears to have much more complicated functions. Very recently, it was shown that gestational nicotine exposure modifies brain myelination with sex differences in rats. 15 To expand these findings, in this study, we report the effect of nicotine on myelin genes in zebrafish.
Myelin, a multilayered membrane structure around axons, plays an important role in maintaining the functions of the nervous system.16–18 In addition to covering and insulating nerves, myelin increases the conduction of the saltatory nerve impulse, supports neuronal survival, and modulates neurotransmission.19,20 Myelin defects have been related to many psychiatric disorders, such as schizophrenia, bipolar disorder, major depression, and drug addiction.21–23 Our recent study using a rodent model suggested that maternal smoking during pregnancy alters myelin development in the brains of offspring when they reach adolescence. 15 However, it is not clear whether myelin development in offspring is vulnerable to maternal smoking before pregnancy.
The zebrafish has been used as a model to study the effects of nicotine, the major psychoactive component in tobacco.24,25 The innate characteristics of zebrafish provide many research advantages, including small size, high reproduction rates, rapid cycle time, and externally developing transparent embryos.24,25 Studies using this model have shown that direct exposure to nicotine can disrupt embryonic development,12,26–28 induce addiction behaviors, 29 reduce stress, 5 and change cognitive function. 7
Myelin in zebrafish is homologous to the protein of mammals, and thus, zebrafish have been used to study myelin development.20,30 Myelination in zebrafish correlates well with the expression of proteins, such as myelin basic protein (Mbp), myelin protein zero (Mpz), and myelin proteolipid protein (Plp1).19,31,32 These proteins are observed in zebrafish larvae as early as 2 days postfertilization (dpf). 19 Some myelin proteins, including 36k, zwilling-A (zwiA), and zwilling-B (zwiB), are specifically expressed in this teleost.33,34 The expression of myelin proteins is highly regulated at the mRNA level. Sex-determining region Y-box containing gene (Sox) 10, a transcription factor, regulates the expression of the mRNA of Mbp either by itself or by interaction with another transcription factor, Olig1.35,36 Sox10 can also stimulate the expression of Mpz in the central nervous system. 32 Kif1b, a mRNA transporter, is required autonomously for proper localization of Mbp and 36k mRNA in oligodendrocytes, the myelin-producing cells in the central nervous system.21,37
The primary objective of this study was to examine whether smoking before pregnancy affects offspring development. To reach this goal, female zebrafish were exposed to nicotine, whereas their embryos and larvae were cultured in a nicotine-free environment. Considering that myelin plays an important role in the nervous system, we assessed the expression of major myelin proteins and their regulators in the larvae.
Materials and Methods
Animal care and maintenance
Adult zebrafish were maintained in water tanks with 28°C aerated water and fed fairy shrimp. The embryos and larvae were kept in 28°C embryo medium and fed yolk-water beginning on 5 dpf and fairy shrimp larvae after 10 dpf. All animals were kept on a 14 h/10 h light/dark cycle. All experiments were performed in full accordance to the requirement by Governing Regulation for the Use of Experimental Animals in Zhejiang Province.
Nicotine administration and larvae collection
Tank water containing liquid nicotine base (Sigma, St. Louis, MO) was prepared freshly and changed every day. Adult females were exposed to nicotine at a dose of 0, 1, 5, 10, 15, 20, 25, or 30 μM for 1 h or continuous 24 h every day for 4 months except for the short time when the tank water and nicotine solution were being changed (n=7 for each group). After nicotine treatment for 1 day, 10 days, 1 month, 2 months, 3 months, or 4 months, the surviving animals were counted at each time point.
Breeding was conducted with one female and two drug-naive males in each breeding pair in a nicotine-free tank. The embryos from the breeding on the last day of the 4-month nicotine treatment at a concentration of 10 μM were continuously cultured in a nicotine-free environment. Twelve larvae from three females in each group were collected at 4, 7, or 14 dpf for RNA expression analysis.
Quantitative RT-PCR
Total RNA was extracted from larvae using the Promega SV total RNA isolation kit according to the manufacturer's instructions. Potential residual DNA in the samples was digested using Turbo DNase I at 37°C for 1 h (Ambion, Austin, TX). Total RNA concentrations were measured by a NanoDrop 1000 Spectrophotometer (Thermo Fisher Scientific, Waltham, MA). First-strand cDNAs were synthesized from 1 μg of total RNA using a Reverse Transcription System Kit (Promega, Madison, WI). Both the primers and the amplicons were tested by PAGE, and the sequences of the primers are given in Table 1. The 10 μL quantitative RT-PCR (qRT-PCR) mix consisted of 5 μL of 2×SYBR Green PCR master mix (Applied Biosystems), 0.25 μM primers, and 0.33 μL of the reverse transcription reaction mixture. The qRT-PCR was performed using the Roche Lightcycler 480 Detection System. The cycle conditions were as follows: 50°C for 1 min, 95°C for 10 min, 40 cycles of 95°C for 15 s, and 60°C for 1 min. Each reaction was conducted in triplicate in a 384-well plate with six biological replicates. Melting curve analysis was applied to characterize the specificity of the amplicons.
F, forward primer; R, reverse primer.
Statistical analysis
Expression of each gene was first normalized to the expression of β-actin and then analyzed using a comparative Ct method. 38 Fold change of each gene at one of three developmental ages (i.e., 4, 7, and 14 dpf) was shown by comparing the expression with that of the corresponding genes in the control group, and these normalized data were then analyzed by ANOVA followed by Tukey's post hoc comparison across three ages for each gene. A statistical probability of p<0.05 was considered significant.
Results
Survival rates of nicotine-exposed female zebrafish
As shown in Figure 1, in the control group, all fish survived for the 4 months examined in this study. For the 1-h daily treatment groups (Fig. 1A), the survival rate was reduced dramatically in 30 μM nicotine, and less than 30% of the animals survived after 1 day of treatment. For the 25 μM nicotine group, the survival rate declined to 42% after 10 days and 14% after 1 month, and no fish survived after treatment for 2 or more months. In the 20 μM nicotine group, the survival rate was 71% after 10 days and 57% after 2 or more months. In the 15 μM group, the survival rate was 57% after 2 and more months of nicotine exposure. In the 5 μM group, the survival rate started to decline after 2 months of treatment. On the other hand, in the continuous 24 h nicotine treatment groups (Fig. 1B), all fish survived except in the 25 and 30 μM nicotine groups, where the survival rate dropped to 57% and 14%, respectively, after 1 day of treatment. After 10 days of treatment, the survival rate had declined to 28% and 14% in the 25 and 30 μM groups, respectively, and no fish in either group survived after 2 months of treatment. For the 15 and 20 μM nicotine groups, the survival rate went down to 71% in both groups after nicotine treatment for 2 months, and no further change was observed after 3 and 4 months of treatment. As for other nicotine treatment groups (i.e., 1, 5, and 10 μM), all fish survived even after 4 months of treatment.

Survival rate (percentage surviving throughout the whole experiment compared with the initial number) of adult female zebrafish after 1 h
Maternal nicotine exposure altered myelin-related gene expression in the offspring
Dose effect
Considering that zebrafish appeared to be more vulnerable to 1 h than continuous 24 h nicotine treatment and because of our interest in the chronic effect of nicotine, we chose a treatment of continuous 24 h each day to evaluate the dose effect on myelin gene expression in the 4 dpf larvae.
Nicotine at doses of 1, 5, and 10 μM significantly decreased the mRNA expression from Mbp (p<0.05 for all doses), Mpz (p<0.05 for all doses), and Plp1b (p<0.05 for all doses) and increased that of Sox10 (p<0.05 for 1 μM nicotine and p<0.01 for 5 and 10 μM nicotine) (Table 2). None of the examined genes achieved significance in animals treated with doses of 15 or 20 μM nicotine (Table 2).
β-actin was used as an internal control, and the expression of each gene at 4 dpf in the maternal 10 μM nicotine-treated larvae was normalized to that of 4 dpf in the control larvae. The number of female zebrafish is 7, 7, 7, 5, and 5 in the 1–20 μM nicotine-treated group separately and 7 in the control group. Six biological replicates were applied in qRT-PCR experiments, and the sample of each replicate was collected from 12 larvae.
p<0.05.
p<0.01.
dpf, days postfertilization; qRT-PCR, quantitative RT-PCR; Plp1, proteolipid protein; Mbp, myelin basic protein; Mpz, myelin protein zero.
Developmental effect
On the basis of the observed survival rates of animals exposed to different nicotine doses, we chose 10 μM nicotine, the highest dose changing myelin gene expression, and the continuous 24 h treatment per day paradigm for the following developmental study. The mRNA expression of many myelin-related genes, including Mbp, Mpz, Plp1b, ZwiA, ZwiB, 36k, Olig2, Olig1, Sox10, and Kif1b, was examined in 4, 7, and 14 dpf larvae (Figs. 2 and 3).

Changes of mRNA expression of myelin genes at different developmental ages after female breeding zebrafish exposed to continuous 24 h nicotine treatment each day at a dose of 10 μM for 4 months. Changes in the expression of each gene at 4, 7, and 14 days postfertilization (dpf) are expressed as the fold change relative to expression of the corresponding gene in the control group (*p<0.05; **p<0.01). Mbp, myelin basic protein; Mpz, myelin protein zero; Plp1b, proteolipid protein 1b; ZwiA, zwilling-A; ZwiB, zwilling-B. Number of female breeding zebrafish is seven for both the nicotine-treated and control groups. Six biological replicates were applied in quantitative RT-PCR experiments, and the sample of each replicate was collected from 12 larvae.
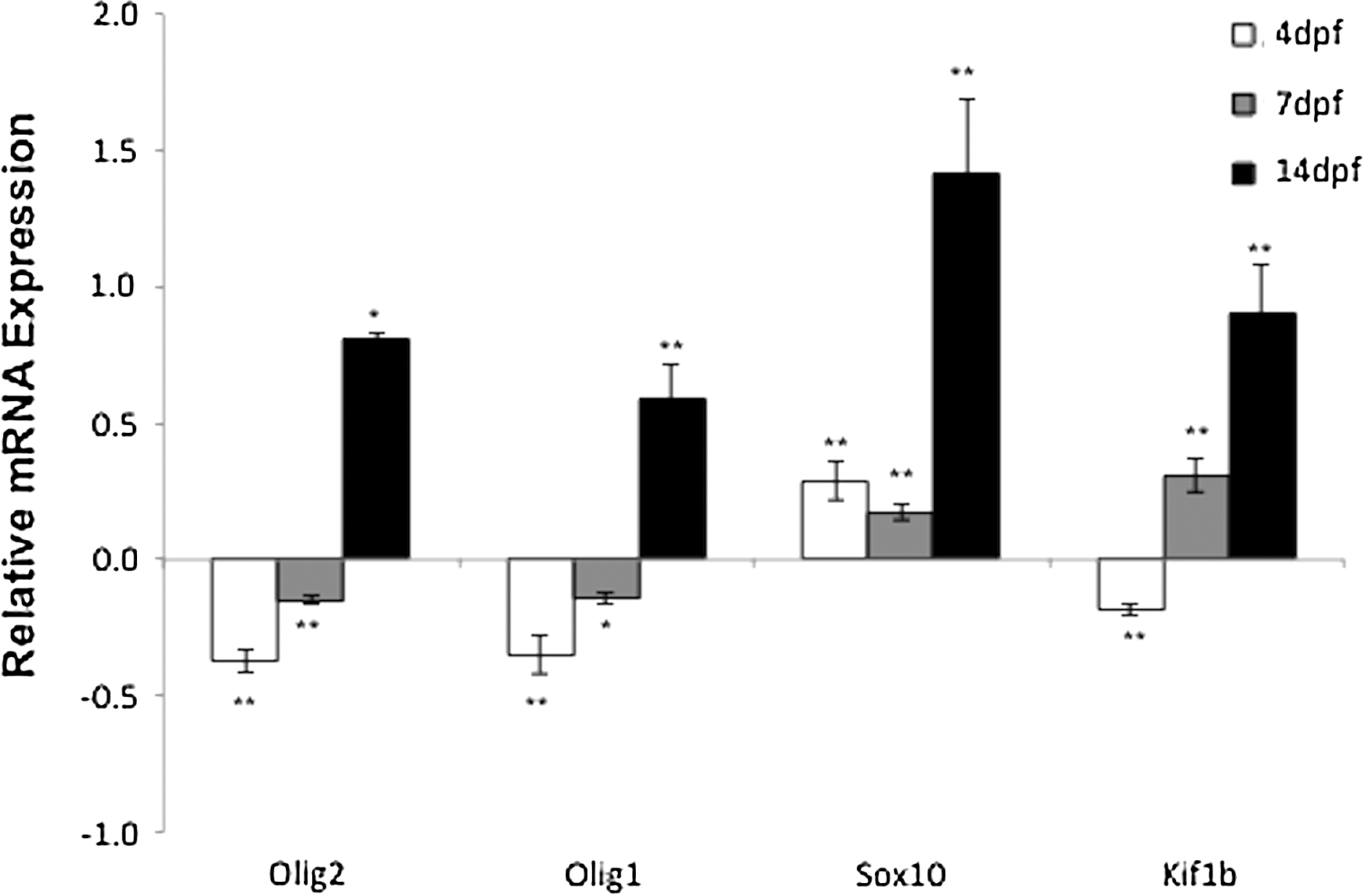
Changes of mRNA expression of transcription factors Olig2, Olig1, and Sox10 and a transporter, Kif1b, at different developmental ages after breeding female zebrafish exposed to continous 24 h nicotine treatment each day at a dose of 10 μM for 4 months. Changes in expression at 4, 7, and 14 dpf are expressed as fold change relative to the expression of the corresponding gene in the control group (*p<0.05; **p<0.01). Olig2, oligodendrocyte transcription factor 2; Olig1, oligodendrocyte transcription factor 1; Sox10, SRY (sex-determining region Y)-box containing gene; Kif1b, kinesin family member1b. Number of female breeding zebrafish is seven for both the nicotine-treated and control groups. Six biological replicates were applied in qPCR experiments, and the sample of each replicate was collected from 12 larvae.
For the major myelin protein genes, namely Mbp, Mpz, Plp1b, ZwiB, and 36k, maternal nicotine treatment significantly altered larval mRNA expression (Fig. 2). Consistent with what we found in the dose effect, 10 μM nicotine significantly decreased the mRNA expression of Mbp (p<0.01), Mpz (p<0.01), and Plp1b (p<0.01) in 4 dpf larvae. Moreover, maternal nicotine exposure significantly decreased the mRNA expression of ZwiB (p=0.03) and 36k (p=0.001), whereas the treatment increased that of ZwiA (p=0.03) in 4 dpf larvae (Fig. 2).
At 7 dpf, the expression of all genes except ZwiB and 36k had increased. At this stage, Mbp (p<0.01), Mpz (p=0.01), Plp1b (p<0.01), and ZwiA (p=0.04) were significantly upregulated by maternal nicotine exposure, whereas the expression of ZwiB (p=0.02) and 36k (p=0.01) was decreased. mRNA expression of all these genes was significantly upregulated by maternal nicotine exposure at 14 dpf (p<0.01 for all genes; Fig. 2).
We also compared the fold change of each gene (nicotine vs. control) at 4, 7, and 14 dpf. The fold change at 7 dpf was significantly higher than that at 4 dpf for Mbp (p<0.01), Mpz (p<0.01), Plp1b (p<0.01), and 36k (p<0.05). All examined genes except ZwiA showed significantly larger fold change at 14 dpf than at 7 and 4 dpf (p<0.01 for all genes except ZwiA; Fig. 2).
Next, we examined how maternal nicotine treatment affected the expression of three myelin-related transcription factors, Olig2, Olig1, and Sox10, and an mRNA transporter, Kif1b (Fig. 3). The mRNA expression of Olig2, Olig1, and Kif1b was significantly downregulated at 4 dpf by maternal nicotine treatment (p<0.01). At 7 dpf, the mRNA expression of Olig2, Olig1 was decreased, whereas that of Sox10 and Kif1b was increased by maternal nicotine treatment. At 14 dpf, mRNA expression of all four genes was significantly upregulated by maternal nicotine treatment (p<0.01 for all genes; Fig. 3).
The fold changes of these four genes in the nicotine group versus controls were also compared across three ages. The fold change in Kif1b expression at 7dpf was significantly greater than that at 4dpf (p<0.01), whereas that of Olig1, Olig2, Sox10, and Kif1b was significantly greater at 14 dpf than at 4 and 7 dpf (p<0.01 for all genes; Fig. 3).
Discussion
Although many studies have shown that maternal smoking during pregnancy disrupts offspring development, it is not clear whether smoking before pregnancy has any impact on the next generation. Using zebrafish as a model, we showed that maternal nicotine exposure changed myelin gene expression in the offspring, even though the offspring were never directly exposed to the drug. This suggests that maternal smoking history alone can affect myelin development in the offspring.
The external fertilization in zebrafish allows us to separate maternal nicotine exposure completely from fertilization and development of embryos and larvae. We found that nicotine in high doses is toxic to maternal zebrafish, as judged by the survival rate, which is consistent with a previous report. 39 For the drug effect on larvae, the expression of almost all myelin genes was significantly altered by nicotine.
The expression of all the major myelin proteins examined, namely Mbp, Mpz, Plp, ZwiA, and ZwiB, was significantly modulated in larvae by maternal exposure to nicotine. However, the effect of maternal nicotine exposure appeared to be different at different developmental ages, as most of these genes showed decreased expression at 4 dpf but increased expression at 14 dpf compared with larvae without maternal nicotine exposure. Previous studies have reported that the loosely wrapped myelin sheath is first apparent at around 4 dpf, and compact myelin starts showing up around 7dpf.19,32 Our data therefore suggest a delay in the initial formation of the loose myelin sheath around axons during early development. Alternatively, our finding may reflect a delay in axon formation. However, the effect appears to be temporary because hyperexpression of major myelin genes was observed at later developmental ages. Although the consequences of the upregulation of myelin gene expression are not clear, our previous study in a rodent model has shown similar increased myelin gene expression in the brain of male rats exposed to nicotine prenatally. 15 Together, these data suggest that smoking before pregnancy has effects on the offspring development similar to those of smoking during pregnancy.
In zebrafish, the expression of genes regulating myelin thickness and myelin compaction has not been well documented in the literature, whereas in mice, myelin thickness is determined by P0, MBP, and PMP22 in the peripheral nervous system (PNS) and by MBP in the CNS; myelin compaction is mediated by MBP and PLP in the CNS, but MBP and P0 in the PNS. 40 Loss of MBP can lead to deficiency in myelin compaction and thickness. 41 On the basis of the findings from this study, we assume that the expression change of mbp, plp, and mpz (othologue of P0) may lead to changes in myelin thickness and compaction, an idea that remains to be investigated in a future study.
Moreover, we found that the expression of a few transcription factors, including Sox10, Olig1, and Olig2, and the mRNA transporter Kif1b were significantly modulated by maternal nicotine treatment. As shown in Figure 4, these transcription factors and Kif1b are involved in regulating myelin gene expression at the mRNA level. Our data thus suggest that abnormal transcriptional regulation contributes to alterations in myelin gene expression in larvae after maternal nicotine exposure. Given the changes detected in mRNA expression of these genes, it would be interesting to know whether these changes could be detected at the protein level as well. Unfortunately, we could not perform this experiment because of the lack of appropriate antibodies for the products of these genes.
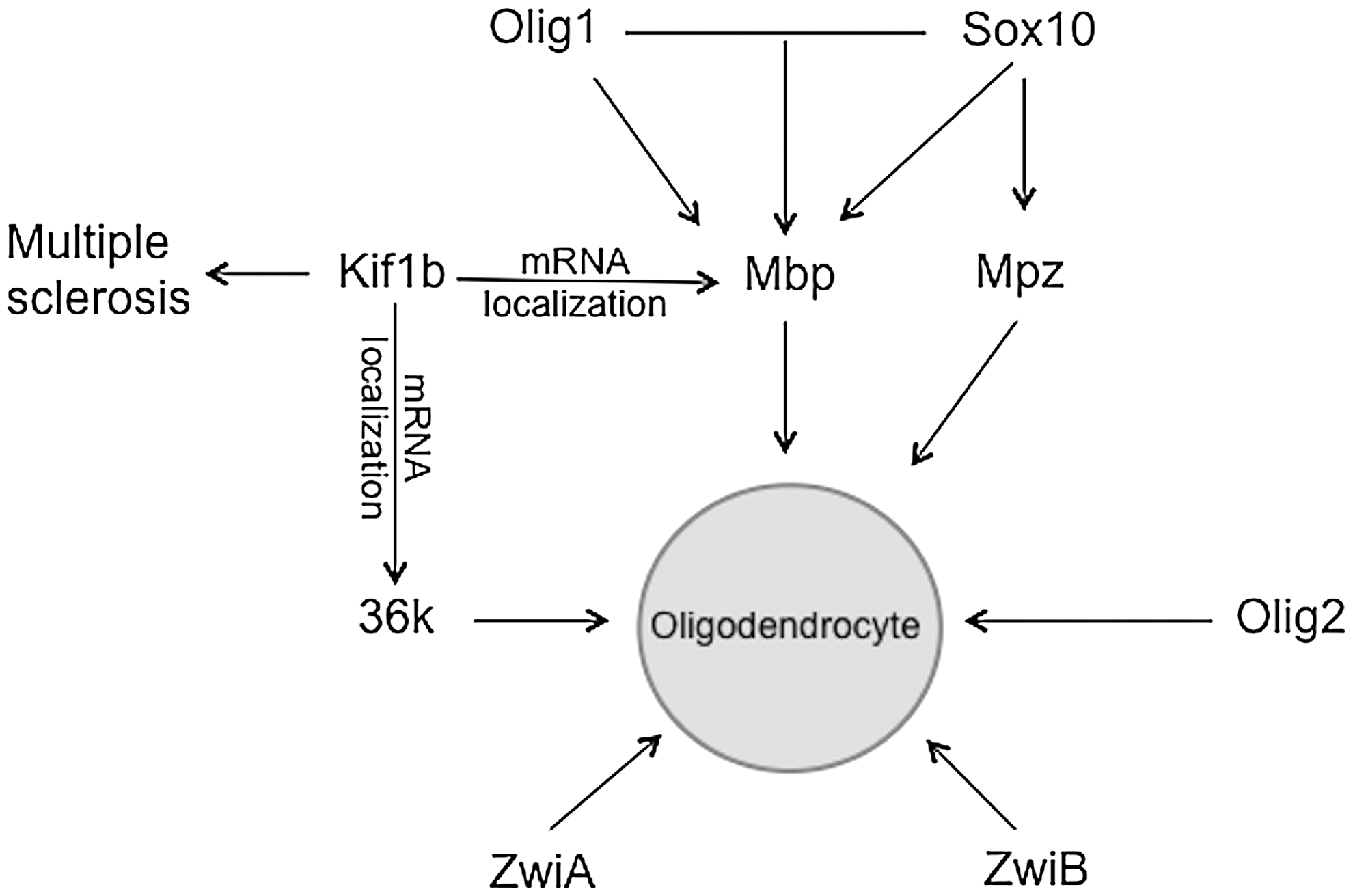
Transcriptional regulation of myelin gene expression.
Proper myelination during development is important, and the timing of axonal myelination is critical for the normal development of neurons. 42 Delayed myelination was observed in the offspring of female mice challenged with lipopolysaccharide during pregnancy; it recovered several weeks later. 43 Meanwhile, in Fmr1-knockout mice, reduction in the expression of mbp was detected at postnatal day 7, concomitant with thinner myelin than in control animals. Expression of mbp then recovered at postnatal day 14 and was upregulated at 2–4 months. 44 These findings are consistent with what we report in this study. The study on Frm1-knockout mice even implied that the reduction of myelin deposition during a critical early postnatal period may cause lasting structural and behavioral deficits even when myelin levels return to normal. 44 The findings from this and other studies suggest that maternal nicotine treatment affects early myelin deposition, which then disrupts proper myelin formation in a later stage.
Because the breeding was conducted in a nicotine-free environment and the embryos and larvae were never directly exposed to nicotine, the effect of maternal nicotine exposure appears to be passed to the next generation through gametes. Nicotine has been reported to change the morphology and maturation of oocytes and maternal gametes.45–47 Although it is not clear how the effect of nicotine on gametes could be passed to the next generation, epigenetic modification of gametes has been implicated as a major mechanism underlying transgenerational effects. 48 Epigenetic effects of nicotine, such as DNA methylation, histone modification, and chromatin restructuring, have been reported.49–53 An effect of nicotine on endocrine and metabolic systems has been observed in several generations, 54 which again suggests that nicotine has transgenerational effects through gametes.
Of note, our results indicate that zebrafish appear to be more sensitive to nicotine for 1 h of exposure per day than that for continuous 24 h of exposure every day. Although the exact reasons remain to be studied, we speculate that continuous 24 h of exposure provides sufficient time for fish to adapt to the nicotine effect, whereas 1 h of treatment per day could not, although nicotine exerts the same pharmacological effect on fish in the two treatment paradigms. We have observed this phenomenon in multiple experiments. However, this interesting observation needs to be replicated in other laboratories.
In conclusion, we observed an effect of prefertilization maternal nicotine exposure on myelin gene expression in the offspring using a zebrafish model. These findings suggest that smoking before pregnancy can alter the development of the nervous system of offspring even if nicotine expression ceases.
Footnotes
Disclosure Statement
No competing financial interests exist.