
Abstract
Abstract
The zebrafish larva is a promising whole-animal model for safety pharmacology, environmental risk assessment, and developmental toxicity. This model has been used for the high-throughput toxicity screening of various compounds. Our aim here is to identify possible phenotypic markers of teratogenicity in zebrafish embryos that could be used for the assaying compounds for reproductive toxicity. We have screened a panel of 60 water-soluble toxicants to examine their effects on zebrafish development. A total of 22,080 wild-type zebrafish larvae were raised in 250 μL defined buffer in 96-well plates at a plating density of one embryo per well. They were exposed for a 96-h period starting at 24 h post-fertilization. A logarithmic concentration series was used for range-finding, followed by a narrower geometric series for developmental toxicity assessment. A total of 9017 survivors were analyzed at 5 days post-fertilization for nine phenotypes, namely, (1) normal, (2) pericardial oedema, (3) yolk sac oedema, (4) melanophores dispersed, (5) bent tail tip, (6) bent body axis, (7) abnormal Meckel's cartilage, (8) abnormal branchial arches, and (9) uninflated swim bladder. For each toxicant, the EC50 (concentration required to produce one or more of these abnormalities in 50% of embryos) was also calculated. For the majority of toxicants (55/60) there was, at the population level, a statistically significant, concentration-dependent increase in the incidence of abnormal phenotypes among survivors. The commonest abnormalities were pericardial oedema, yolk sac oedema, dispersed melanophores, and uninflated swim bladder. It is possible therefore that these could prove to be general indicators of reproductive toxicity in the zebrafish embryo assay.
Introduction
T
The zebrafish has been extensively used in toxicological studies to screen either single compounds or small panels of compounds (reviewed by refs.18–22 ). Examples include the use of adult zebrafish for the testing of lead and uranium, 23 malathion, 24 colchicine, 25 anilines, 26 and metronidazole, 27 and the use of juveniles and embryos for testing agricultural biocides.28,29 Not only adult zebrafish, but also zebrafish embryos or larvae are used in toxicity studies (reviewed by Truong et al. 30 ). Examples of this application include the toxicity testing of nanoparticles31–34 and chemical compounds from different pharmacological classes, 11 and developmental toxicity testing of ethanol35–37 and other compounds.13,14,38–47
There are several advantages of using zebrafish embryos in biomedical research (reviewed in Ali et al. 20 ). These include the small size of the embryo, the small volume of test compound required for testing, and the relatively rapid development of the embryo. Many major organ systems are partially developed at 5 days post-fertilization (dpf).5,48 Further, cellular and molecular pathways implicated in the response to chemicals or stress, as well as many developmental pathways, show evolutionary conservation between the zebrafish and mammals.48–51 Broad homologies of zebrafish to other vertebrate species (including rodents and humans) include similarities in their genome, brain patterning, and the structure and function of several neural and physiological systems, including the stress-regulating axis.3,52–64
Although there are some essential similarities between zebrafish and mammals, there are also some important differences. The zebrafish is ectothermic, lacks cardiac septa, synovial joints, lungs, and other structures.65–67 The last common ancestor of zebrafish and mammals probably lived around 445 million years ago. 68 For these and other reasons, some phenotypic effects produced by toxicants in humans are difficult or impossible to model in the zebrafish. Another negative issue concerning the use of the early zebrafish embryo in compound screening is that it remains inside the chorion (an extraembryonic membrane) until at least 48 hours post-fertilization (hpf). 48 This is significant because the chorion may constitute a barrier to compound diffusion into the embryo.69,70
Given that the use of zebrafish embryos in biomedical research is increasing, there is a need for further research into the model. 7 Our aim here is to examine the teratogenicity to zebrafish embryos of a panel of 60 toxicants. Because we add the compounds to the water in which the embryos develop, we focus on water-soluble compounds to avoid any confusing effects of carrier solvents. The compounds were selected (in a previous study 11 ) to represent a range of chemical classes and toxicological mechanisms without a priori knowledge of their effects on the zebrafish embryo. Survivors were analyzed at 5 dpf for the presence of eight abnormal phenotypes (the remainder being classified as normal). We did not aim to make a comprehensive survey of organ morphology since this would be impracticable given the large scale of this study and the number of compounds and replicates used. Instead, we selected readouts for their ease of screening in a medium-throughput context. Note that the present study is part of a large-scale toxicity study in this lab, and the LC50 data (96-h duration of exposure) of this same panel of 60 water-soluble toxicants have been already published by us. 11 The same set of embryos used in that study are analyzed here. In addition, we have previously tested the effects of these 60 compounds on zebrafish embryo behavioral responses. 71
Materials and Methods
The embryos analyzed in this study are the same specimens for which mortality rates (and behavioral responses to toxicants) have already been reported.11,71 For the sake of completeness, we give the materials and methods used in that study 11 in the following sections, together with additional information on the EC50 calculations and malformations.
Ethics statement
All animal experimental procedures were conducted in accordance with local and international regulations. The local regulation is the Wet op de dierproeven (Article 9) of Dutch Law (National) and the same law administered by the Bureau of Animal Experiment Licensing, Leiden University (Local). This local regulation serves as the implementation of Guidelines on the protection of experimental animals by the Council of Europe, Directive 86/609/EEC, which allows zebrafish embryos to be used up to the moment of free living (∼5–7 days after fertilization). Because embryos used here were no more than 5 days old, no license is required by Council of Europe (1986), Directive 86/609/EEC, or the Leiden University ethics committee.
Animals
Male and female adult zebrafish (Danio rerio) of AB wild-type were purchased from Selecta Aquarium Speciaalzaak who obtains stock from Europet Bernina International BV. The AB strain is a wild-type strain (see www.zfin.org) and shows high genetic diversity, increasing the likelihood that we will detect idiosyncratic responses to the toxicants. Fish were kept at a maximum density of 100 individuals in glass recirculation aquaria (L=80 cm, H=50 cm, and W=46 cm) on a 14-h light:10-h dark cycle (lights on at 08.00). Water and air were temperature controlled (26±0.5°C and 23°C, respectively). The fish were fed twice daily with “Sprirulina” brand flake food (O.S.L. Marine Lab., Inc.) and twice a week with frozen Artemia (Dutch Select Food; Aquadistri BV).
Defined embryo buffer
To produce a defined and standardized vehicle (control) for these experiments, we used 10% Hanks' balanced salt solution (made from cell-culture-tested, powdered Hanks' salts, without sodium bicarbonate, Cat. No. H6136-10X1L; Sigma-Aldrich) at a concentration of 0.98 g/L in Milli-Q water (resistivity=18.2 MΩ·cm), with the addition of sodium bicarbonate at 0.035 g/L (cell culture tested; Sigma Cat. No. S5761), and adjusted to pH 7.46. A similar medium has been used previously as a zebrafish embryo buffer.11,35,72
Embryo care
Eggs were obtained by random pairwise mating of zebrafish. Three adult males and four females were placed together in small breeding tanks (Ehret GmbH) the evening before eggs were required. The breeding tanks (L=26 cm, H=12.5 cm, and W=20 cm) had mesh egg traps to prevent the eggs from being eaten. The eggs were harvested the following morning and transferred into 92-mm plastic Petri dishes (50 eggs per dish) containing 40 mL fresh embryo buffer. Eggs were washed four times to remove debris. Further, unfertilized, unhealthy, and dead embryos were identified under a dissecting microscope and removed by selective aspiration with a pipette. At 3.5 hpf, embryos were again screened and any further dead and unhealthy embryos were removed. Throughout all procedures, the embryos and the solutions were kept at 28±0.5°C, either in the incubator or a climatized room under a light cycle of 14 h light:10 h dark (lights on at 08.00). All pipetting was done manually, with an eight-channel pipetter.
Test compounds
We used water-soluble toxic compounds representing a range of different chemical classes and biochemical activities (Supplementary Table S1; Supplementary Data are available online at www.liebertpub.com/zeb). These compounds have been screened by us for embryo lethality in two previous studies.11,71 The required dilution was always freshly prepared in buffer just prior to the assay on zebrafish embryos.
Range finding
To determine a suitable range of concentrations for testing, we performed range finding using a logarithmic series (0, 1, 10, 100, and 1000 mg/L) as recommended in standard protocols. 73 Zebrafish embryos of 24 hpf were gently transferred from the Petri dish using a sterile plastic pipette into 96-well sterile microtitre plates (Costar 3599; Corning, Inc.). A single embryo was plated per well, so that dead embryos would not affect others, and also to allow individual embryos to be tracked for the whole duration of the experiment. A static nonreplacement regime was used (thus, there was no replacement or refreshment of buffer after the addition of compound). Each well contained 250 μL of either freshly prepared test compound, or vehicle only (buffer) as control. All pipetting of compounds and vehicle was done manually, with an eight-channel pipetter. We used 16 embryos for each concentration and 16 embryos as controls for each compound. The embryos for controls and treatment groups for each compound were plated in the same 96-well microtitre plates in each independent experiment.
Mortality scoring
Mortality rates at 48, 72, 96, and 120 hpf in both logarithmic series and geometric series were determined using a dissecting stereomicroscope as previously described. 11
Geometric series
After the range-finding experiments, a series of concentrations lying in the range between 0% and 100% mortality were selected. The actual concentrations used are shown in Supplementary Table S2. The concentrations were in a geometric series in which each concentration was double the next lowest value. 73 Each geometric series of concentrations of each compound was repeated three times (in total 48 embryos per concentration and 48 embryos for vehicle for each compound). The embryos for controls and treatment groups for each compound were plated in the same 96-well microtitre plates in each independent experiment.
Morphological assessment of larval phenotypes in the survivor population
Larvae at 5 days were fixed in 4% paraformaldehyde in phosphate-buffered saline at pH 7.2 at 4°C overnight. They were then rinsed five times in distilled water and dehydrated in a graded ethanol series (25%, 50%, and 70%) for 5 min each. Larvae were then rinsed in acid alcohol (1% concentrated hydrochloric acid in 70% ethanol) for 10 min. They were then placed in filtered Alcian blue solution (0.03% Alcian blue in acid alcohol) overnight. Larvae were subsequently differentiated in acid alcohol for 1 h and washed 2×30 min in distilled water. Finally, they were cleared and stored in glycerol. All larvae remained in their original multiwell plates, so that each individual could be tracked throughout the entire experimental and analysis procedure. Analysis of larval morphology was carried out using a dissecting stereomicroscope. The phenotypes were scored according to the criteria listed in Table 1.
Description of the nine categories used to score larval phenotypes at 5 dpf. See Figure 2 for selected illustrations of these phenotypes.
We take the presence of any one or more of the seven phenotypic abnormalities listed in this table to classify a compound as a teratogen for the purposes of this study.
dpf, days post-fertilization.
Statistical analysis and EC50 determination
Statistical analyses were performed using GraphPad Prism for Windows (version 5.03). To see the impact of compounds on zebrafish larvae development, we used one-way analysis of variance and Dunnett's multiple-comparison test with a probability level of 5% as the minimal criterion of significance. EC50 (expressed in mg compound/L of buffer) was determined based on morphological assessment of three independent experiments from geometric series using Regression Probit Analysis with SPSS Statistics for Windows version 17.0 (SPSS, Inc.). The EC50 in mg/L was converted into EC50 mM using the molecular weights shown in Supplementary Table S1.
Results and Discussion
We have studied the effects of differing concentrations of a panel of 60 toxicants on zebrafish development. Embryos were exposed continuously from 24 to 120 hpf. They were then assessed for malformations at 5 dpf. An overview of the data is given in Table 2 and Supplementary Table S3. Figures 1 and 2 show examples of abnormal phenotypes and the profile of teratogenic effects is summarized in Figure 3.

Concentration-dependent phenotypic abnormalities in zebrafish larvae (survivors) produced by selected toxicants [
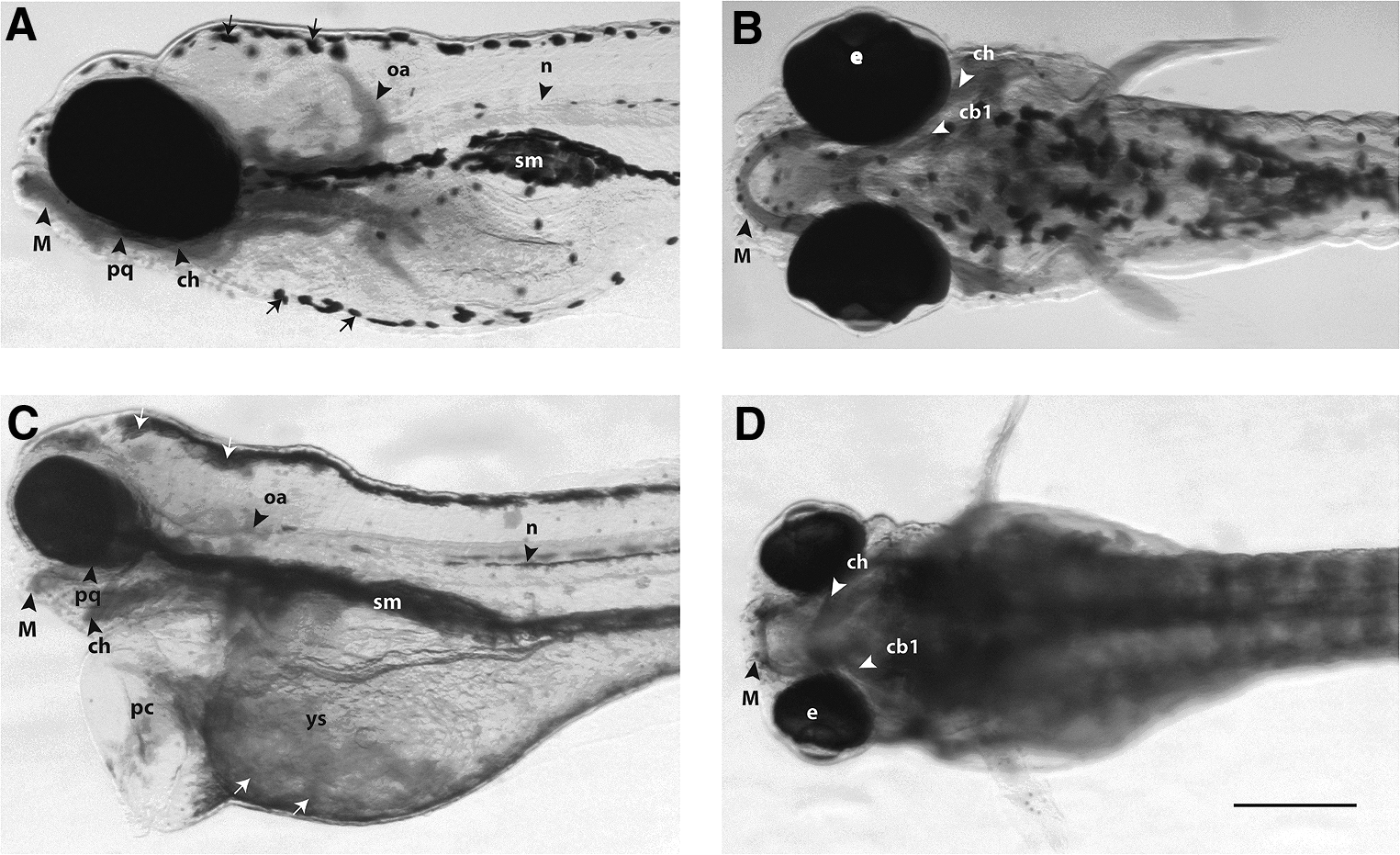
Morphological analysis of zebrafish larvae at 5 days post-fertilization. The larvae were fixed, stained with Alcian blue, and cleared in glycerol to show cartilage and other structures in the head and branchial region. The aim of this figure is to show examples of the range of malformations obtained.
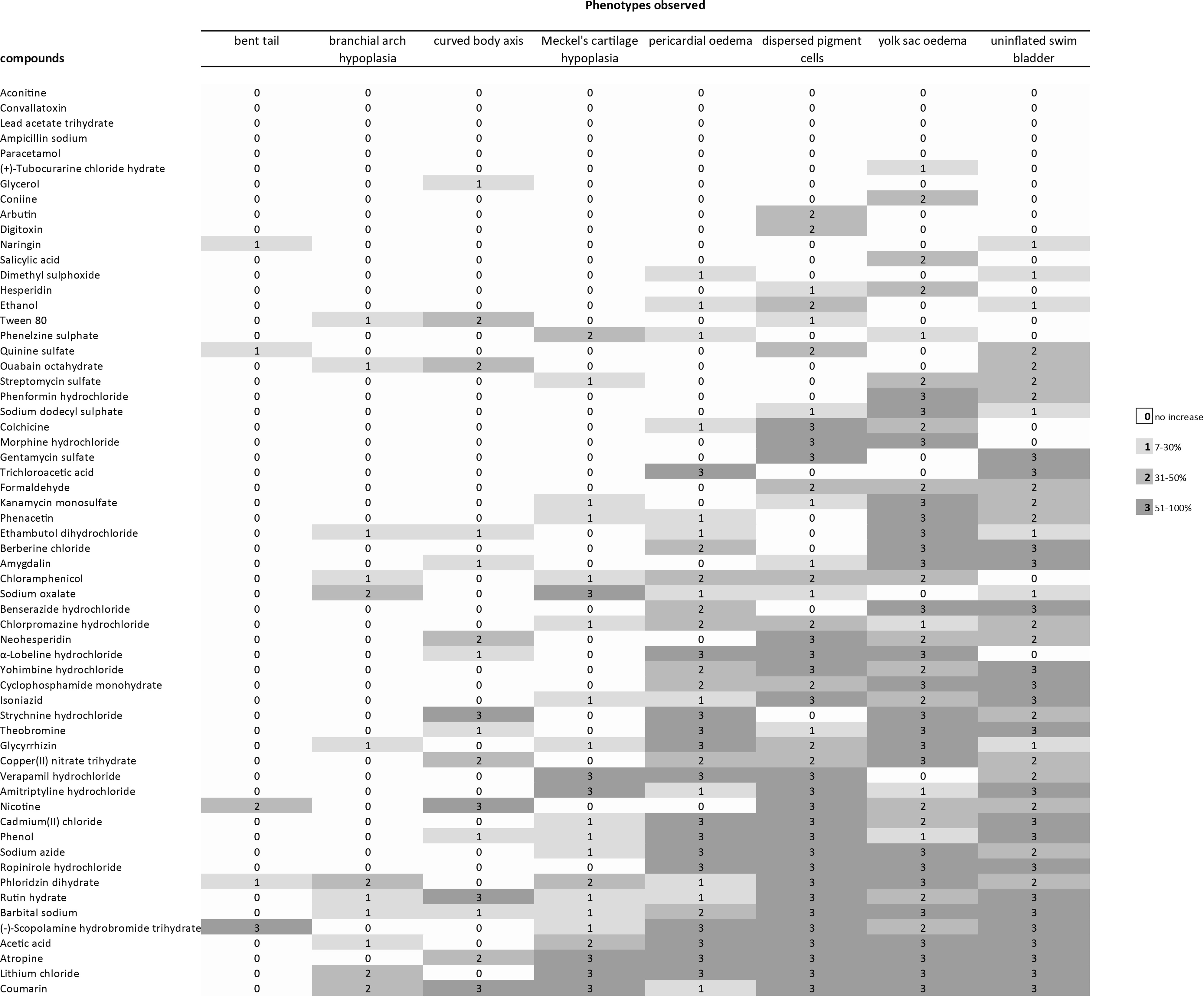
Profile of developmental effects in zebrafish larvae after exposure to different compounds. Phenotypic effects of compounds were compared to embryos exposed to vehicle only. The percentage increase of malformations at any given compound concentration is indicated by the color of the cell (see legend). The excess incidence of phenotypic defects at each concentration of each compound was calculated from three independent experiments. Only increases that were statistically significant are included; thus, in the range 7–30% increase, the values for 1–6% increase are omitted because they were not statistically significant. For full dataset, see Supplementary Table S3. As can be seen, from top to bottom, there is a general increase in the severity of effects; thus, the compounds at the top of that were not associated with a statistically significant increase in any abnormal phenotypes in survivors; by contrast, those at the bottom caused multiple abnormal phenotypes. By reading the figure form left to right, the frequency of different abnormal phenotypes is seen, with the least common on the left and the most common on the right. For the five remaining toxicants (aconitine, convallotoxin, lead acetate trihydrate, ampicillin sodium, and paracetomol) we could not find a statistically significant increase in the survivor population. This is either because the compounds are not teratogenic, or because they are so toxic that the number of survivors was too small to detect a statistically significant increase in abnormal phenotypes.
For the full range of concentrations used, see Supplementary Tables S2 and S3.
The presence of teratogenic phenotypes (i.e., any of the abnormal phenotypes 2–9 in Table 1) was scored in this column only if the incidence was significantly higher after exposure to compound as compared with vehicle.
NOAEC, no observed adverse effects (malformations) concentration.
Discussion and criticism of the methodology used
Recently, in a study using the same culture protocols (and the same embryos that we have reanalyzed here), we found that, in controls (embryo buffer only), 5% of zebrafish eggs were unfertilized, and a further 9% represented embryos that died spontaneously in the first 24 hpf. 11 Spontaneous mortality of 5–25% was also reported for zebrafish development. 74 To avoid this early mortality, we began our assays at 24 hpf. This also makes our study consistent with a previous one, in which the zebrafish was exposed to various compounds from 24 hpf to find the predictivity of zebrafish assays for the toxicity of compounds in rodents. 4 Therefore, we must assume that compounds that induce phenotype abnormalities only following early exposure will not be detected in our assays. The results are summarized in Tables 2 and 3. In previous studies, similar morphological parameters have been used to examine zebrafish embryos/larvae.13,31,75–78
Note that for aconitine, convallotoxin, lead acetate, ampicillin sodium, and paracetomol, we could not find a statistically significant increase in phenotypic abnormalities in the survivor population. This is either because the compounds are not teratogenic, or because they are so toxic that the number of survivors was too small to detect a statistically significant increase in abnormal phenotypes. Producing a statistically significant increase in any one or more of the abnormal phenotypes in Table 1.
General findings
Of the 60 toxicants tested, 55 produced one or more phenotypic effects at the range of concentrations used (Fig. 3 and Table 2; Supplementary Table S3). For the five remaining toxicants (aconitine, convallotoxin, lead acetate trihydrate, ampicillin sodium, and paracetomol) we could not find a statistically significant increase in the survivor population. This could be because these compounds are not teratogenic according to the criteria used here. An alternative explanation is that these five compounds are so toxic that there were too few survivors to yield a statistically significant change. EC50 values of compounds were estimated and shown in Table 4.
We could not estimate EC50 because only few survivors were obtained on only lowest concentration of these compounds.
n/a, not applicable; SEM, standard error of the mean.
Relationship between malformations and concentration of compound
For most of compounds, the incidence of malformations was concentration dependent. To give one example, the incidence of yolk sac oedema for coumarin was 38.9% at 70 mg/L, 47.9% (140 mg/L), and 87.5% (240 mg/L). The data in the same case for dispersed pigment phenotype were 22.2% (70 mg/L), 29.2% (140 mg/L), and 100% (240 mg/L) (see Fig. 1A). Further, selected examples are shown in Figure 1B–D and the full dataset in Supplementary Table S3. These data are consistent with previous studies14,38,76,77 in which incidences of malformations were concentration dependent.
The results of morphological analyses of larvae are summarized in Table 2. The wide range of phenotypic effects that can be seen in one treatment group are illustrated in Figure 2, which compares an untreated larva (Fig. 2A, B) with larva exposed to scopolamine hydrobromide trihydrate with 4 g/L (Fig. 2C, D). The larva in Figure 2C and D shows a range of phenotypic abnormalities, including Meckel's cartilage hypoplasia, branchial arch hypoplasia, pericardial oedema, yolk sac oedema, “dispersed” morphology of melanocytes, and uninflated swim bladder.
Compound specificity of malformations
A range of phenotypic effects were recorded in zebrafish larvae after exposure (Fig. 3 and Table 2; Supplementary Table S3). Uninflated swim bladder, yolk sac oedema, and dispersed pigment cells were the phenotypes most frequently observed. Bent tail, brachial arch hypoplasia, and bent body axis were the least recorded phenotypes (Fig. 3). Five compounds (Fig. 3) did not produce any significant effects. Five produced only one effect (Fig. 3) while the majority of compounds (50) produced multiple phenotypic effects (Fig. 3).
That types of malformation that we report here after exposure to cadmium(II) chloride, chloramphenicol, lithium chloride, chlorpromazine hydrochloride, kanamycin, and isoniazid (Fig. 3) are consistent with previous studies using zebrafish embryos.14,38,77,79 Pericardial oedema induced by glycyrrhizin has also been reported in a human adult. 80 Other literatures are consistent with the compound-specific malformation reported here (Fig. 3): the report of craniofacial malformations in mouse embryos exposure to lithium, 81 craniofacial defects in monkeys exposed to Isoniazid, and craniofacial defects in rats exposed to ethambutol (reviewed in Holdiness 82 ). The teratogenicity of colchicine is known from one case study of a human fetus that had heart malformations 83 ; however, another larger scale study found no evidence of teratogenicity of colchicine in humans. 84
It could be argued that, by beginning exposure at 24 h, we are missing out on early developmental toxicity effects, such as the action of compounds on gastrula stages. However, this is likely to be a general phenomenon because other compounds mainly cause embryo death at these early stages. For example, we showed 35 that exposure of zebrafish embryos at early stages (dome to 26-somite) to ethanol resulted in high mortality, while exposure at later stages (prim-6 and prim-16) led to a high incidence of malformations. Other compounds, such as copper and cadmium, also show a development window of sensitivity and are more toxic to larval stages than to embryonic and adult stages of freshwater fish species.85,86
These previous results are consistent with a time window of sensitivity within the range of stages exposed here. It should also be remembered that early embryos may be shielded from teratogen actions because of the presence of the chorion at early stages. This membrane acts as a possible barrier to diffusion of some compounds.35,69,70
Teratogenicity per compound class in zebrafish larvae
To see whether the variation in developmental toxicity of compounds screened in the zebrafish assay was due to compound class, we sorted the compounds by chemical class according to Ali et al. 11 The classes were alcohols, alkaloids, amides, carboxylic acids, glycosides, and the remaining compounds (others). The break-down by compound class shows that teratogens were detected in all compound classes (Table 3).
Conclusions
Our findings show that teratogenicity assessment on zebrafish larvae can provide a sensitive evaluation of the teratogenicity of a wide range of toxicants. Thus, it could, in principle, also provide a useful tool in the screening of new drugs for treating human diseases. The zebrafish larval assay is compatible with high-throughput screening and can be implemented early in the drug-discovery pipeline for early assessment of drug safety. However, future work in the validation of the zebrafish larval assay must include a wider range of compounds, including those that are known teratogens in humans.
Footnotes
Acknowledgments
The authors thank Merijn A.G. de Bakker and Peter J. Steenbergen for expert technical assistance and zebrafish breeding.
This work was supported by the Smart Mix Programme of the Netherlands Ministry of Economic Affairs and the Netherlands Ministry of Education, Culture and Science (M.K.R.) under grant number SSM06010 and University of Azad Jammu and Kashmir, Pakistan (S.A.) under project (no. F-3/PD/Main and Mirpur/369/2007). Note that the funding bodies had no role in designing, conducting the study, analyzing data, and writing article.
Authors' Contributions
S.A. and M.K.R. designed the experiments; S.A. conducted the experiments; M.K.R. provided the materials and facilities; J.A. scored the zebrafish larvae for morphological malformations (and S.A. rechecked a sample); S.A., J.A., and M.K.R. analyzed the data; and S.A. and M.K.R. wrote the article.
Disclosure Statement
The authors have no competing interests to declare.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.