
Abstract
Abstract
Paclobutrazol (PBZ), a trazole-containing fungicide, is widely used on food crops. Frequent usage of PBZ may contaminate water, but its toxicity to aquatic organisms is understudied. Although the chronic effects of PBZ exposure on reproductive, antioxidant defense, and liver metabolism systems in rockfish have been reported, the toxic effects of PBZ on aquatic embryos are unknown. Here, we report that PBZ disrupts the development of heart and craniofacial cartilage in zebrafish embryos, and decreases their survival and hatching rates. PBZ affects the normal process of cardiac looping, which may lead to a slower heart beat accompanied by pericardia edema and apoptotic myocytes. PBZ also decreases the population of migratory neural crest cells, which give rise to craniofacial cartilage. Our results reveal high embryonic toxicity of PBZ on aquatic organisms, and thus hold significance for the impact of fungicides on public health and ecology.
Introduction
P
Zebrafish is a small freshwater fish, the embryos of which have been used for developmental toxicity testing of environmental chemicals. Zebrafish is suitable for such studies, as its housing and husbandry require little space and low cost. In addition, zebrafish have high fecundity—a single breeding pair can produce 300 embryos per week. Further, zebrafish embryos are transplants, and grow rapidly (most organs form by 3 days postfertilization) and independently from the maternal fish, making for ease of observation of developmental events. Most importantly, embryonic stages, organogenesis, and development processes in zebrafish are well characterized.
The heart is the first and most important organ to form during vertebrate embryonic development. In zebrafish, heart progenitor cells, including endocardial and myocardial precursors, bilaterally align along the embryonic axis by 12 hours postfertilization (hpf). Endocardial precursors are located at the most medial position, and myocardial precursors at the most lateral. Subsequent steps involve complex morphogenetic movements, which ultimately form a functional heart. First, myocardial precursors migrate medially to enclose endocardial cells, thereby forming a heart tube at around 19 hpf. The heart tube starts beating to enable circulation by 24 hpf, and distinct ventricular and atrial chambers form by 30 hpf. By 36 hpf, the venous end of the heart orients to the left of the midline, and the heart undergoes looping morphogenesis. Finally, by 48 hpf, specialized endocardial cells form cushions at the arterioventricular boundary, which give rise to functional valves.
The vertebrate head skeleton is subdivided along the anterior–posterior axis into reiterated segments, the pharyngeal arches, and a dorsal neurocranium. It has been determined that in all vertebrates, the pharyngeal skeleton derives from the neural crest, which in turn originates in the dorsal neural tube during embryogenesis, and migrates ventrally to populate the pharyngeal arches. In zebrafish, a functional larval skeleton develops for only a few days. The head skeleton of the zebrafish contains seven pharyngeal arches, which support the feeding and gill-breathing structure, and a neurocranium, which protects the brain and sensory organs.
In this study, we demonstrate that 20 ppm of PBZ, a dose far lower than that applied to crops, caused a decreased hatching rate and increased lethality in zebrafish embryos. Decreasing the PBZ concentration tenfold still disrupted the heart and head development. These results emphasize the potential of PBZ to induce acute toxic effects on aquatic organisms when used in agriculture.
Materials and Methods
Embryo collection
Zebrafish (AB strain, wild type) were maintained under standard laboratory conditions at 28.5°C. Embryos were collected by natural spawning and staged as described previously. 10
PBZ exposure
PBZ (Sigma Cat No. 46046) was dissolved in dimethyl sulfoxide (DMSO) to a stock concentration of 300,000 ppm and stored at −20°C. Twenty newly fertilized embryos in 2 mL of sea salt egg water (0.0375% sea salt in deionized distilled water) were added to each well of a six-well plate, and the PBZ stock solution was added to each well to a final concentration of 5, 10, 20, 50, 100, or 150 ppm (equivalent to concentrations of 17, 34, 68, 170, and 340 μM, respectively). Control embryos were treated with 0.01% DMSO; each concentration was tested in triplicate for each experiment. Embryos were cultured at 28.5°C until analysis. For certain experiments, 0.2 mM 1-phenyl-2- thiourea (Sigma-Aldrich) was added to inhibit pigmentation, thereby improving visualization of internal structures.
Survival rate and hatching rate analyses
One hundred embryos treated with different concentrations of PBZ were used for analyzing the survival rate and hatching rate, which were calculated and averaged from triplicate samples. Unfertilized embryos were removed 4 h after fertilization (the remaining embryos were considered as the total embryo number) and dead embryos were removed every day. The survival rate was calculated by dividing the surviving embryos by total embryos for each concentration every day until 5 days postfertilization (dpf). The hatching rate of embryos for each concentration was obtained by dividing the surviving embryos by total embryos at 5 dpf.
Heart beat measurement
Ten embryos in each treatment group were randomly selected at 3 dpf. Embryos were embedded in 2.6% methylcellulose on a slide in the correct orientation. The number of heart beats in a 1-min interval was counted using an Olympus microscope.
Head size and eye area measurement
To determine the size of the head and eyes, 10 embryos in each treatment group were randomly selected at 5 dpf, and imaged from the lateral view. The head size and eye area in the images were measured using ImageJ.
Cartilage staining
Alcian Blue staining was performed as previously described. 11 Embryos at 5 dpf were anesthetized and fixed overnight in 4% paraformaldehyde at 4°C, and were bleached in 1 mL of 10% H2O2 supplemented with 50 μL of 2 M KOH for 1 h. They were then stained overnight in 0.1% Alcian Blue dissolved in acidic ethanol, washed extensively in acidic ethanol, dehydrated, and stored in 80% glycerol. For improved exposure of cartilage elements, embryos were digested with proteinase K.
Whole-mount in situ hybridization and cell death assay
Whole-mount in situ hybridization was performed as described previously. 12 The cmlc2 and amhc probes were used to mark cardiac cells,12,13 and the col2 a1, sox9a, and dlx2 probes were used to mark the chondrocytes and migratory neural crest cells.14–17 Stained embryos were placed in 80% glycerol and photographed with the Olympus microscope imaging system.
TUNEL labeling was performed as described previously, with modifications. 18 In brief, embryos were staged, fixed, and stored in methanol at −20°C. After rehydration, embryos were incubated for 1 h at 37°C in terminal transferase and biotin-labeled ddUTP (Roche), and then extensively washed in phosphate-buffered saline with 0.1% Tween 20. Negative controls were treated with fluorescein-labeled dUTP polymers only. To detect incorporated biotin-labeled ddUTP, embryos were incubated with a phosphatase-conjugated anti-fluorescein antibody, and colorized using the NBT/BCIP staining buffer at room temperature until staining was visible.
Statistical analysis
The toxicity of PBZ on zebrafish embryos was determined based on the frequency of the observed abnormalities. Data were subjected to the analysis of variance (ANOVA) or chi-squared test, and are presented as the mean±standard deviation. For the hatching rate, defective heart looping, heart rate, head width and length, eye area, and head cartilage length, significance differences between the treatment groups were evaluated using the one-way ANOVA followed by the Fisher's least significance difference test (p<0.05). All statistical analyses were performed using SSPS 21.0 (IBM).
Results
Survival and hatching rates of zebrafish embryos are decreased after PBZ exposure
To determine if PBZ causes embryonic toxicity in aquatic organisms, we first examined the survival rates of zebrafish embryos after PBZ (5, 10, 20, 50, 100, or 150 ppm) exposure for 5 days (Fig. 1A). One day after PBZ exposure, only embryos exposed to 100 or 150 ppm of PBZ exhibited significantly decreased survival rates as compared to embryos treated with DMSO (control) or low dosages of PBZ. Two days after PBZ exposure, all embryos treated with 150 ppm of PBZ were dead, and only 8.3%±1.2% of embryos (n=406) treated with 100 ppm of PBZ remained. Embryos treated with 50 ppm of PBZ exhibited a high death rate, but a significant difference with the control was only observed at 5 dpf. At 5 dpf, the survival rate of embryos exposed to 20 ppm of PBZ was also significantly decreased. These results indicate that 5 days of exposure to 20 ppm of PBZ greatly reduced embryo viability. We also examined the hatching rate of zebrafish embryos after PBZ treatment at 5 dpf; dead embryos were considered as failures to hatch (Fig. 1B). We found that 20 ppm of PBZ or higher had a significant inhibitory effect on embryo hatching as compared to controls. Of note, all embryos exposed to 100 ppm of PBZ failed to hatch at 5 dpf. This suggests that exposure to high doses of PBZ causes embryonic mortality and hatching retardation, indicating that PBZ is potentially developmentally toxic to aquatic organisms.

Effect of PBZ treatment on survival and hatching rate of zebrafish embryos. Embryos were treated with dimethyl sulfoxide (DMSO) (control) or the indicated concentrations of PBZ, and then
Pericardial edema in zebrafish embryos after PBZ exposure
We also observed pericardial edema in PBZ-treated embryos, even at low doses (5, 10, and 20 ppm) and at 2 dpf (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/zeb); this phenomenon was more pronounced at later stages (Fig. 2 and Supplementary Fig. S1). Magnified views of the heart chambers revealed that 3 days of PBZ exposure also caused failure of heart development (Fig. 2A). More than 99% of control embryos (n=395) (treated with DMSO) presented with a normal pericardium, while all of 10 (n=385) or 20 ppm PBZ-treated embryos (n=385) presented with pericardial edema, including mild edema with enlarged ventricles and atria, and severe edema with elongated ventricles and atria, forming a tubular-like heart. Of the embryos (n=390) exposed to 5 ppm PBZ, 6.5% exhibited a normal pericardium, and 93.5% exhibited pericardial edema (Fig. 2B).
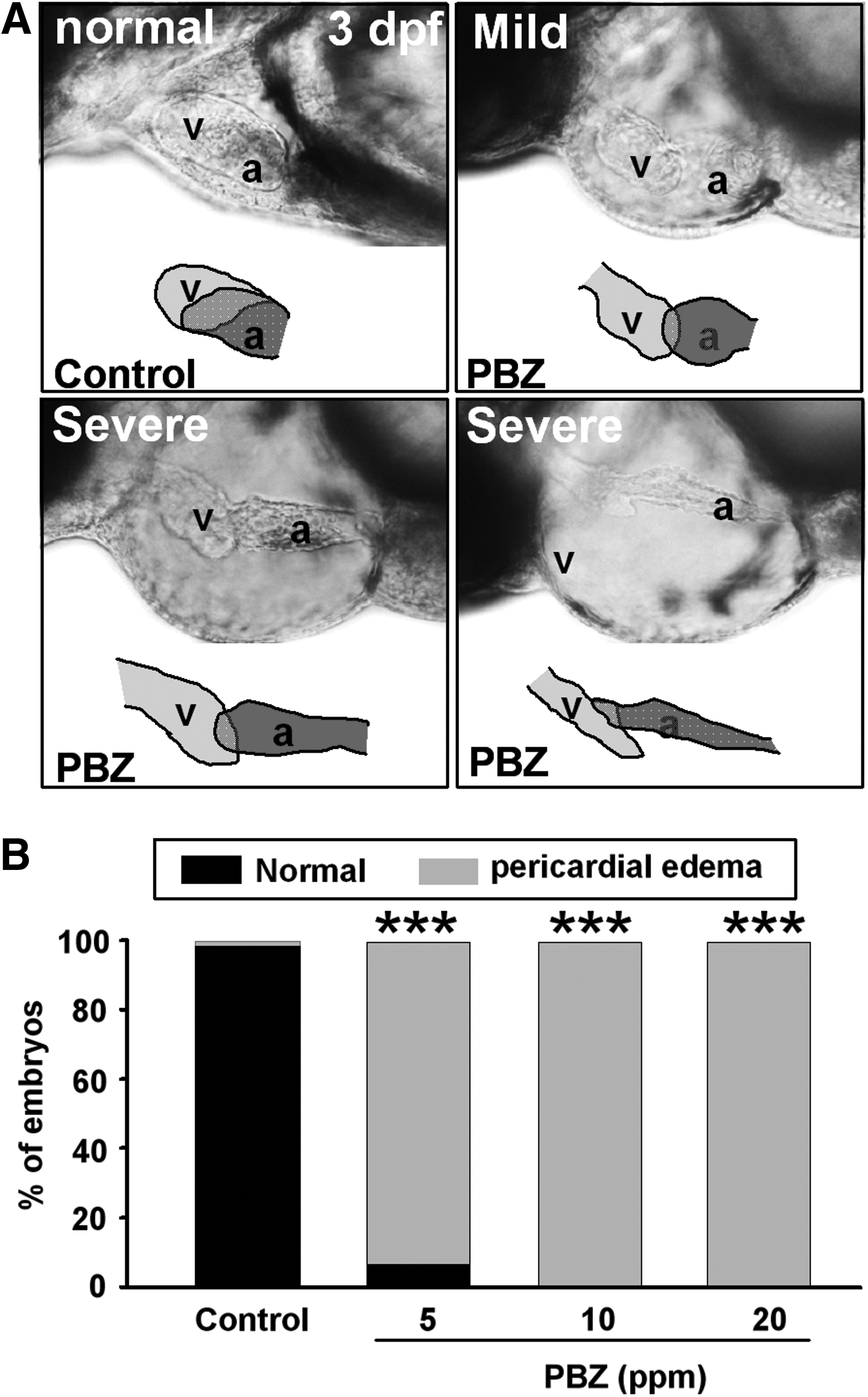
Pericardial edema and abnormal heart morphology in zebrafish embryos after PBZ treatment.
Heart disorientation and looping defects in zebrafish embryos exposed to PBZ
Substantial evidence indicates that pericardial edema is highly associated with heart looping defects in embryos.19–22 Indeed, we observed this defect in embryos treated with 5, 10, or 20 ppm PBZ by 3 dpf (Figs. 2A and 3). During normal zebrafish embryogenesis, the looping process results in the ventricle and atrium lying side by side, so that the two chambers largely overlap with each other from the lateral view at 3 dpf. In contrast, PBZ-treated embryos exhibited stretched-out hearts, such that the ventricle was positioned anterior to the atrium. Thus, little, if any, overlap was observed between the two heart chambers in PBZ-treated embryos. This result was confirmed by performing in situ hybridization to visualize the expression of cardiac myosin light chain-2 (Fig. 3A), which is expressed in the atria and ventricular regions, thereby outlining the whole heart. In addition, the ventricle and the atrium are arranged from anterior to posterior in control hearts; however, the direction in certain PBZ-treated embryos was found to be vertical to the anterior–posterior axis. The percentage of embryos exhibiting the heart looping defect was increased in a dose-dependent manner by PBZ (Fig. 3B); the defect was observed in 8.61%±4.28%, 19.2%±6.4%, and 84%±1.8% of embryos exposed to 5 (n=162), 10 (n=150), and 20 ppm PBZ (n=140), respectively.

Misorientation and defective heart looping in zebrafish embryos after PBZ treatment.
Decreased heart rates in zebrafish embryos after PBZ exposure
To determine if PBZ exposure causes heart dysfunction, we examined the heartbeat in 10 embryos randomly selected from each treatment group (Fig. 4). In the controls (DMSO), the heart rate was 190.3±10.5 beats/min, while embryos exposed to 5, 10, and 20 ppm of PBZ had heart rates of 148.1±15.1, 137±13.7, and 120.8±3.8 beats/min, respectively. Therefore, the heart rate was significantly decreased in PBZ-treated embryos. This result indicates that PBZ exposure causes heart dysfunction, which may in turn result in embryonic lethality. We also detected substantial cell death in the heart chamber of PBZ-treated embryos at 3 dpf, which was accompanied by severe heart edema (Fig. 5).

Decreased heart rate in 3-day-old zebrafish embryos exposed to PBZ. Data are presented as the mean±SEM. Ten embryos were analyzed in each group. Bars sharing a letter are not significantly different from one another at p<0.05, as assessed by ANOVA.
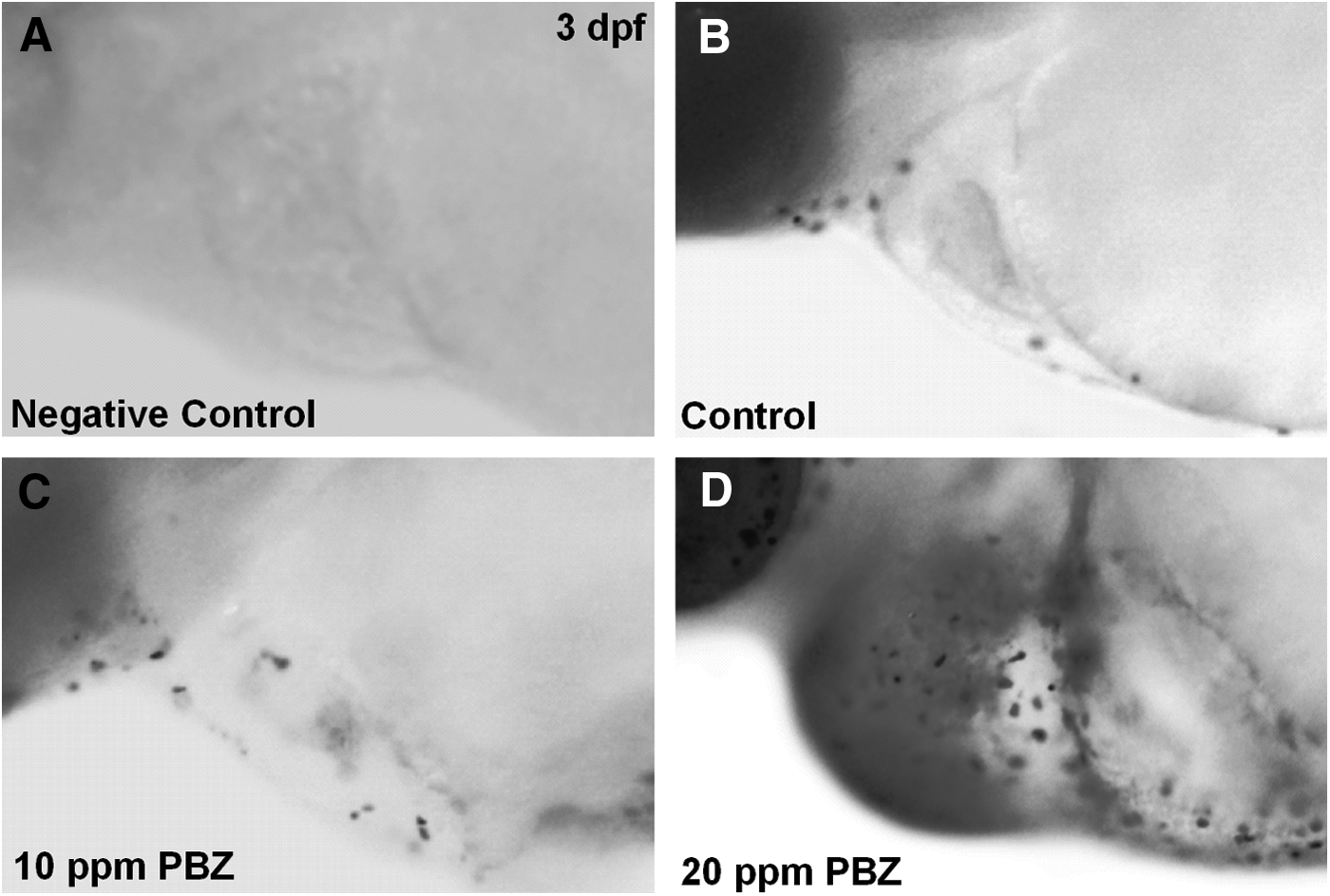
Apoptosis is induced in the pericardial chamber of zebrafish embryos exposed to PBZ. Lateral views of the area encompassing the heart chamber of 3 dpf embryos treated with
Head abnormalities in zebrafish embryos after PBZ exposure
In addition to heart morphology, we also observed that head development was impaired in PBZ-treated embryos (Fig. 6). Heads and eyes were obviously smaller in embryos treated with 5, 10, and 20 ppm PBZ as compared to controls (Fig. 6A). To analyze the changes in head components, we measured the head width, head length, and eye area in 5 dpf embryos, according to the reference lines shown in Figure 6A. The head width was measured behind the eyes, and the head length was measured from the tip of the snout to the root of the pectoral fin. We observed that the head width was no different in embryos exposed to 5 or 10 ppm PBZ as compared to controls, but it was significantly narrower in embryos treated with 20 ppm (Fig. 6B). PBZ exposure also significantly reduced the head length and eye size in a dose-dependent manner (Fig. 6C, D). These results reveal that PBZ exposure may disrupt the formation of the head skeleton, thus resulting in smaller heads.
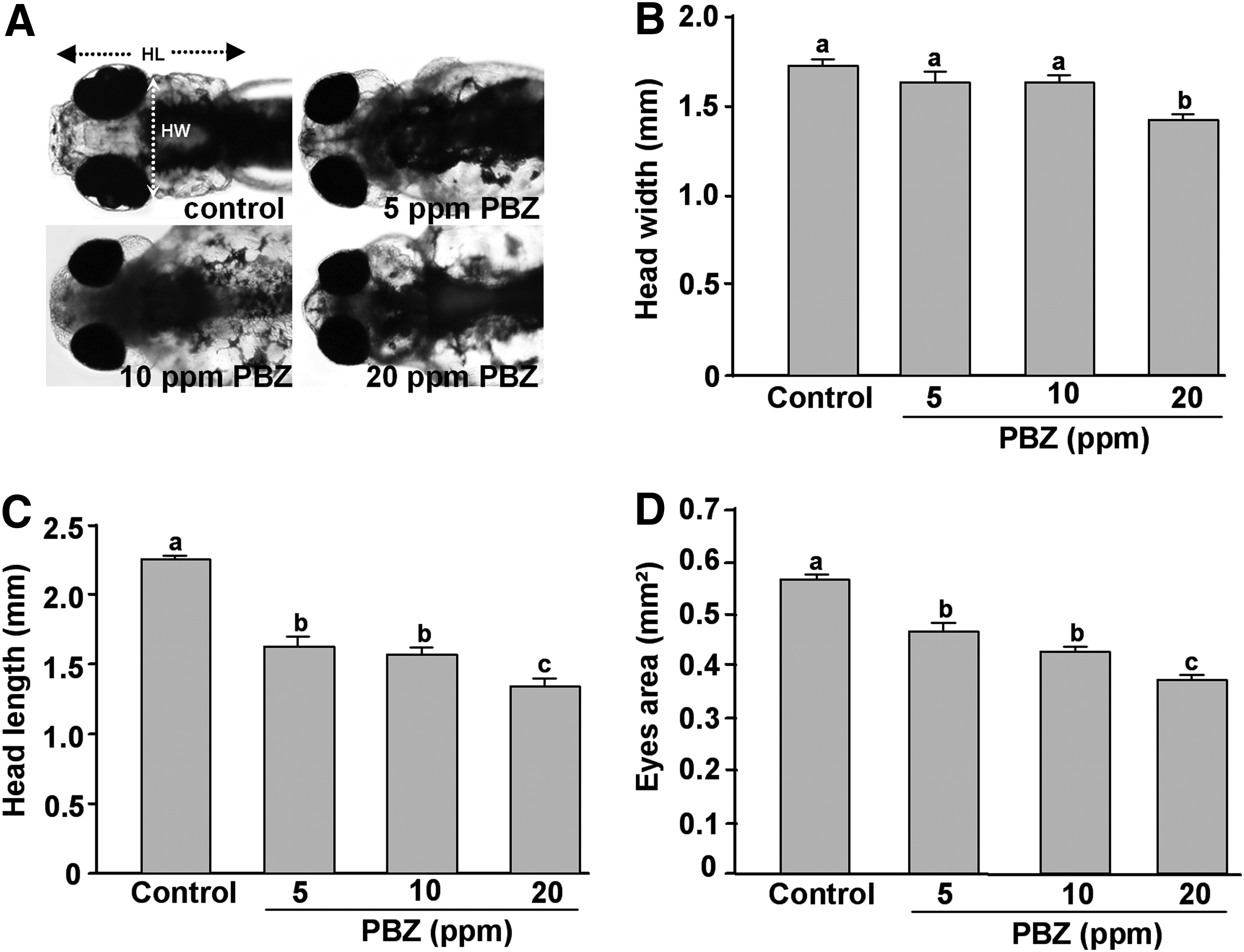
Head abnormalities in zebrafish embryos exposed to PBZ.
Defective craniofacial cartilage formation induced by PBZ treatment
We proceeded to examine whether the small heads of PBZ-treated embryos resulted from malformed craniofacial cartilage by performing Alcian blue staining of 5 dpf embryos (Fig. 7). In the control (DMSO), all elements of the pharyngeal arches (from the ventral view) and the neurocranium (from the dorsal view) were well developed (Fig. 7A). In contrast, the anterior most cartilage (arrows) was deformed, and the posterior pharyngeal arches were severely reduced, in embryos exposed to 5 and 10 ppm of PBZ. Surprisingly, most cartilage elements were absent from 38% of embryos exposed to 20 ppm PBZ, and the other 62% contained only a few craniofacial skeletal components (analyzed embryos=145). We performed a more detailed analysis by measuring the ventral ceratohyal cartilage length (CCL) (Fig. 7B), ceratohyal cartilage width (CCW) (Fig. 7C), lower jaw length (LJL) (Fig. 7D), and dorsal trabecula length (TL) (Fig. 7E), according to the reference lines shown in Figure 7A. For measuring the length and width of craniofacial cartilage in 20 ppm PBZ-treated larvae, we only measured the remaining cartilage in positions equivalent to those in the control group. We found that CCL, CCW, LJL, and TL were reduced by PBZ in a dose-dependent manner.

Defective craniofacial skeleton development in zebrafish embryos after PBZ treatment.
The migratory neural crest is abolished in zebrafish embryos exposed to PBZ
To determine how craniofacial cartilage was affected by PBZ, we first examined chondrocyte maturation by analyzing expressions of col2a1 by in situ hybridization at 5 dpf (Fig. 8A–D); col2a1 encodes an extracellular matrix protein that surrounds chondrocytes. We found that col2a1 expression was significantly reduced by PBZ in a dose-dependent manner. Expression of sox9a, an essential factor for craniofacial chondrocyte differentiation and maturation,15,16 was also reduced in the pharynx of PBZ-treated embryos at 2 dpf (Fig. 8E–H), implying an effect of PBZ on the pharyngeal primordia. We therefore examined the migratory neural crest cells that give raise to the pharynx, through hybridization against dlx2 at 36 hpf (Fig. 8I–L). This revealed that migratory neural crest cells were reduced by PBZ treatment. Taken together, these results indicate that PBZ exposure decreases the population of migratory neural crest cells in the pharyngeal area, which results in the disruption of craniofacial cartilage and malformed heads.
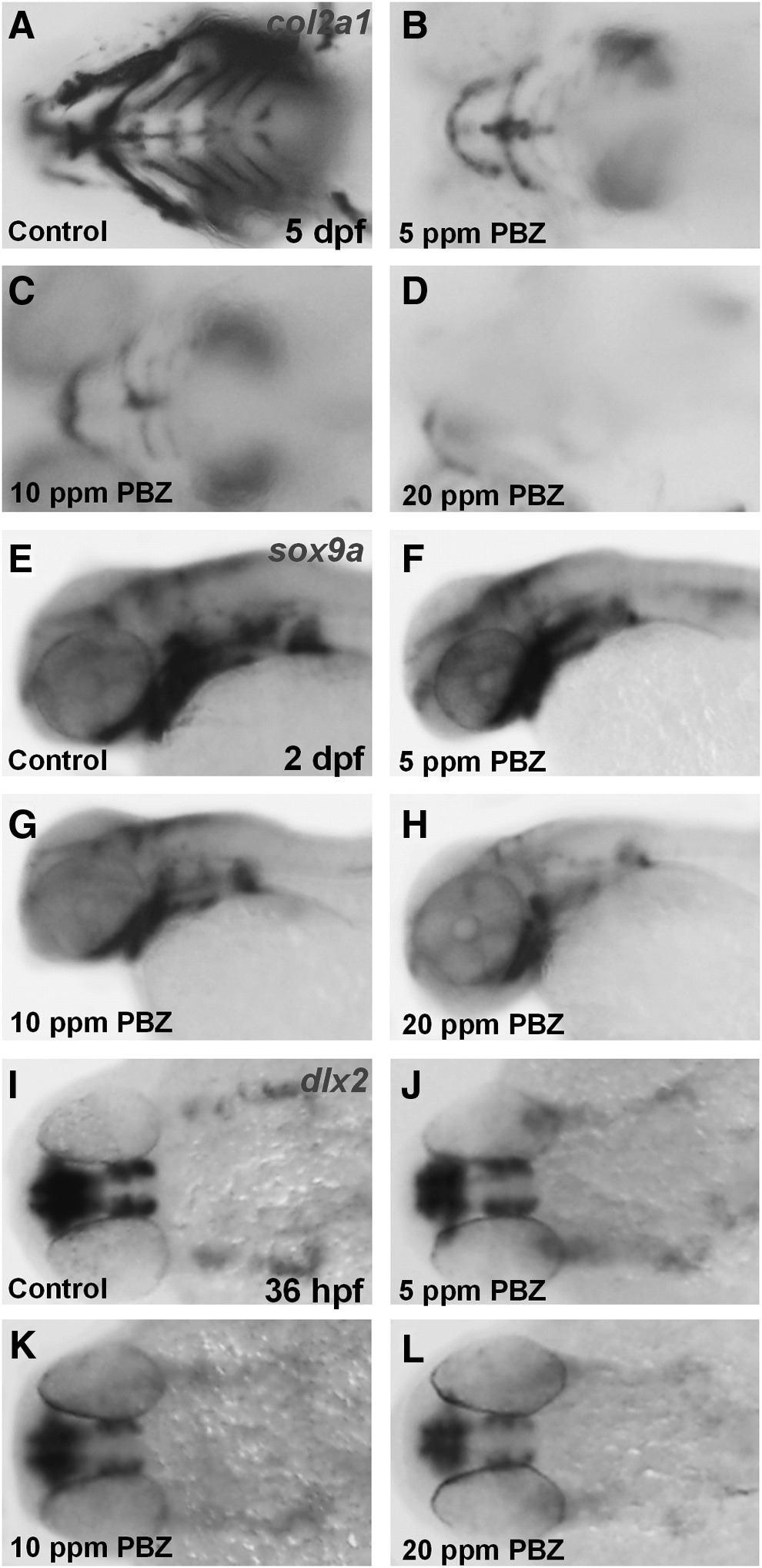
PBZ exposure affects pharyngeal cartilage development. Zebrafish embryos exposed to DMSO (control), or 5, 10, or 20 ppm PBZ were analyzed by in situ hybridization with probes against
Discussion and Conclusions
PBZ causes strong embryonic toxicity, affecting hatching and survival rate, as well as heart and head development
PBZ has been widely used in agriculture and floriculture by injection into the soil. The concentration of PBZ used by farmers ranges from tens to thousands of ppm. Rainfall can result in PBZ entering environmental water, thus potentially affecting aquatic organisms. However, only the effects of PBZ on adult aquatic organisms have been reported. In this study, we demonstrate that exposure to PBZ at a concentration as low as 10 ppm can reduce the hatching and survival rate, and retard heart and head development. As such, PBZ may have toxic effects on embryogenesis.
PBZ affects heart morphogenesis, resulting in heart failure and pericardial edema
Edema can accompany cardiovascular dysfunction, due to interconnections between the osmoregulatory function of the skin and pronephric kidney, and the circulatory function of the heart and vasculature. 23 Edema is also caused by the loss of osmotic balance for a water barrier. 24 In addition, insufficient synthesis of cardiac troponin T, an essential component of the sarcomere in cardiomyocytes, may also cause pericardiac edema in zebrafish embryos. In this study, we report that PBZ causes a heart-looping defect, raising the possibility that this looping defect may result in a slowed heartbeat associated with heart edema and cardiomyocyte apoptosis. However, it is not clear how PBZ caused the looping defects during heart development.
PBZ may affect the migratory neural crest through effects on neural crest formation
Our results have demonstrated that PBZ alters the development of craniofacial cartilage by disrupting migratory neural crest cells. The migratory neural crest is derived from neural crest cells; such cells are transient, migratory, and multipotent, giving rise to diverse cell lineages. 25 Neural crest cells are specified at the border of the neural plate and the non-neural ectoderm during gastrulation; they proceed to undergo an epithelial to mesenchymal transition during neurulation, delaminating from the neuroepithelium and migrating through the periphery, where they develop into different cell types.25–27 A gene regulatory network tightly controls the development of neural crest cells. For example, BMP signaling is involved in the induction, formation, determination, and migration of the cranial neural crest cells, which develop to form most of the craniofacial structures. Interestingly, BMP signaling is also coordinated with several different signaling pathways to ensure the proper development of the neural crest.11,28–30 Abnormalities in the neural crest can result in frontonasal dysplasia, Waardenburg-Shah syndrome, and DiGeorge syndrome.31–33 Future studies should identify whether PBZ affects any step of neural crest development or the molecules involved, and whether PBZ-treated embryos are suitable animal models for studies into the aforementioned craniofacial syndromes.
The potential molecular target of PBZ in aquatic organisms
In plants, PBZ inhibits the biosynthesis of gibberellin, a growth hormone, by preventing the formation of ent-kaurenoic acid through suppression of ent-kaurene oxidase.1,2 This inhibitory effect of PBZ gives rise to the possibility that PBZ may also affect oxidation-reduction in other species. It has been reported that PBZ disrupts liver lipid metabolism and spermatogenesis in rockfish. Interestingly, the activity of heme oxygenases may play a role in liver lipid metabolism and spermatogenesis.
In our study, we found that exposure of zebrafish to PBZ decreases the head size and causes heart failure associated with pericardial edema. It should be noted that disrupting the function of β, β-carotene-15, 15′-oxygenase, an enzyme that converts provitamin A to retinal, also results in small heads and heart edema in zebrafish. Interestingly, it has been shown that neural crest cells (which ultimately form craniofacial cartilage) may be particularly sensitive to reactive oxygen species (ROS), as they lack inherent SOD and catalase activity. 34 It has also been reported that isotretinoin treatment induces ROS production in neural crest cells, and this is related to the viability of neural crest-derived ectomesenchymal cells and normal craniofacial morphogenesis. 34 These findings may explain how PBZ affects the heart and craniofacial cartilage.
Authors' Contributions
Conceived and designed the experiments: H.J.H., A.P.A.Y., W.D.W. Performed the experiments: A.P.A.Y., W.D.W. Analyzed the data: A.P.A.Y., H.J.H., W.D.W. Wrote the article: A.P.A.Y., H.J.H., W.D.W. Corrected manuscript drafts: H.J.H., W.D.W.
Footnotes
Acknowledgments
We thank the Taiwan Zebrafish Core Facility at Academia Sinica and ZeTH (supported by the National Science Council) for providing zebrafish embryos and technical assistance. We also thank D.E. Wright for English editing. This research was supported, in part, by the National Science Council, ROC (NSC
Disclosure Statement
The authors declare no competing financial interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.