
Abstract
Abstract
Zebrafish are a powerful model organism to study disease. Like other animal models, Danio rerio colonies are at risk of pathogenic infection. Microsporidia, a group of intracellular fungus-like parasites, are one potential threat. Microsporidian spores germinate and spread causing pathological changes in the central nervous system, skeletal muscle, and other anatomic sites. Infection can impair breeding, cause other morbidities, and ultimately be lethal. Previously, detecting microsporidia in zebrafish has required sacrificing animals for histopathologic analysis or microscopic examination of fresh tissues. Here, we show that fish with microsporidial infection often have autofluorescent nodules, and we demonstrate infectious spread from nodule-bearing fish to healthy D. rerio. Histologic analyses revealed that fluorescent nodules are granulomatous lesions composed of spores, degenerating muscle, and inflammatory cells. Additional histologic staining verified that microsporidia were present, specifically, Pseudoloma neurophilia. Polymerase chain reaction (PCR)-based testing confirmed the presence of P. neurophilia. Further PCR testing excluded infection by another common zebrafish microsporidial parasite, Pleistophora hyphessobryconis. Collectively, these studies show that P. neurophilia can induce skeletal muscle granulomas in D. rerio, a previously unknown finding. Moreover, since granulomas autofluoresce, microscopic screening for P. neurophilia infection is feasible in live fish, avoiding the need to sacrifice fish for surveillance for this pathogen.
Introduction
P
Microsporidia are eukaryotic obligate intracellular parasites, and they can infect vertebrates and invertebrates. 5 First identified as a pathogen of silk worms, microsporidial research intensified following reports of human infections in 1959. 6 In humans, infection usually occurs only in patients with rare inherited immunodeficiencies or HIV/AIDS, but microsporidia can afflict immunocompetent crustaceans, insects, and fish. 4
Infections occur by microsporidial organisms parasitizing host cells. 2 Once intracellular, meronts—an asexual reproductive stage of the microsporidial lifecycle—utilize host nutrients to multiply in the cytoplasm and form spores. 2 Spores are then released by cells to move throughout the host and infect other tissues. Spores can also escape from the host to infect other organisms. A hardy exospore consisting largely of chitin allows microsporidial spores to withstand extreme environments, including decontamination by egg surface disinfectants. 2
Microsporidia can spread by spore inhalation, ingestion of spores in contaminated food, water, or fecal matter, or by living fish cohabitating in the same tank, which presumably occurs by infected eggs or sperm that are released during spawning. 2 Another important mode of transmission in D. rerio is cannibalism of deceased infected fish, resulting in ingestion of spores and meronts already present in dead tissues. 6 After consumption, microsporidia then spread through the new host to infect multiple tissues, such as the central nervous system (CNS) and skeletal muscle. Last, as with some other microsporidia, there is strong evidence that P. neurophilia is maternally transmitted. 2
Pseudoloma neurophilia infects the zebrafish CNS and peripheral nerves as well as other tissues. CNS infection can lead to inflammation and subsequent muscle wasting in the animal. 1 Spore aggregates may accumulate in the hindbrain, spinal cord, and nerve roots. Consequently, infected fish commonly have spinal deformities and abnormal body curvature. 1 However, kyphotic, lordotic, and scoliotic curvature also occurs in normal fish as a consequence of age-associated epaxial muscle degeneration, so these physical findings are not specific for P. neurophilia infection. 7
Although the CNS is the classic infectious site for P. neurophilia, this parasite can invade many other tissues, with chronic myositis also reported. Other signs of infection include emaciation, lethargy, and dorsal skin hypo- and/or hyperpigmentation. 8 However, these physical signs are not always present in P. neurophilia-infected fish. Hence, monitoring for these signs is insufficient to detect microsporidial infestation. Nonetheless, if these signs are seen in a colony, investigation for possible microsporidial infection is warranted.
Since microsporidia can cause substantial morbidity and mortality, detection of this parasite is important. In this study, we report atypical P. neurophilia infections characterized by abundant intramuscular granulomas and myonecrosis, in addition to classic CNS pathology caused by this parasite. Moreover, lesions exhibited striking autofluorescence, yielding a simple means to screen D. rerio for granulomas elicited by P. neurophilia. Thus, surveillance may be feasible in at-risk colonies or in fish manifesting signs of infection. Finally, while myositis occurs in P. neurophilia-infected fish, 9 granulomas in skeletal muscle have not been reported previously with this microsporidian. This novel finding of myonecrotic granulomas yields new insight into P. neurophilia pathology, and simultaneously expands D. rerio's utility as a granulomatous infectious diseases model.
Materials and Methods
Zebrafish care and maintenance
Wild-type (WT) Nacre fish and WIK-strain fish of the hlk, srk, and otg lines 10 were housed in a colony at 28°C on a 14/10-h circadian cycle. Fish were handled according to NIH guidelines, under protocols approved by the Institutional Animal Care and Use Committees of the University of Utah (#11-07006) and the University of Oklahoma Health Sciences Center (#12-066).
Microscopic examination and collection of infected zebrafish
Zebrafish were anesthetized using 0.02% buffered pH 7.0 Tricaine (Tricaine methanesulfonate, MS-222) and examined using an Olympus SZX12 microscope (Center Valley, PA). To try to limit infectious spread, fish with fluorescent nodules were quarantined from our colony in distinct tanks.
Cohabitation of infected zebrafish with nacre fish
Infected fish were identified as described above and housed in groups of 50–80 animals in 10-L tanks. To test whether nodules were caused by a transmissible pathogen, three groups of six healthy fish were introduced at different times into these large tanks for cohabitation with nodule-bearing fish. To facilitate identification of test fish, we used nacre strain animals, which lack the melanin pigment, thereby making them easily recognizable in the cohort. Cohabitant nacre fish were then examined by fluorescent microscopy at 3 and 6 months to assess whether they acquired fluorescent nodules.
Image capture
Fish with suspected microsporidian infection were screened using microscopes equipped with fluorescein isothiocyanate (FITC) and tetramethylrhoodamine-5-isothiocyanate (TRITC) filters. Images were obtained using an Olympus SZX12 microscope and Olympus America camera (Center Valley, PA). Image processing was performed using PictureFrame software (Center Valley, PA).
Examinations of histologic slide preparations were conducted using a Nikon Eclipse E600W microscope (Tokyo, Japan). Brightfield, 4,6-diamidino-2-phenylindole (DAPI), and FITC images were captured using a Spot Insight Firewire camera and Spot Advanced software (Sterling Heights, MI).
Histologic preparation of zebrafish tissues
Fish were fixed in 10% formalin or Dietrich's fixative, 11 processed on an automated histologic tissue processor, embedded in paraffin, sectioned, and stained using hematoxylin and eosin (H&E) (performed at ARUP Laboratories [Salt Lake City, UT] according to standard clinical laboratory protocols), Periodic acid Schiff (PAS), 12 Grocott's methenamine silver (GMS), 12 Gram [performed at ARUP Laboratories according to standard clinical laboratory protocols], Luna, 5 Trichrome,12,13 PTAH, 12 or Fungi-Fluor (Polysciences, Warrington, PA) stains. All stains were completed using standard methodology or according to the manufacturer's protocol.
Polymerase chain reaction assays
Living fish with fluorescent nodules were submitted to IDEXX RADIL (Columbia, MO) for microsporidial polymerase chain reaction (PCR) testing with proprietary species-specific primer pairs and PCR conditions. Amplicons were analyzed by capillary electrophoresis.
Results
We study mutant cancer-prone D. rerio lines with an lck:EGFP transgene. 10 These lines are on the WIK strain background and have T cell-specific GFP expression. 14 To detect T-cell cancers in these lines, we screen 1000–2000 fish on a monthly basis using fluorescence microscopy. Our cancer-prone lines, like WT zebrafish, exhibit vibrant coloring and oval body contour (Fig. 1A).
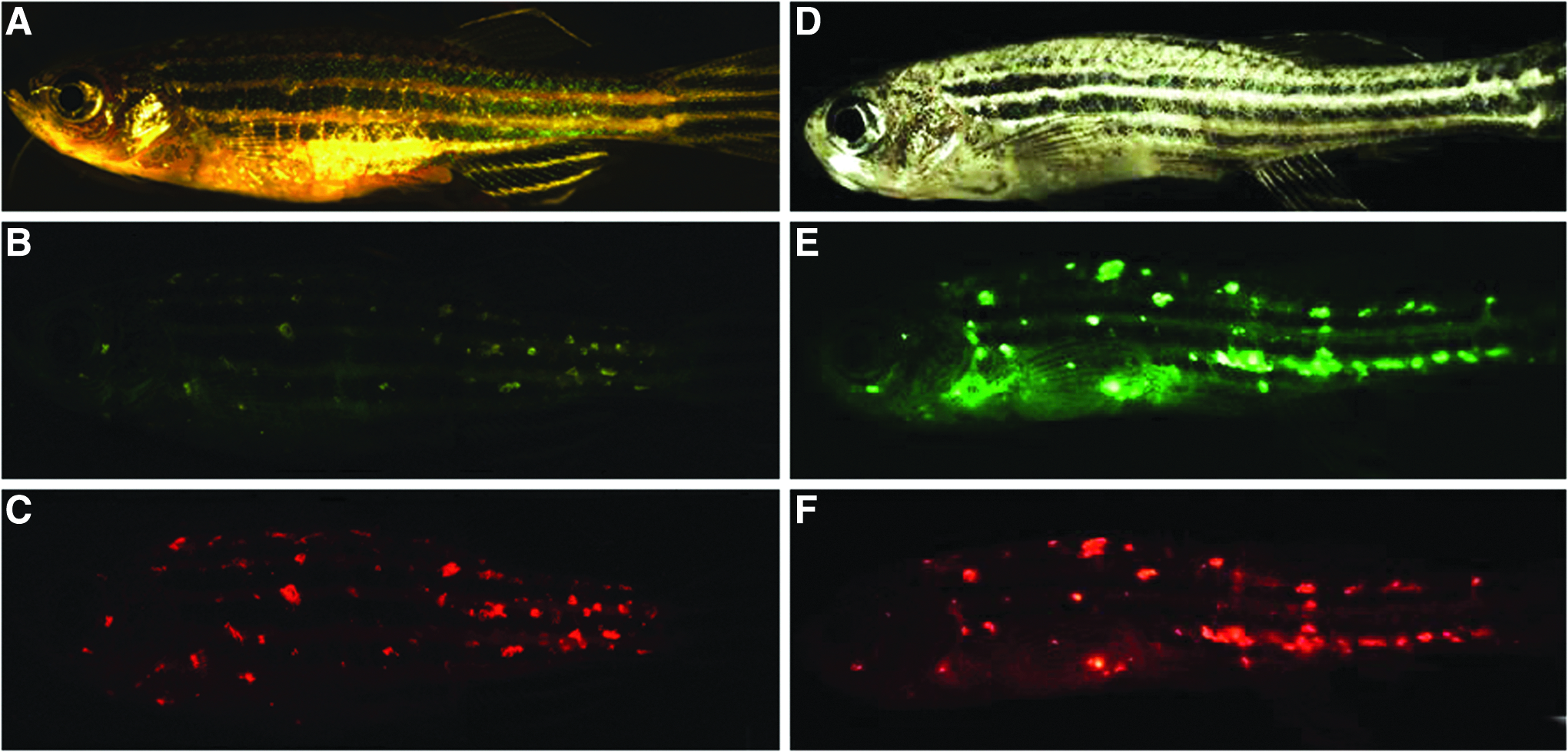
Autofluorescent nodules in Danio rerio. Microscopy demonstrates fluorescent nodules in healthy appearing zebrafish and fish displaying pathologic features.
Over a period of 3 years, encompassing repeated examinations of over 4000 fish, we identified 123 animals (∼3% of the population) that exhibited nodules emitting yellow-green fluorescence (Fig. 1B). Fluorescence was detectable at several wavelengths, but was most specific in the red (TRITC filter) spectrum (Fig. 1C). Notably, these lines do not carry transgenes for red fluorophores such as DsRed or mCherry. In this same colony, particularly in fish >1 year of age, we also observed many animals with wasting, abnormal spinal curvature, and hypopigmentation (Fig. 1D). Many of these fish also displayed green (Fig. 1E) and red (Fig. 1F) fluorescent nodules. Overall, we estimate the incidence of wasting to be ∼5% in our colony by 12 months and ∼25% of fish by 24 months. However, many fish with physical signs of infection lacked fluorescent lesions. Alternatively, occasional animals with no outward evidence of pathology harbored abundant fluorescent nodules (Fig. 1A–C).
Fluorescence had different color, size, shape, and anatomic distribution than T-cell cancers occurring in these fish. 10 Nodules were abundant in affected fish and chiefly localized to flank musculature. Lesions were also seen along the spine that, together with abnormal spine curvature, suggested CNS pathology.
The D. rerio lines where these unusual lesions were found also carry distinct, but unidentified, germline mutations responsible for their cancer predisposition. It is unlikely that three different mutant lines would all share an unrecognized immunodeficiency, as these fish have normal lifespan, fertility, and health. Nonetheless, to exclude the possibility that our cancer-prone lines might also be susceptible to an unusual and perhaps opportunistic pathogen, we explored whether other fish could also develop these lesions. To limit the potential spread of this condition in our colony, we quarantined all nodule-bearing fish in groups of up to 80 animals. Then, to test for transmission to healthy fish, we added groups of immunocompetent nacre zebrafish. 15
Because nacre fish lack melanophores, their body pigment is largely absent (Supplementary Fig. S1A; Supplementary Data are available online at www.liebertpub.com/zeb), allowing us to easily remove these test fish from the larger cohort for serial fluorescent screening. In addition, our nacre fish lack lck:EGFP, allowing us to further confirm that fluorescent nodules were unrelated to the GFP transgene. We housed three groups of six nacre fish each amidst ∼80 other nodule-bearing fish for up to 6 months, with regular microscopic evaluations. In this series, 3/18 nacre fish acquired fluorescent nodules after 3 months. Like prior fish, nacre developed abundant and vividly fluorescent lesions throughout their flank musculature, but nodules were smaller than those in our other fish strains (Supplementary Fig. S1B, C).
To investigate the identity of these nodules, we performed histologic analysis on 6 fish bearing fluorescent lesions. H&E sections revealed abundant intramuscular granulomas with surrounding chronic inflammation (Fig. 2A and Supplementary Fig. S2A). H&E stains also detected CNS lesions (upper panels of Fig. 2C). To further define lesions, we used PAS, which stains polysaccharides in the cell walls of viable fungal organisms. CNS lesions on H&E stains were avidly PAS+ (lower panels of Fig. 2C), as were intramuscular granulomas (Fig. 2E and Supplementary Fig. S2C).
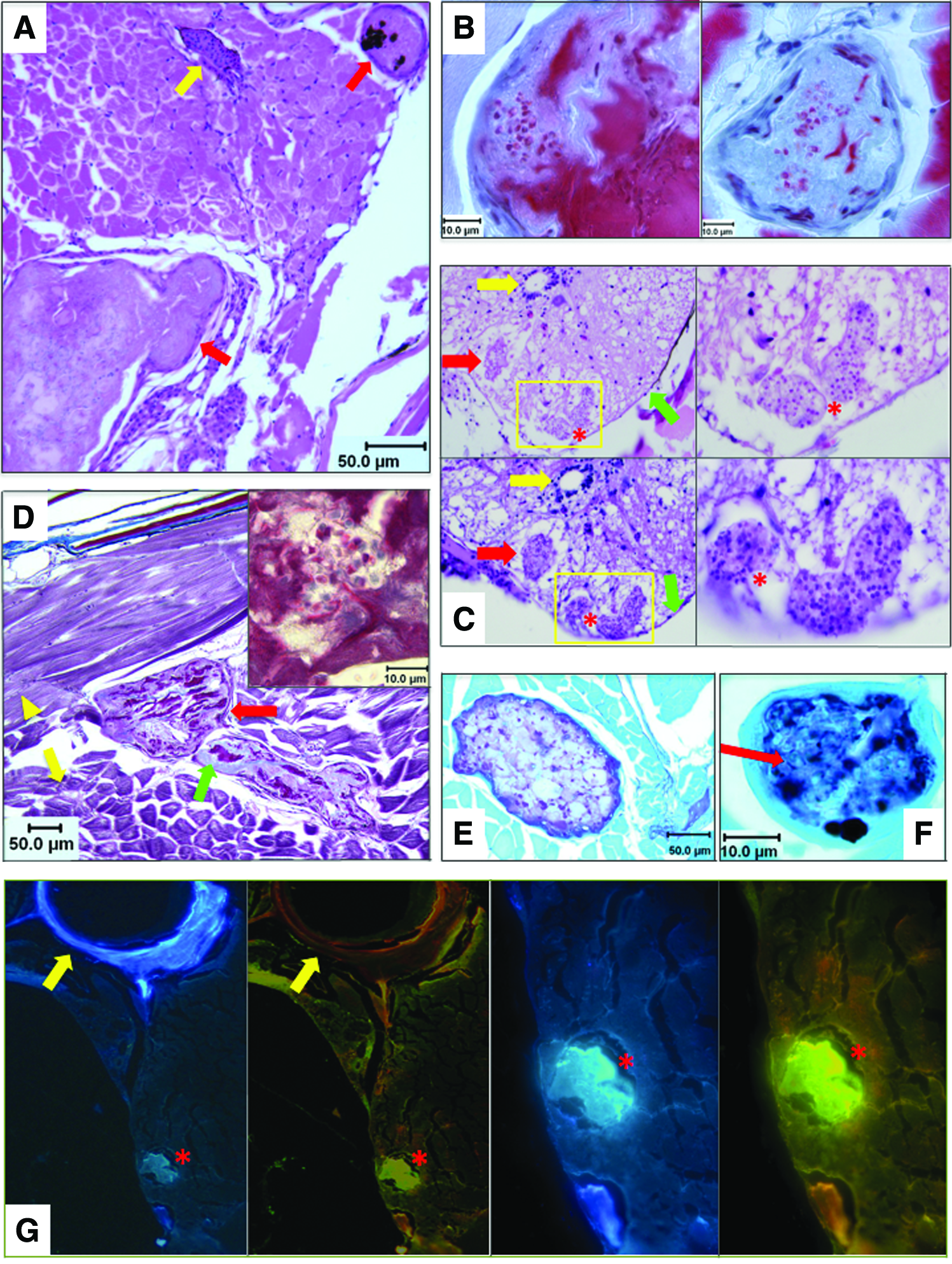
Histopathologic analysis of microsporidia-infected D. rerio. Zebrafish with fluorescent nodules and other signs of infection were analyzed with a panel of histologic stains:
Luna stains, a method that highlights chitin in microsporidian spores, detected microsporidia in skeletal muscle granulomas (Fig. 2B). 5 Luna also revealed abundant degenerating muscle in granulomas. The finding of degenerating muscle led us to perform trichrome stains, which can also highlight necrotic skeletal muscle. Trichrome stains demonstrated degenerating connective tissue (blue) and necrotic skeletal muscle (dark pink) in granulomas (Fig. 2D). To further confirm myonecrosis, PTAH stains were used, showing striated remnants of skeletal muscle and fibrin deposits in lesions (Supplementary Fig. S2D).
We also performed Grocott's methenamine silver (GMS) stains, which mark carbohydrates in cell walls of living and dead fungi, including microsporidia. 16 Granulomas were avidly GMS+ for microsporidian elements in granulomas (Fig. 2F and Supplementary Fig. S2G, H). Microsporidian spores are gram positive, and this stain highlighted the same lesions that were GMS+ (compare Supplementary Fig. S2E–H). To verify the presence of microsporidia, we used Fungi-Fluor™, which stains chitin in spores. Granulomas showed avid Fungi-Fluor positivity (Fig. 2G).
This comprehensive staining panel yielded identical findings in all six fish examined, and demonstrates that microsporidia are present in lesions of the CNS, skeletal muscle, and other sites in D. rerio with fluorescent nodules. However, it does not prove which microsporidial species caused fluorescent muscle granulomas. We suspected two microsporidians that commonly afflict zebrafish, P. neurophilia and P. hyphessobryconis. P. neurophilia, as its name indicates, has tropism for neural tissue. In addition, the CNS lesions seen by H&E and PAS stains (Fig. 2C) are classic for infection by this organism. However, muscle granulomas have not been previously reported in P. neurophilia infection, although chronic myositis can occur. 9 Alternatively, P. hyphessobryconis is known to cause skeletal muscle lesions, 17 and infections by this pathogen are prevalent in CG1 zebrafish, a strain also housed in our colony. 17
To investigate whether DNA from P. neurophilia and/or P. hyphessobryconis could be detected in infected fish, we performed PCR assays on the brain and skeletal muscle from 2 fish with fluorescent lesions using commercial testing by IDEXX RADIL (Columbia, MO). PCR failed to detect P. hyphessobryconis in the skeletal muscle of either animal, but P. neurophilia DNA was present in the brain and muscle tissue of both fish (Table 1 and Supplementary Fig. S3). Based on histologic results and PCR testing, we conclude P. neurophilia causes the CNS lesions and skeletal muscle granulomas seen in our fish, explaining our novel finding of autofluorescent nodules in D. rerio.
CNS, central nervous system; PCR, polymerase chain reaction.
To verify that P. neurophilia-induced fluorescent granulomas were not unique to our zebrafish colony, we also investigated fish from a collaborator's laboratory (MLK, Oregon State University, Corvallis, OR). Five fish from their at-risk colony were selected for histologic analysis and fluorescent microscopy. Three fish exhibited pathologic features characteristic of P. neurophilia, and in agreement with our findings, 1/3 fish with confirmed infection also had autofluorescent muscle lesions. This suggests that fluorescent granulomas are a consistent feature of P. neurophilia infection in D. rerio, and that this finding can be generalized to other laboratories.
Discussion
Microsporidia are fungal-like organisms that infect invertebrate and vertebrate species, including zebrafish. These intracellular pathogens parasitize hosts and form durable spores capable of infectious spread. Transmission occurs by ingestion of contaminated water, fecal matter, and cannibalism of diseased fish (Fig. 3A). Spores germinate in host cells and reproduce intracellularly (Fig. 3B). Microsporidia can then spread throughout the host causing disseminated infection. Physical signs associated with advanced infection include pigmentary changes, wasting, unusual body contour, and lethargy (Fig. 3C). Infestations cause morbidity and, ultimately, fish demise (Fig. 3D), conceivably leading to skewed results, inaccurate data, and loss of study animals.
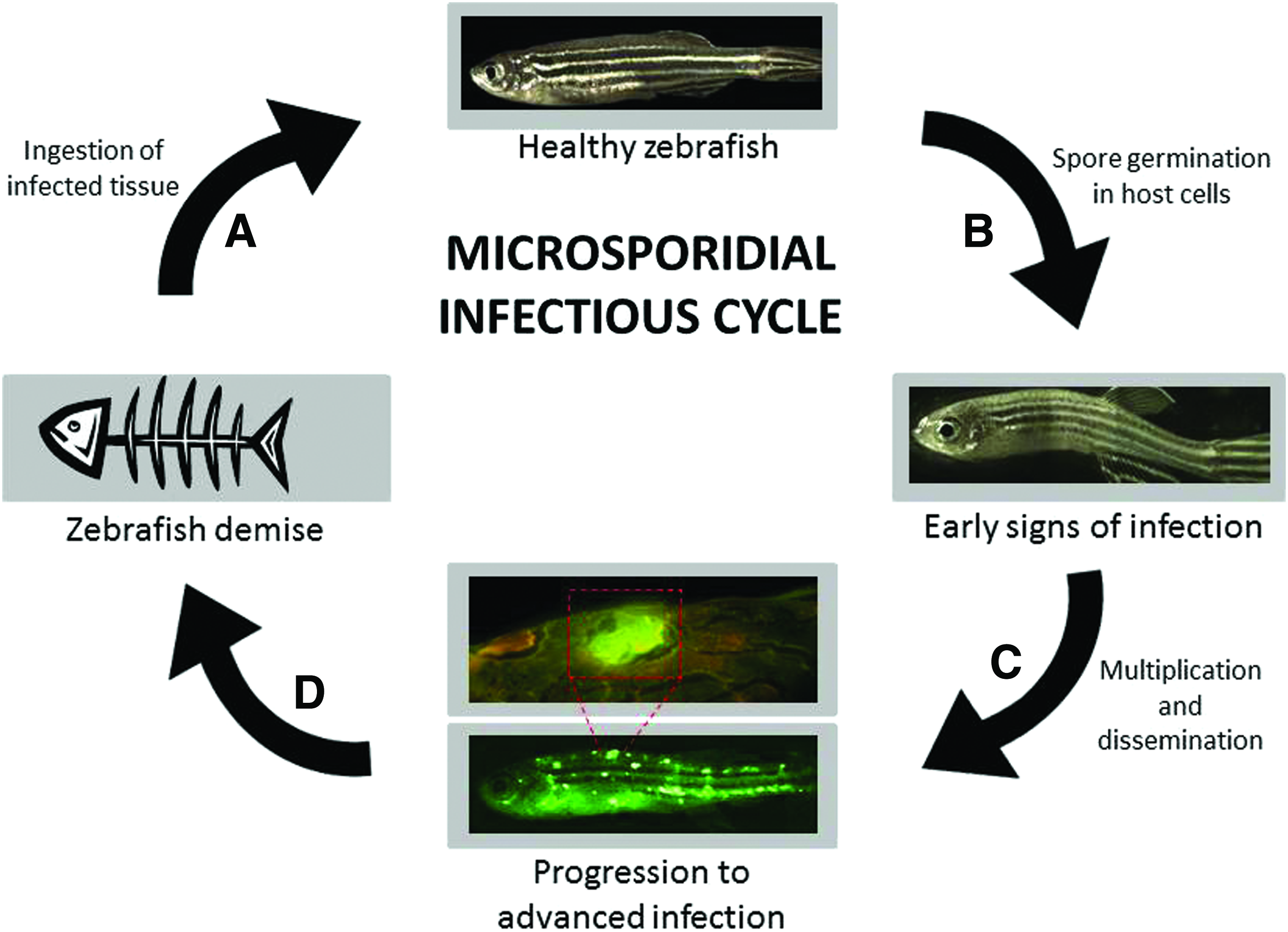
Schematic depiction of microsporidial infection in zebrafish. Infection begins when healthy D. rerio are exposed to spores in food or water, are exposed to infected eggs or sperm during spawning, or consume infected tissue from deceased fish
Our laboratory uses D. rerio with transgenic GFP expressed by T cells. We identified >100 fish with fluorescent lesions that were visible using GFP/FITC filters (Fig. 1B, E and Supplementary Fig. S1B; GFP emission spectra: 503–537 nM, FITC: 515–545 nM). Although visible with green filters, we noted fluorescence was yellow-green and distinct from green fluorescence emitted by T cells. Red filters (TRITC: 573–613 nM) highlighted lesions with greater specificity (Fig. 1C, F and Supplementary Fig. S1C). We suspected these lesions were infectious in nature, so we performed histologic analyses to determine their origin.
Lesion-bearing fish exhibited pathologic findings classic for P. neurophilia CNS infection, but also had granulomas in skeletal muscle, which is novel for this pathogen. Myonecrosis in granulomas (Fig. 2B, D and Supplementary Fig. S2D) may explain why autofluorescent nodules render infected fish easily detectable by microscopy. 18 When skeletal muscle and other tissues degrade, lipofuscin, ceroid, and other necrotic materials accumulate, and these products also have fluorescent characteristics.
Fish with fluorescent nodules usually had physical hallmarks of illness (Fig. 1D), but occasional animals did not manifest outward signs of infection (Fig. 1A), indicating granulomas can sometimes precede clinical features. In these instances, parasitic infestation was only apparent by microscopy. In addition, because fluorophores are widely used in D. rerio, other laboratories may incidentally observe similar lesions in their fish, making this finding of potential utility in that circumstance.
Histologic or DNA testing proved P. neurophilia infection in all 8 fish examined from our colony (Fig. 2 and Supplementary Figs. S2 and S3, Table 1), and this was independently verified in different fish from another laboratory. Notably, lesions contained microsporidial spores and organisms, and PCR proved that P. neurophilia DNA was present in muscle and CNS tissue from fish with fluorescent nodules. Parallel PCR testing also excluded a microsporidian, P. hyphessobryconis, with known tropism for D. rerio muscle tissue. 17 Whereas a noninvasive test to detect P. neurophilia in water has been described, 19 this method only evaluates if this microsporidian—not actual infection—is present. Examining living fish by fluorescent microscopy provides a second nonlethal means to detect this parasite. In addition, microscopy can reveal whether infection genuinely exists and has the advantage of being able to screen individual fish.
Abnormal spinal curvature is characteristic of P. neurophilia CNS infection, and CNS lesions were apparent in fish (Fig. 2C). However, scoliosis, kyphosis, and lordosis also occur in normal fish at advanced age, 7 so this is a nonspecific finding. P. neurophilia can infect skeletal muscle also, causing myositis and muscle degeneration. 9 Although chronic, multifocal myositis has been described for P. neurophilia, to our knowledge, this is the first study to report myonecrotic granulomas with this pathogen. Most strains used in our study were mutant lines prone to T-cell cancers, 10 but they have no known immunodeficiency. In fact, these fish are healthy and have normal lifespans. We note that 3/18 nacre fish housed with P. neurophilia-infected D. rerio also acquired widely distributed, although smaller, fluorescent nodules (Supplementary Fig. S1). However, zebrafish strains can show varying susceptibility to different pathogens, 20 so it remains possible that our mutant lines and nacre fish both have an atypical predilection to P. neurophilia-induced granulomas. Even so, interanimal passage confirms infectious transmission, so we conclude that fluorescent granulomas represent a communicable feature of P. neurophilia infection.
Fish were analyzed by a comprehensive histology panel consisting of H&E, Luna, trichrome, PTAH, PAS, GMS, Gram, and Fungi-Fluor stains. H&E showed granulomas and surrounding inflammation. PAS highlighted spores in CNS. Luna, trichrome, and GMS all demonstrated spores in granulomatous lesions. Luna, trichrome, and PTAH stains also revealed that degenerating muscle is the dominant histologic component in granulomatous lesions, and this feature may explain the autofluorescence seen. 18 Finally, Luna and Fungi-Fluor both detected microsporidial chitin in granulomas. In particular, Luna stains are powerful for detecting of P. neurophilia, the pathogen here. 5 Collectively, stains displayed features both characteristic of, and previously unreported in, D. rerio infections by P. neurophilia. Since this pathogen is the most prevalent microsporidian parasite of zebrafish, recognizing this variant pathology is important, as P. neurophilia afflicts many fish colonies. 2
PCR detected P. neurophilia in the CNS and muscle tissue of infected fish, while P. hyphessobryconis was absent in muscle, where this infection typically occurs (Table 1 and Supplementary Fig. S3). Whereas atypical infection by P. neurophilia is the most parsimonious explanation for our findings, there are other possible interpretations of our data. Although we ruled out P. hyphessobryconis as the source of muscle lesions, we did not exclude the possibility of other microsporidian pathogens (e.g., a novel strain or species of Pseudoloma or Pleistophora). In theory, this hypothetical pathogen might cause the muscle pathology seen with P. neurophilia merely causing coincident CNS infections. In this case, the P. neurophilia DNA detected in muscle could derive from cross-contaminating DNA from adjacent neural tissue, low levels of circulating P. neurophilia organisms/spores or circulating macrophages infected by this intracellular parasite. However, the vast diversity of microsporidia and the lack of diagnostic genotyping for the >1000 known microsporidian species preclude feasible testing of this hypothesis.
Collectively, our results from microscopy, cohabitation, histology, and molecular testing all indicate that P. neurophilia can cause both CNS infection and myonecrotic granulomas in zebrafish. Serendipitously, granulomatous lesions have vivid fluorescence, which we attribute to the autofluorescent nature of necrotic components of degenerating muscle in granulomas. This permits microscopic screening of fish for granulomas, which is more rapid and less costly than pathologic examination of individual fish. Moreover, microscopy does not require sacrificing fish for testing. Obviously, many fish infected by P. neurophilia lack granulomas, limiting the utility of microscopy as a screening tool. However, the typical physical signs of microsporidial infestation also lack sensitivity as a means of detection, so cognizance of this finding as a potential indicator of P. neurophilia infection is a potentially useful adjunct for zebrafish husbandry.
Perhaps, more importantly, these findings imply that the native zebrafish pathogen P. neurophilia may be a useful model of human microsporidial infections. D. rerio are already an important research tool for other human pathogens, such as Escherichia coli, 21 Vibrio cholera, 22 Streptococci, 23 and Staphylococci. 24 The microsporidian Encephalitozoon intestinalis afflicts HIV/AIDS and other immunocompromised patients, 25 and fumagillin is an antimicrobial agent to treat this infection. 26 Granulomas induced by P. neurophilia provide a substrate to test the fumagillin activity in zebrafish. This might salvage important D. rerio lines parasitized by this pathogen, and also establish zebrafish as a preclinical model to test new antimicrosporidial drugs. In addition, D. rerio are also used to model human granulomatous diseases caused by mycobacteria, particularly Mycobacterium tuberculosis, where native infection by Mycobacterium marinum is used as a surrogate. 27 Our results show that P. neurophilia is a native zebrafish pathogen that can elicit granulomatous reactions. As such, P. neurophilia infections provide another addition to the expanding utility of D. rerio in the study of human infectious diseases and immune responses.
Footnotes
Acknowledgments
The authors thank Diana F. Bradley, Matthew Mau, Lauren Shih, Nikolaus Trede, and Julie Hawkins for their technical and intellectual contributions to this work. They also appreciate Marcus Crim, Head of Aquatic Diagnostics, with Idexx Radil™ (Columbia, MO) for discussions regarding PCR testing of D. rerio for P. neurophilia and P. hyphessobryconis.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.