
Abstract
Abstract
Astyanax mexicanus, a teleost fish comprising both sighted river-dwelling and blind cave-dwelling morphs, is becoming increasingly used in the field of developmental and evolutionary biology. Thus, new experimental and technological tools are needed on this emerging fish model by the expanding scientific community. Here, we describe Astyanax husbandry and egg spawning habits, a prerequisite to the successful establishment of Astyanax transgenic lines. We then compare two different transgenesis methods on both surface and cave Astyanax. Both meganuclease (I-SceI)- and transposase (Tol2)-mediated transgenesis are equivalently efficient, resulting in ∼40% mosaic transgenic fish in F0. Furthermore, the transmission rate was analyzed in F1 in the case of the I-SceI method and was found to be 16%. Finally, the transgene was found stable up the F3 generation, demonstrating the feasibility of generating stable transgenic lines in Astyanax and opening a wide range of possibilities for this fish model.
Introduction
T
The emergence of the A. mexicanus blind CF as a model system is relatively recent compared to other well-established fish model species such as zebrafish (Danio rerio). However, the community of laboratories working on CF grows continuously (307 hits in Medline for Astyanax; 133 in the past 5 years). Astyanax is more and more used to explore the various aspects of evolutionary biology, including population biology,4–6 developmental biology,7–11 behavioral studies,12–22 and genetics.23–27 Some of these researches have shown a direct link between genes and specific CF phenotypes. This includes, for example, the mutations in Oca2 and Mcr1 responsible for the depigmented CF phenotype,26,28 or a mutation in the enzyme Mono Amine Oxidase, which explains part of the CF behavioral syndrome. 29
In the past few years, efforts have been made to develop resources to establish Astyanax as a suitable laboratory animal model. Major improvements include the availability of a developmental staging table, 30 developmental and adult transcriptome assemblies,31,32 cDNA libraries and an Astyanax browser, 32 and the ongoing Astyanax cavefish genome project (McGaugh et al., in preparation; www.ensembl.org/Astyanax_mexicanus/). For this fish model to enter the genome era, it is now necessary to master genome manipulation techniques to generate stable transgenic lines, for instance, carrying reporter genes or inactivated genes.
In this study, we present methodological data for the setup and running of an Astyanax facility, which allows obtaining frequent spawning, as well as protocols for establishing stable transgenic lines of A. mexicanus CF or SF. We compare the efficiencies of meganuclease- and transposase-based transgenesis strategies, two methods routinely used in well-established models, such as zebrafish and medaka (the ricefish Oryzias latipes).33–35
Materials and Methods
Facility
Laboratory stocks of A. mexicanus SF (origin: Balmorhea Spring State Park, Toyahvale, TX) and Pachón CF (origin: Pachón cave, San Luis Potosi, Mexico) were obtained from 2004 to 2006 from the Jeffrey Laboratory (University of Maryland, College Park, MD) and since then bred in our local facility. Adult fish are maintained in large tanks of 120 L (Supplementary Fig. S1A; Supplementary Data are available online at www.liebertpub.com/zeb), under a 12-h light–12-h dark cycle, in groups of 20–30 individuals (a population density that we have determined as optimal to cope with the natural aggressiveness of SF). They are fed twice a day with dry food consisting of flakes for SF (TetraMin, Europrix 3020220) and pellets for CF (Tetra Prima Discus, Europrix 3030120), plus live food like adult brine shrimp Artemia and bloodworms (Chironomidae). CF are starved for 1 week by month. The tank water is changed once a week manually with tap water (50% renewal). In our local animal facility (Gif-sur-Yvette, France), the water parameters are 25–50 mg·L−1 NO3−; 0 mg·L−1 NO2− pH 8.07; 10°<GH (Gesamt Härte, i.e., general hardness)<16°; 3° KH (Karbonat Härte, i.e., carbonate hardness), and 526 μS·cm−1 of conductivity. The water temperature is maintained at 26°C for SF and 22°C for CF. This mimics the conditions we have observed in the wild (22.3°C in Pachón cave; 22.5°C in Los Sabinos cave; 23.4°C in Tinaja cave; 26.2°C in El Nacimiento del Rio Choy; recorded in March 2009, 2011, and 2013).
Animals are treated according to the French and European regulations for handling of animals in research. SRs authorization for use of animals in research is number 91-116, and the Paris Centre-Sud Ethic Committee authorization number is 2012-0052.
Spawning
Spawning is stimulated twice a week. To this end, the temperature is switched progressively from 22°C to 26°C in CF tanks and from 26°C to 22°C in SF tanks, respectively, overnight (this is intended to simulate temperature change after rain in natural conditions). The next morning, the tanks are switched back to 22°C and 26°C for CF and SF, respectively. Moreover, boxes containing artificial plastic substrates that we call MOPS (EHFIFIX; Eheim, Deizisau, Germany) (Supplementary Fig. S1B) are added in tanks to simulate vegetation or other substrates and to protect eggs from parental predation. The fertilized eggs are collected in the morning following temperature change and are recovered from MOPS. Embryos are then raised in 90-mm Petri dishes in 30–35 mL blue water at 23°C in an incubator, a density of about 50 embryos by dish being optimal. The composition of the blue water is NaCl (1 g·L−1), KCl (30 mg·L−1), CaCl2 (40 mg·L−1), MgSO4 (160 mg·L−1), and a trace of methylene blue. The dead embryos are removed and blue water is changed every day. From 6 days postfertilization (dpf ) onward (which corresponds to mouth opening), larvae are fed with 1-day-old Artemia nauplii (supplier: INVE Aquaculture, Dendermonde, Belgium), once or twice a day. From ∼15 dpf, larvae are transferred into 1.5-L tanks (Aquatic Habitat zebrafish tank). When they reach the size of 2 cm and then 4 cm, juveniles are transferred into tanks of 20 and 40 L, respectively. They are fed Artemia nauplii plus pellets (M-0.3 Gemma Wean; Skretting, Stavanger, Norway). After about 6 months of age, they are maintained in 120-L tanks.
In vitro fertilization
To obtain eggs at the one-cell stage, we perform in vitro fertilization (IVF), adapted from Borowsky's method. 36 During the night (between 11 pm and 4 am, see Results section), behavioral observation of breeding tanks with a red headlight allows checking when fish are naturally excited in the tanks (recognizable by elevated locomotor activity and breeding stereotypies) or when some eggs start spawning. In such cases, they were fished. Males are gently held and their urogenital orifice is dried. Their abdomen is stimulated by repetitive slight pressure of fingers and the sperm is collected in a capillary tube (Harvard Apparatus GC100-10), avoiding as much as possible the presence of water. Females are held in a Petri dish and stripped by gentle pressure on the abdomen. If the eggs are mature, females spawn immediately. Egg quality is checked under a dissecting microscope (Olympus SZX2-16), and sperm is added on eggs as dry as possible. After 2 min, to avoid desiccation, a few milliliters of water is added, and then after 5 min, the Petri dish is filled with water.
Vectors
We used vectors routinely used by AMAGEN (www.celphedia.eu/en/centers/amagen), a platform laboratory (UMS for Unité Mixte de Service) specialized in model fish transgenesis (zebrafish, medaka). Astyanax promoter/regulatory sequences are now available for Pachón CF, thanks to the efforts of the cavefish genome sequencing project (see Introduction section), but they were unknown at the beginning of this study. We therefore used classical, nonubiquitous heterologous promoters (β-crystallin and cardiac actin) widely used as controls for transgenesis in other fish species. In addition, the β-crystallin promoter is biologically relevant for Astyanax, as CF eye degeneration starts with lens apoptosis.7,37 The complete maps of the two backbone vectors used in this study are shown in Supplementary Figure S2. The plasmids were designed by AMAGEN as multispecies fish transgenesis vectors using data from the literature35,38–41 and unpublished in-house tests. The combination of the various elements of the transgene was realized by gene synthesis and inserted into a pUC57 backbone by the GenScript Company. The pDEST_GFP_35H, hereby called “blue lens,” contained two I-SceI restriction sites and a CFP reporter gene under the control of zebrafish crystallin β-b1 promoter. The pDEST_ZAM12H_GFP_ref, hereby called “red muscle,” contained two I-SceI restriction sites, 41 two specific sites for the Tol2 transposase, 35 and a RFP reporter gene under the control of Xenopus cardiac actin promoter.
Injection
A binocular microscope (Leica MZ75), micromanipulator elements (Märzhäuser Wetzlar MM33 Rechts), and a pressure injection system (Eppendorf Femtojet) were used. Capillary tubes (Harvard Apparatus GC100-10) were stretched with a Narishige needle puller (PP-802) (parameters: heater level 1 at 24.8 to heater level 2 at 67.4). Both I-SceI and Tol2 injection mix contained the transgenesis vector at a final concentration of 15 ng·μL−1. The mix was completed either with the I-SceI protein (1 U·μL−1; 11362399001; Roche, Basel, Switzerland) or with Tol2 transposase mRNA (8 ng·μL−1) for meganuclease- or transposon-mediated transgenesis, respectively. In both cases, 1 nL of solution was injected directly inside the egg cell at the one-cell stage.
Screening
Depending on the vector used, injected embryos were screened for CFP or RFP fluorescence at 1 and 7 dpf under a dissecting microscope (Olympus SZX2-16) equipped with an epifluorescence source (EXFO X-cite 120-PC). Fluorescence-positive embryos were counted. The survival rate was assessed both for injected and control noninjected embryos.
Statistical analyses
The binomial test, Mann–Whitney–Wilcoxon and paired Wilcoxon test, correlation test of Spearman, chi-square and Fisher exact tests were performed with R. Significance was set at p<0.05.
Results and Discussion
General statistics of facility
Our Astyanax facility currently hosts ∼1000 SF and Pachón CF fish (plus some Molino CF and various F1 hybrids). They are distributed into seven racks (total 600 L/rack) constituted of four levels of fish tanks divided into compartments of various sizes (20–120 L) and a common filtration system per rack (Supplementary Fig. S1). Adult reproducers are kept in 120-L breeding tanks (we have six tanks for SF and seven tanks for CF) and are stimulated for spawning two consecutive days every week. Each tank contains 22–24 fish, with a sex distribution ranging from 17 females/7 males to 11 females/11 males.
The development and growth of Astyanax have been described elsewhere. 30 Below we present some general features of SF and Pachón CF adults.
Table 1 shows standard length (SL) and weight parameters for representative samples of adult individuals aged between 6 and 30 months. CF have a higher body mass than SF and in the two morphs, females are always heavier than males. Table 2 shows the sex ratio analyzed at adult stage (>6 months). Forty-eight percent of SF and 40% of Pachón CF sexed from random tanks are females. These ratios are statistically not different from the 50:50 sex ratio. Outcrosses of Molino or Pachón males with SF females seem to lead to a 50:50 sex ratio, even if the sample sizes are small to be fully conclusive. Conversely, outcrosses of Pachón females with either SF or Molino males are devoid of F1 male progeny. Again, the sample sizes are small, but this observation might suggest that Pachón CF are in the early stages of speciation. 42 More work will be necessary to support this hypothesis.
The right part of the table gives statistical comparison (Mann–Whitney test).
CF, cavefish; SF, surface fish.
, p<0.001.
p-values correspond to comparisons between female ratios and a 50% theoretical ratio (Binomial test).
, p<0.05.
Sexual dimorphism
An important issue in the breeding facility is to discern males from females. Three main dimorphic characters, shared by SF and CF, were described previously. 43
First, a slight difference in body shape was recorded. Indeed, the body depth of females is generally higher and the abdomen of males shows a concave curve just anterior to the anal fin (Fig. 1A, D, G, and J). This criterion serves as a good proxy for sex identification.

Sexual dimorphism in Astyanax mexicanus.
Second, the anal fin of females had a different shape and was longer, and the male anal fin was more opaque. 43 On fished animals, we find this criterion less reliable than the presence of denticles on the anal fin (see next paragraph). On a swimming animal, the whole body shape was far easier to observe and at least as reliable as the fin shape. Therefore, we hardly used this criterion in our day-to-day practice.
Third, males harbor denticles on their anal fin (Fig. 1A–L). It has been hypothesized that these denticles help the male to hook the female during mating and to reduce sperm dilution before egg fertilization. To document this anatomical feature, we counted anal denticles. The denticle number on male anal fins is highly variable (from 7 to 78 on sampled individuals). Interestingly, ages or sizes being equal, CF have more denticles than SF (Fig. 1M). Moreover, the denticle number is correlated with body length (Spearman correlation test; p=0.038 for SF and p=0.020 for CF), but not with age or weight, both in CF and SF. Most crucially, every male carries denticles (n=38 SF and n=22 CF). However, denticles were never observed on females (n=43 SF and n=11 CF), all identified a posteriori and unambiguously by the presence/absence of eggs in their abdomen. Interestingly, such secondary sexual character has been reported for other teleost species, including other characids44–47 and also far-related species like medaka (O. latipes). 48
Breeding colony
From 2011 to 2013, 642 SF and 385 Pachón spawns were recovered and counted. Altogether, they represent a total of ∼58,000 SF eggs and 21,000 CF eggs. Forty-three percent of SF eggs and 42% of CF eggs were viable; these rates being constant along the years (Supplementary Fig. S3). More eggs per spawn were recovered from SF (90 eggs/spawn) than from Pachón CF (54 eggs/spawn). This striking difference is mainly due to the way we harvest the eggs. To protect the eggs from adult predation during the course of spawning, we provide both groups with artificial plastic containers and substrates (Supplementary Fig. S1B–D). If such a system is provided, SF will indeed lay their eggs preferentially in the containers. In contrast, CF seem to be less aware of the presence of the device. Hence, they tend to lay their eggs mainly aside the provided substrate, resulting in a significant part of the eggs being eaten before they can be recovered.
When considering viable eggs (recognizable by their transparency and their initial correct development/segmentation/gastrulation), about 80% survive after hatching (36 hpf; no difference between SF and CF). According to our protocol to grow larvae (see Materials and Methods section), 18% of SF larvae and 36% of CF larvae reach adulthood. Concerning SF, aggressiveness is responsible for most of the juvenile mortality. As for CF, we feel that the changes of water volume in which they are kept are the critical steps.
Concerning the spawning rhythm (circadian, seasonal, and lunar), our observations over a 3-year period lead to the following conclusions. Both SF and CF spawn during the dark period, as deduced from the hour of fertilization estimated according to the developmental stages of the embryo collected the next morning. 29 Thus, a spawning peak occurs in the middle of the night, with some additional eggs being produced a few hours before the light is turned on (Fig. 2A). On average, SF and CF spawn at 1:22 am and 1:25 am, respectively. There was no correlation between the quality or the quantity of eggs per spawning and the spawning hour (data not shown). No correlation was found between spawning efficiency and either the seasonal or lunar cycle (Fig. 2B, C). Finally, the quantity (data not shown) and quality (Supplementary Fig. S3) of eggs per spawn were constant along the year.
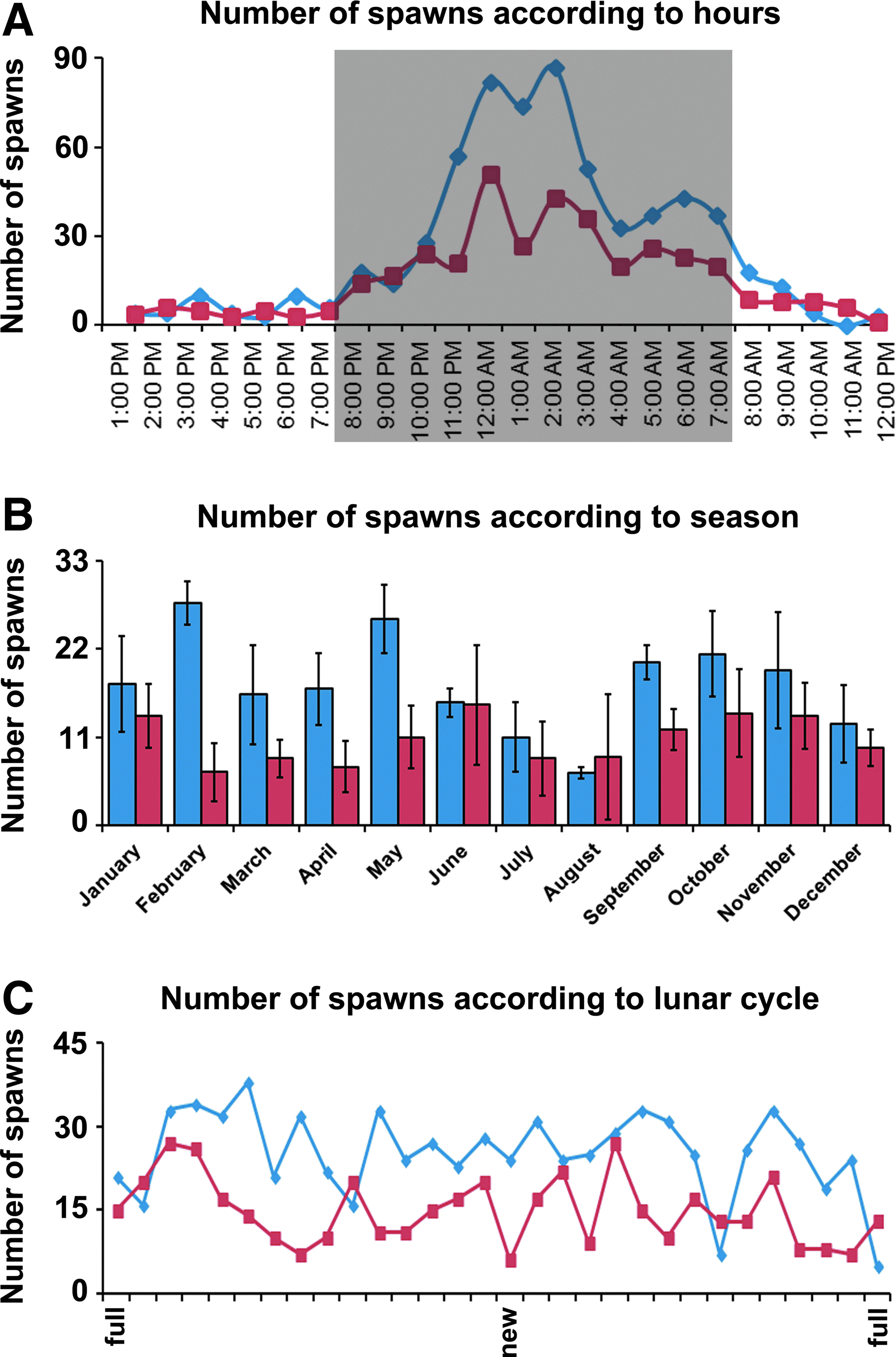
Spawning statistics according to circadian, seasonal, and lunar cycles.
Such qualitative and quantitative records on spawning habits in Astyanax SF and CF are a prerequisite to develop transgenesis techniques. The most reliable transgene expression and offspring transmission are achieved when one-cell stage eggs are injected. Therefore, based on our knowledge of Astyanax spawning cycle, IVF was deemed the best strategy to maximize the chance to get one-cell stage eggs. Hence, the eggs were harvested in the middle of the night when the fish display the strongest spawning behavior and when females are naturally excited by collective behavior. Their oocytes are mature and fertilizable at this time of the day. Before transgenesis, the females were stripped and IVF was carried out (see Materials and Methods section) to obtain large quantities of synchronously fertilized eggs.
Transgenesis in Astyanax using meganuclease
We first evaluated I-SceI-mediated transgenesis in SF, using the blue lens vector (Fig. 3A; CFP reporter gene under the control of zebrafish β-b1-crystallin promoter). Injections were performed in 134 eggs at the one-cell stage. Seventy-seven survived to 36 hpf (57%, similar to uninjected control eggs obtained by IVF: 60%). In the days following injection, 37.4% of the F0-injected embryos showed CFP expression in the lens (Fig. 3I), uni- or bilaterally, and with various degrees of mosaicism. The CFP reporter expression in the lens was weak at 36 hpf and increased with time (Fig. 3C, D, 10 dpf ). As expected and similar to zebrafish (F. Sohm and L. Legendre, personal observations), CFP was exclusively observed in the lens, suggesting that the zebrafish orthologous sequences can advantageously be used for Astyanax transgenesis if Astyanax sequences are unavailable. Twenty-five F0 CFP-positive animals (7 females and 18 males) survived to adulthood.

Meganuclease- and transposase-mediated transgenesis in Astyanax.
Among the F0 CFP-positive fish, potential founders were screened using IVF. Only males were tested because it is easy to obtain sperm, whereas it is more difficult to obtain mature oocytes from one specific female (see end of previous section). Twelve F0 CFP-positive males were tested. Of those, two were found to transmit the blue lens transgene to their offspring (2 males out of 12 tested: 16.6%). Some founders transmitting the transgene at a low frequency might have been missed out as only 100–300 eggs were screened per male. However, such inefficient transmitters would be of low practical use to establish a line and can therefore be safely ignored. The two blue lens founders were then further analyzed.
The first male founder showed a 23.6% transgene transmission rate from F0 to F1 (n=1267 eggs analyzed). In the F1 offspring, slight ectopic expression of CFP was observed at the basis of pectoral fins, probably due to the integration site of the transgene in the genome. In F2 (obtained by a cross between two F1 individuals), 69% of the fish had CFP-positive blue lenses (n=310). This rate is statistically coherent with one integration site in the genome (in theory, 75% of positive larvae expected in a cross resulting from two heterozygous parents; Fisher test, p=0.1285), although we cannot rule out the possibility of two closely linked integration sites. In F3 (obtained by a cross between two F2 individuals), 77.4% of larvae showed CFP-positive lenses (Fig. 3E). In all cases, CFP expression was homogeneous in the lens. This demonstrates that this blue lens Astyanax SF line is stable, with no extinction detected up to the third generation.
The second male founder showed a 14.6% transgene transmission rate to its F1 progeny (n=410 eggs analyzed). Interindividual variations in CFP lens expression were detected in F1. Moreover, only 44.8% of positive larvae (n=618) were observed in F2 (obtained by a cross between two F1 individuals). In this case, statistical analysis would suggest that there was less than one integration site (Fisher test, p<0.0001), which is biologically impossible. Thus, we have to hypothesize that epigenetic phenomena occurred and inhibited transgene expression. This hypothesis is corroborated by observations of strong interindividual variations of CFP expression in F2 (like in F1) and of only 51.4% of F2 offspring showing CFP in their lenses (again, different from the expected 75%; Fisher test, p<0.0001).
Pachón CF eggs were also injected with I-SceI meganuclease and the blue lens vector. The CF lens starts degenerating around 40 hpf.7,37 Before this stage, it remains relatively healthy. Unfortunately, this narrow time frame between screening (36 hpf ) and degeneration (40 hpf ) was insufficient to allow reliable detection of crystallin-CFP expression in injected Pachón CF. None but a few barely visible CFP-expressing embryos were observed (n=153 eggs). Most probably, the early lens degeneration in CF prevented the production of enough CFP to achieve detectable levels of fluorescence. This hypothesis is indeed reinforced by our observation of CFP fluorescence in the lens of transgenic CF×SF F1 hybrid larvae, which develop small but functional eyes and harbor a normal lens (data not shown).
To test transgenesis in CF, we therefore used a red muscle vector, in which a RFP reporter gene is under the control of the Xenopus cardiac actin promoter (Fig. 3B). This vector was coinjected with I-SceI meganuclease in 1565 eggs at the one-cell stage. Four hundred ninety-nine survived to 36 hpf (=59% survival) and 42.4% of the F0-injected larvae showed mosaic RFP expression in their muscle myotomes at 36 hpf (Fig. 3F, G, H, and I). This percentage of positive embryos in F0 for CF with the red muscle transgene is statistically identical to the percentage of 37.4% found in F0 for SF with the blue lens transgene (Mann–Whitney test, p=0.2361). However, in contrast to the crystallin promoter, while the cardiac actin promoter behaves adequately when expressed in zebrafish (L. Legendre and F. Sohm, personal observations), it displays a weak activity and altered pattern in Astyanax. In particular, no RFP expression was detected in the heart of Astyanax CF at 7 dpf. Altered transgene expression is not uncommon when using orthologous promoters. Other less likely explanations might be high mosaicism in F0 (not obvious earlier in development). Alternatively, Astyanax larvae being bigger and thicker than zebrafish larvae could prevent in toto visualization of weak RFP fluorescence signals.
Taken together, these data suggest that meganuclease-mediated transgenesis is efficient in Astyanax SF and CF and can be used in the future to generate stable transgenic lines.
Transgenesis using Tol2 transposase and comparison with meganuclease
To evaluate Tol2 transposase-mediated transgenesis in Astyanax, we used the red muscle vector, which also contains Tol2 transposition sites (Fig. 3B and Supplementary Fig. S2B). As the aim was to compare the efficiency with I-SceI meganuclease, this method was only tested on CF. The red muscle vector was coinjected with Tol2 mRNA in 639 CF eggs at the one-cell stage. 46.5% of F0 larvae had RFP-positive myotomes at 36 hpf (Fig. 3I) displaying the same specific pattern described above. This is very similar to the results achieved with I-SceI (42.4% positive F0, Fisher test, p=0.3307). Of note and importantly, these rates are also in the same order as for the zebrafish and medaka models, with 58% (zebrafish), 76% (medaka), and 75% (zebrafish) of positive embryos in F0, after coinjection of a circular vector with I-SceI or Tol2, respectively.34,41,49
Survival rate wise, both methods perform roughly the same: 61% of I-SceI/red muscle-injected CF embryos survived at 36 hpf (n=1818), which is virtually identical to the 59.5% of noninjected surviving embryos (n=839, eight independent experiments/egg batches, paired Wilcoxon test, p=0.1953). The survival rate of Tol2-injected CF eggs is slightly lower (38.5%, n=1460), but not significantly different from noninjected eggs (61.4%, n=776, paired Wilcoxon test p=0.4316). Furthermore, no global increase of malformation rate was observed after injection. This is coherent with our zebrafish transgenesis experience where both methods perform as well as each other (F. Sohm and L. Legendre, unpublished data). Taken together, these results indicate that both I-SceI and Tol2 methods perform equally well in Astyanax, are not deleterious for embryo development, and are suited for routine transgenesis protocols.
Conclusions
We provide here, practical elements to establish a productive Astyanax facility. Indeed, a healthy breeding colony is the prerequisite for this emerging model to enter the next step of its development, that is, to enter the genome era. The development of transgenesis and genome editing tools will require high-quality eggs obtained in fair quantity on a regular basis. This aim can only be achieved if the zootechnical aspects of reproduction and maintenance are fully mastered. While our work presents a number of recipes to set a breeding facility, room for improvement remains, in particular, concerning the production of eggs, on demand, by breeding couples such as zebrafish.
We have tested two transgenesis methods. Taken together, our data show that I-SceI meganuclease- and Tol2-mediated transgenesis are both efficient in Astyanax. This opens the possibility for the community to use these methods to generate stable Astyanax transgenic lines. In the course of our work, we also generated an Astyanax SF stable line expressing CFP in the lens. This line is readily available upon request and might be of interest, for example, for lens transplantation purpose or to study eye development. Zebrafish β-b1-crystallin promoter allows for a tight control of the CFP expression in Astyanax as it does in zebrafish 39 and medaka (F. Sohm and L. Legendre, unpublished data). It appears that at least in some favorable cases and despite the important evolutionary divergence time (100–200 Ma) between characiforms such as Astyanax and cypriniforms such as zebrafish,50–53 the use of zebrafish promoter sequence/regulatory elements could circumvent the lack of available Astyanax regulatory sequences. Still, this solution has some limitations in terms of expression pattern fidelity and will require beforehand validation of each zebrafish promoter used that way. However, this limitation applies to any heterologous promoter used in transgenesis, whatever the host and donor species. In the next step, it will be essential to further develop the next-generation technologies for gene editing, such as those using TALEN or CRISPR-cas, in our Astyanax model.
Footnotes
Acknowledgments
The authors thank Magalie Bouvet for taking care of our Astyanax colony, Jacques Callebert and Didier Casane for advice with statistical analyses, all members of the AMAGEN platform and particularly Joanne Edouard for help, and FRM (Fondation pour la Recherche Médicale, ING20140129350) for financial support to Y.E. Our work is supported by ANR (Agence Nationale pour la Recherche) grants [ASTYCO] and [BLINDTEST] to S.R.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.