
Abstract
Abstract
A cytogenetic analysis was performed on four populations of Hypostomus strigaticeps from the Paraná River basin, Brazil. Two populations were collected from the large channel river at the Itaipu reservoir area and the other two were from the upper stretches of tributaries of the Paraná River. All populations showed 2n=72 chromosomes (12m+12sm+18st+30a), intra- and interpopulation 18S rDNA site polymorphisms (two to three acrocentric chromosome pairs), and multiple 5S rDNA sites in three chromosome pairs (4, 21, and 28). C-banding revealed heterochromatin located in the centromere and pericentromere regions of most chromosome; however, large heterochromatic blocks (CMA3−/DAPI+) on the long arm of acrocentric chromosomes identified intra- and interpopulation polymorphism. The amount and distribution of heterochromatin seem to be correlated to biogeographical characteristics of H. strigaticeps along the Paraná River. Morphometric results also showed diversity among the populations, suggesting phenotypic plasticity of this species. Evolutionary, taxonomy, and biogeographical approaches with regard to H. strigaticeps and interrelationships in Hypostomus are discussed.
Introduction
L
Concerning the cytogenetic approach, Hypostomus has shown a large variation in diploid number, from low values of 2n=54 chromosomes for H. plecostomus 7 (however, this number has been questioned because these specimens might be misidentified) to 2n=84 chromosomes, as reported for Hypostomus perdido Zawadzki, Tencatt, Froehlich (2014), described previously as Hypostomus sp. 2-Rio Perdido (MS). 8 Previous cytogenetic studies in Hypostomus suggested a diverging trend in the chromosomal evolution of this group. Karyotypic diversity of the genus has shown, besides the numerical variability, intra- and interspecific structural chromosomal diversity, suggesting that chromosomal rearrangements such as centric fissions and pericentric inversions were the major mechanisms involved in the diversification of this group.9–12
Related to the nucleolar organizer regions (NORs), simple NORs are considered basal characteristic of Loricariidae 11 ; however, Hypostomus has demonstrated variations in the number and position of these regions. Karyotype diversification in the genus Hypostomus provided information to establish some biogeographic and phylogenetic correlations. Based on diploid number/karyotype formulae and NORs patterns, a correlation between these data and the hydrographic distribution of the species of Hypostomus throughout the different South American basins was reported; the species from northern basins have a smaller diploid number (2n=64 chromosomes) and single NORs, whereas species from southern basins have higher diploid numbers (2n=66–68, 2n=70 and above) and, in most cases, multiple NORs. 13
The heterochromatin pattern has played an important role in the karyotypic diversification of fish, mainly involving differentiation of sex chromosomes14–17 and karyotype differentiation,18,19 among others. Despite a few studies about the distribution and composition of heterochromatin in species of Hypostomus, analyses of these segments have contributed to different evolutionary approaches.20–22 These studies showed that heterochromatin presents heterogeneity in its distribution and composition in species of Hypostomus and is likely to represent an important marker for taxonomic and chromosomal evolutionary studies in this group.
Hypostomus strigaticeps from the Piracicaba River was described in 1907 23 and its distribution is limited to the Tietê River basin (SP). 6 However, phylogenetic and phylogeographic analyses in different populations of H. strigaticeps from the Paraná River basin (from Mogi-Guaçu, Tietê, Paranapanema, and Peixe rivers) using mtDNA (ATP synthase sequences) showed that these specimens belong to the same species (H. strigaticeps), refuting the hypothesis of cryptic species and, by phylogeographic analysis, the authors concluded its distribution among other subbasins of the upper Paraná River. 24
Correlations of environment factors, such as flow and no-flow conditions, with morphometric variabilty have already been reported in Cyprinella venusta.25,26 As H. strigaticeps inhabits medium to large river channels, the present article compared, through means of cytogenetic and morphometric analysis, two populations of medium rivers and two populations of large river channels to identify possible interpopulational chromosomal markers supporting the environmental and evolutionary trends.
Materials and Methods
Four populations of H. strigaticeps were collected and deposited in the Coleção Ictiológica do Núcleo de Pesquisas em Limnologia, Ictiologia e Aquicultura (NUPELIA) from the Universidade Estadual de Maringá, Brazil. The locality from where the specimens were collected and voucher number are summarized in Table 1.
Population downstream from the Iguassu Falls.
All specimens were anesthetized and sacrificed by an overdose of clove oil. 27 Chromosome preparations were obtained from the anterior kidney. 28 AgNORs were evidenced by silver nitrate impregnation. 29 Heterochromatin regions were determined following the C-banding technique, 30 with staining modifications. 31 Physical mapping of the 5S rDNA and 18S rDNA sequences was carried out by fluorescence in situ hybridization (FISH) 32 and modifications, 33 with probes obtained from Leporinus elongatus 34 and from Prochilodus argenteus, 35 respectively. Hybridization was performed under high-stringency conditions (77%). Probes were labeled by nick translation with digoxigenin-11-dUTP (5S rDNA) and biotin-16-dUTP (18S rDNA) (Roche®). Detection of signals was performed with antidigoxigenin-rhodamine (Roche®) for the probe of 5S rDNA and amplified avidin-FITC with biotinylated anti-avidin (Sigma) for the probe of 18S rDNA, being the chromosomes subsequently counterstained with 4′-6-diamin-2-phenylindole (DAPI, 50 μg/mL). Double staining was carried out with chromomycin A3 (CMA3) e DAPI 36 fluorochromes. Metaphases were photographed using a BX 61 epifluorescence microscope, coupled with an Olympus DP 71 digital camera with the DP Controller software 3.2.1.276. The chromosomes were classified and organized into metacentric, submetacentric, subtelocentric, and acrocentric. 37
A total of 20 specimens of each population were used for morphometric analyses: the Atlântico Stream (10 males and 10 females), Piquiri River (9 males, 6 females, and 5 not eviscerated), Paraná River (5 males, 6 females, and 9 not eviscerated), and Iguassu River (7 males, 4 females, and 9 not eviscerated). The measurements were taken from the left side of the sample using a pachymeter. 38 The morphometric parameters were the standard length, axial length, total length, predorsal length, length of head, cleithral width, depth of head, length of snout, orbital diameter, interorbital width, length of dorsal spine, length of base of first dorsal fin, interdorsal length, thoracic length along median line, length of pectoral spine, abdominal length along median line, length of the first ventral ray, postanal peduncular length, and depth of caudal peduncle. For a better interpretation of the morphologic data, the values were subjected to the canonical variable analysis (CVA) using the free software PAST version 2.13, to create a representative graph of the measurements.
Results
The cytogenetic analysis of the four samples of H. strigaticeps revealed a diploid number of 72 chromosomes and a karyotype formula of 12m+12sm+18st+30a for all populations, without chromosomal differentiation between the sexes (Fig. 1). The populations from the Paraná and Iguassu rivers presented few individuals with a size heteromorphism in pair 24 of acrocentric chromosomes, consisting of one large acrocentric chromosome and one small (Fig. 1c, d).

Giemsa-stained karyotypes of Hypostomus strigaticeps populations from
In populations from the Piquiri River and Atlântico Stream, AgNORs sites were visualized in the terminal position on the long arms of two pairs of acrocentric chromosomes (pairs 22 and 23) and one homologue of pair 24 (Fig. 2a, b). In the populations of the Paraná and Iguassu rivers, AgNORs were also located in the terminal position on the long arm of acrocentric chromosomes (pairs 22, 23, and 24) (Fig. 2c, d). These regions were confirmed by 18S rDNA-FISH (Fig. 3) and were CMA3+(Fig. 4). The 5S rDNA in situ hybridization revealed multiple cistrons for all populations located in the pericentromeric region on the short arm of pair 4, coincident with a heterochromatin block, and in the terminal position on the short arm of subtelocentric 21 and acrocentric 28 (Fig. 3).

C-banded karyotypes and AgNORs bearing pairs of H. strigaticeps populations from
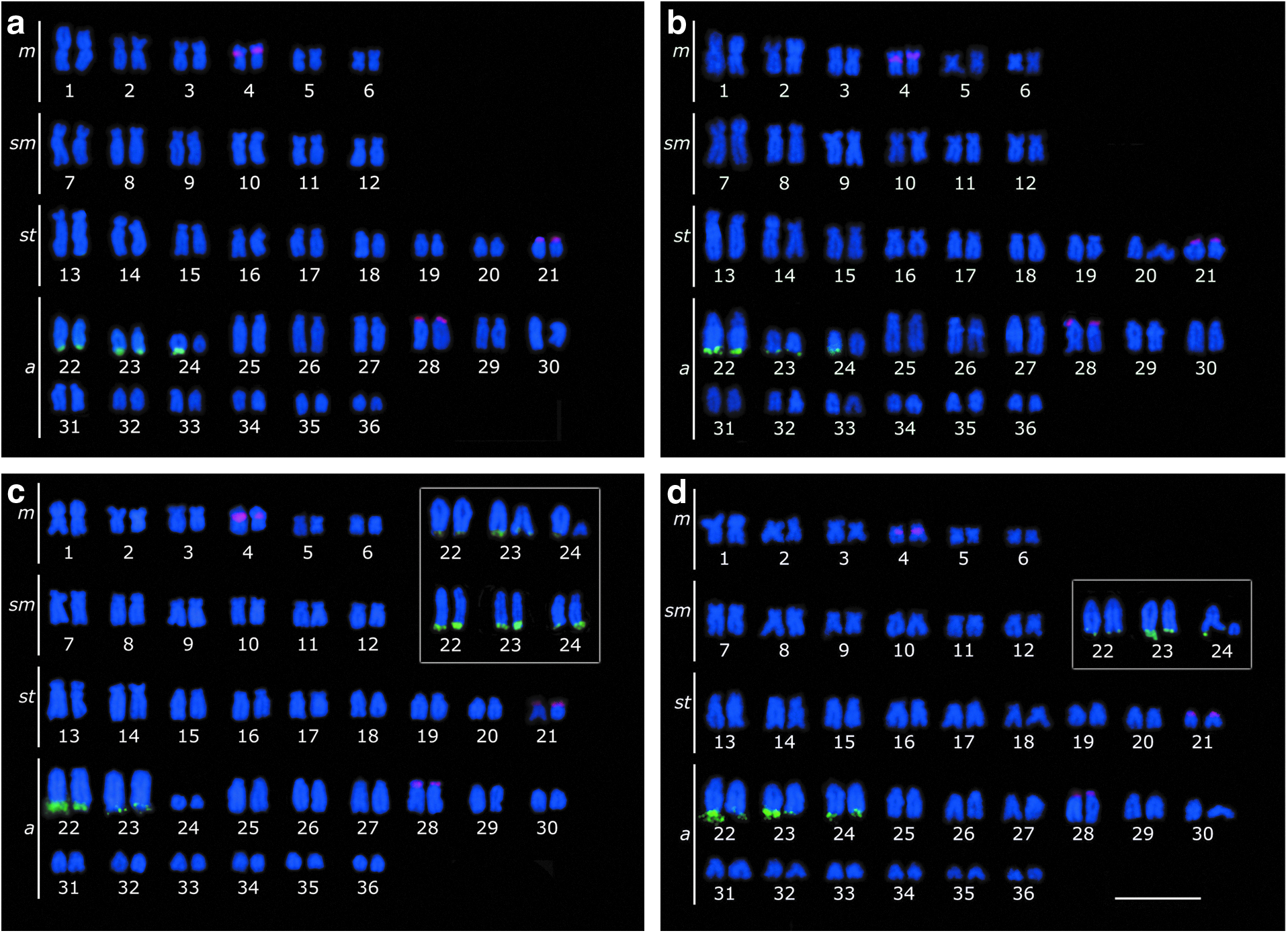
Karyotypes of H. strigaticeps after in situ fluorescence hybridization with the 5S rDNA probe (rodamine, red) and 18S rDNA probe (FITC, green) of populations from

Metaphases after CMA3 (left) and DAPI (right) treatment of populations of H. strigaticeps from
The heterochromatin revealed by C-banding was evident for all populations in the centromeric and pericentromeric regions of most chromosomes, especially small interstitial blocks on the short arm of the first pair of metacentric chromosomes, and terminal in other chromosomes (Fig. 2).
Large heterochromatic blocks CMA3−/DAPI+ were found in the proximal to terminal position of the long arm of acrocentric pair 22 for populations from the Atlântico Stream (Figs. 2a and 4a, b) and the Piquiri River (Figs. 2b and 4c, d). Additionally, the population from the Piquiri River had interstitial heterochromatic blocks, CMA3−/DAPI+, on pairs of acrocentric chromosomes 26, 29, 31, and 35 (Figs. 2b and 4c, d). The populations from the Iguassu River and Paraná River showed similar results, in the form of large CMA3−/DAPI+ heterochromatin blocks on acrocentric chromosomes on pairs 22, 23, and 24 adjacent to NORs (Figs. 2c, d and 4–h).
All populations had acrocentric chromosomes bearing the NORs; thus, the pairs were arranged in the same position within the karyotype (pairs 22, 23, and 24) to facilitate discussions about the evolution of these segments as well as the adjacent sequences.
Additionally, populations of the Paraná and Iguassu rivers showed interpopulational polymorphism of the CMA3−/DAPI+ heterochromatin blocks on pair 24, revealing three situations: (1) homozygous (HH) with large heterochromatic blocks (proximal to terminal), (2) heterozygous (Hh) with an extensive heterochromatin block (proximal to terminal) on only one of the homologous chromosomes and a small terminal heterochromatic block on the other, and (3) homozygous (hh) with the small terminal heterochromatic in both chromosomes (Fig. 2c, d).
Morphological results of the four populations of H. strigaticeps by canonical analysis allowed identifying three groups in the graph of CVA: (1) Atlântico Stream, (2) Piquiri River, and (3) Paraná and Iguassu Rivers gathered together (Fig. 5).
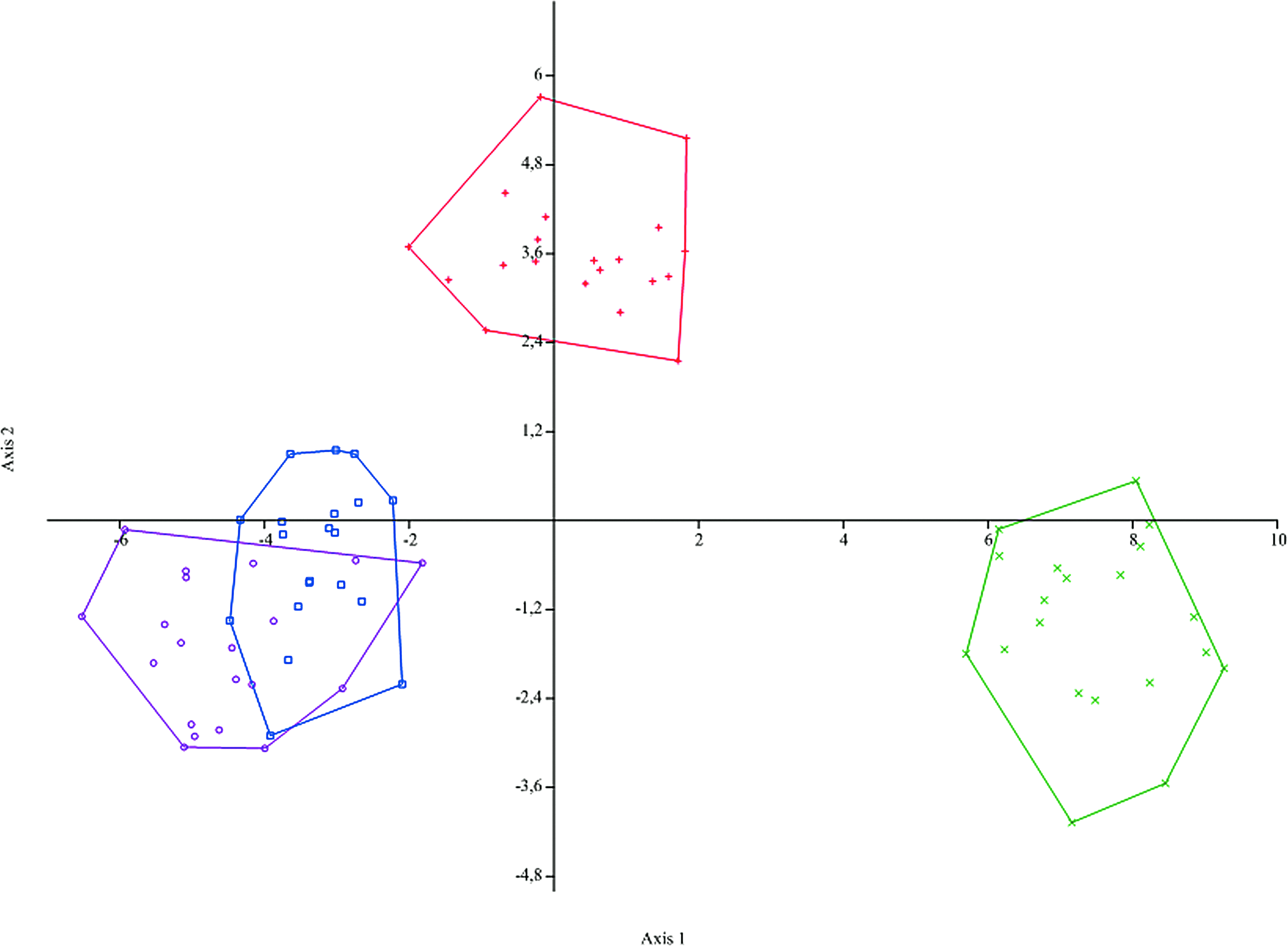
Projection of canonical variables analyses (CV) of the size-independent morphometric data of populations of H. strigaticeps from (x, green) Atlântico Stream, (+, red) Piquiri River, (□, blue) Paraná River, and (○, purple) Iguassu River. Color images available online at www.liebertpub.com/zeb
Discussion
The karyotypic macrostructure of H. strigaticeps is relatively conserved since all four populations analyzed showed 72 chromosomes, conservative karyotype formulae, and similarity in number (at least two pairs) and position of NORs. Multiple NORs are considered a derived characteristic and is most often found in species of Hypostomus from the southern basins of South America. 13 However, differences could be observed in the number of AgNOR-bearing chromosomes in the populations of the present study (2 to 3 pairs), as well as a diversity in the size of the acrocentric chromosomes carrying these sequences, and also some divergence from populations from the Paranapanema River basin39,40 that showed two pairs of acrocentric NOR-bearing chromosomes. Nevertheless, all populations of H. strigaticeps share NORs on the terminal position of the long arm of acrocentric chromosomes. The numerical polymorphism might be related to dispersion of rDNA sites that, in the populations of H. strigaticeps in this study, are associated with large polymorphic heterochromatin blocks.
The four populations of H. strigaticeps presented a conserved number and location (interstitial and terminal) pattern of 5S rDNA sequences. These genes, located at the interstitial position of a metacentric chromosome pair, similar to pair 4 in the present study, have been reported for H. strigaticeps, 40 Hypostomus regani, 41 Hypostomus nigromaculatus 42 and Hypostomus iheringii. 21 Although only few studies about 5S rDNA sequences in Hypostomus have been carried out so far, frequent interstitial marking on a pair of metacentric chromosomes suggests its ancestry. Thus, this interstitial position of 5S rDNA has been reported in many species of fish and might also represent a more stable region less prone to dispersion.43,44
Multiple 5S rDNA cistrons besides the interstitial marking have been observed in other species of Hypostomus such as Hypostomus affinis, 44 H. regani, 41 H. nigromaculatus and H. strigaticeps from the Hortelã Stream. 40 In the populations of H. strigaticeps, in the present study, the additional 5S rDNA cistrons identified on two pairs of chromosomes (subtelo/acrocentric) in the terminal position on the short arm confirm the diversity in the distribution of these sequences in Hypostomus. An alternative and parsimonious hypothesis is that the increased dispersion of copies of the 5S rDNA genes from interstitial sites (primitive condition) might be associated with different types of Robertsonian and non-Robertsonian rearrangements that occurred during the karyotype evolution of this genus. 44 Centric fission is likely to be an important mechanism that contributed to the 5S rDNA reorganization. Considering 2n=54 chromosomes as basal for Loricariidae, 10 these kind of rearrangements would result in an increase in the diploid number, as verified for Hypostomus species (2n=64, 66, 70, 72, 74, 76, 77, 80, 82, and 84) and therefore an increase in the number of 5S rDNA sites as well as in the complexity of the genome organization. Another hypothesis that could explain the increased dispersion of copies of the 5S rDNA genes would be the use of a spreading mechanism of retrotransposons by the ribosomal DNA, which subsequently affects recombination rates in genomes and leads to a rapid genome/karyotype divergence, as observed for Coregonus albula and Coregonus fontanae genomes. 45
Regarding the distribution of heterochromatin, the four populations of H. strigaticeps shared heterochromatic blocks at the centromeric, pericentromeric, and interstitial positions on pairs 1 and 4. Nevertheless, particularities regarding the location and amount of heterochromatin in different populations allow them to be characterized by this marker. The interstitial heterochromatic blocks on acrocentric chromosomes (pairs 26, 29, 31, and 35) verified for the population from the Piquiri River, the large heterochromatic blocks on pair 22 (acrocentric) in four populations, and additional heterochromatic blocks on acrocentric chromosomes (pairs 23 and 24) in the populations from the Paraná and Iguassu rivers are a clear interpopulational polymorphism, which resulted in differences in the microstructure of the karyotype of these populations of H. strigaticeps.
H. strigaticeps from the Atlântico Stream was correlated and shown to be similar to the population type from the Piracicaba River by a molecular study. 24 The large heterochromatic block present in pair 22 is also shared by other populations; however, there is an increase in the heterochromatin amount in populations from the Paraná and Iguassu rivers at an equivalent position on pairs 23 and 24. Furthermore, the size variations between homologues of pair 24, evidenced by Giemsa staining and C-banding, also characterize interpopulation chromosomal polymorphism. Mechanisms of heterochromatin amplification could explain the observed polymorphism (Fig. 6), probably either as a result of gene duplication generated by unequal crossing over or transposition of sequences that have become heterochromatic, or else by unequal exchanges generated by proximity and arrangement at interphase nuclei that, according to the model for heterochromatin dispersion, might facilitate this type of reorganization between homologous segments. 46
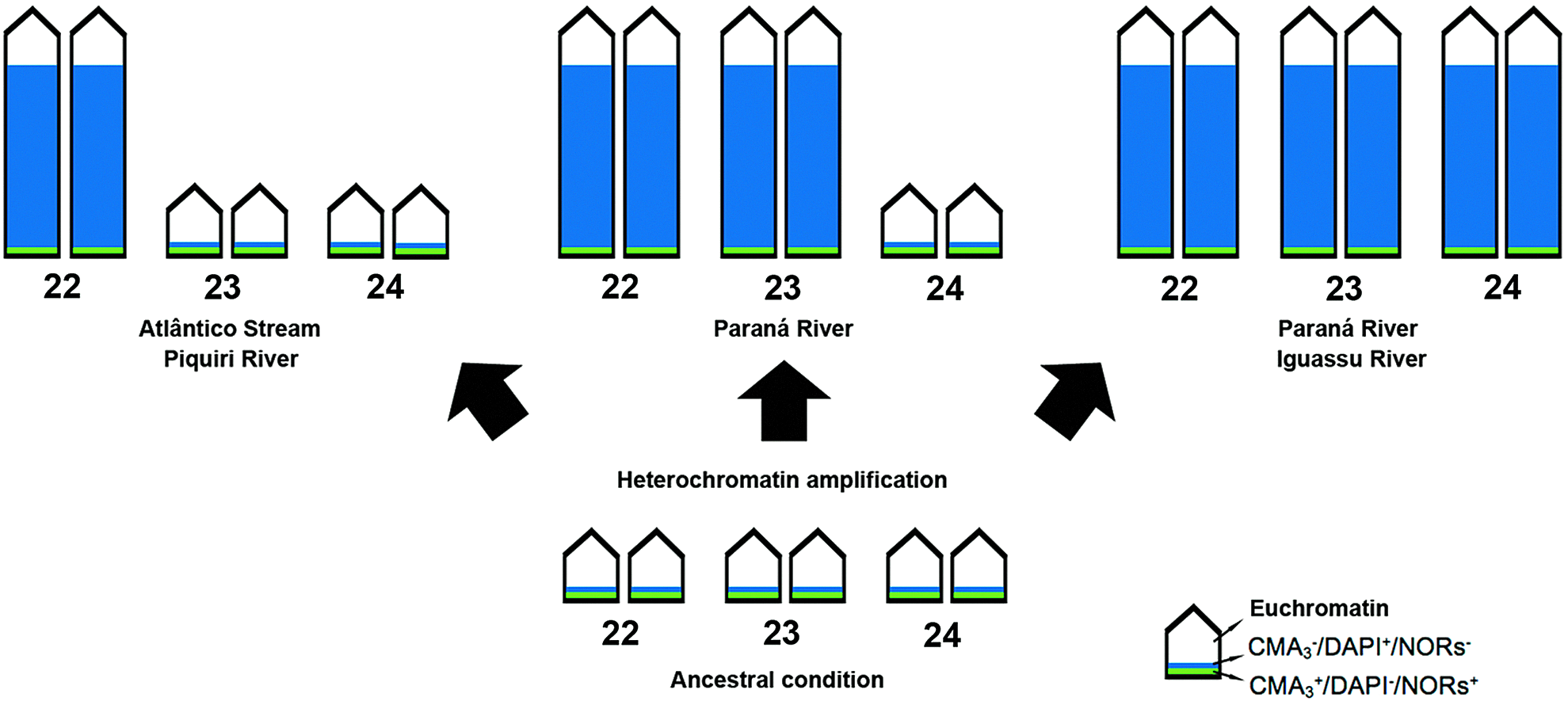
Representative idiogram of the heterochromatin amplification process in H. strigaticeps from hypothetic ancestral condition. AgNORs (green), heterochromatin (blue), and euchromatin (white). Color images available online at www.liebertpub.com/zeb
The heterochromatin located at the interstitial position on four pairs of acrocentric chromosomes (pairs 26, 29, 31, and 35) in the population from the Piquiri River was not observed for other populations of H. strigaticeps. On the other hand, interstitial blocks have been reported in karyotypes of Hypostomus sp. E, Hypostomus sp. F, 10 and H. regani. 41 Interstitial heterochromatic blocks on large acrocentric chromosomes were verified in Hypostomus sp. E and Hypostomus sp. F and suggested a transposition from terminal segments of small chromosomes to equilocal interstitial sites between nonhomologous chromosomes that are close to each other in interphase nuclei, 10 as predicted in the heterochromatin dispersion model. 46 This model could explain the origin of the interstitial heterochromatin blocks in the populations from the Piquiri River; however, further analyses involving satellite DNA markers in species with this type of distribution of heterochromatin are required to provide evidence for these mechanisms.
Chromosomal differentiation among populations, through the distribution and amount of heterocromatin, showed that these repetitive sequences are an important populational marker. It seems that the increase in number of these segments is directly proportional to the path of the Paraná River, that is, populations collected downstream in the Paraná River basin (Paraná and Iguassu River populations) have more heterochromatin compared with upstream populations (Atlântico Stream and Piquiri River populations). It is likely that this heterochromatin may have occurred in response to selective environmental pressures. Subpopulations based on heterochromatin distribution were well demonstrated in L. elongatus from the Paraná River basin. 47 According to the authors, the construction of several hydroelectric dams in the last 50 years between the Mogi-Guaçu and Paranapanema Rivers blocked their physical continuity, inducing the formation of subpopulations that are currently differentiated by their heterochromatin distribution. However, the authors do not rule out the possibility that geographical barriers played this role before the construction of these dams.
The quantity and distribution of heterochromatin seem to be correlated to the biogeographical characteristics of H. strigaticeps. Thus, the Atlântico Stream (present study) and Paranapanema River populations 39 are differentiated from the others by its low level of heterochromatin, a characteristic that is probably a result of its isolation. Therefore, interstitial heterochromatin blocks present in the population from the Piquiri River are also likely the result of its geographic isolation. The Piquiri River is characterized by the presence of rapids, springs, and straits and its ichthyofauna differs considerably from other tributaries of the Paraná River. 48 On the other hand, the similarity in the cytogenetic data between populations from the Paraná River (Upper Parana River basin) and the Iguassu River (Middle Paraná River basin) is probably due to the connection between these regions. This natural contact occurred before the construction of the Itaipu Hydroelectric Dam in the 80s, which limited contact between these two populations to the Spawning Channel, a deviation created by ITAIPU to provide the fishes with contact between the Paraná River (downstream of the dam) and the reservoir.
Not only cytogenetic data, but also morphometric results showed the diversity of the populations of the present article, including the isolation of the populations from the Atlântico Stream, Piquiri River, and the two overlapped populations from the Paraná and Iguassu Rivers in the CVA graph (Fig. 5). Thus, the environmental diversity of each population, aforementioned, probably also reflects in the morphologic pattern of these populations. Morphological diversity was also demonstrated in seven Characiformes fish species, concerning the development of dermal lip protuberances; it seems like these structures are associated with environmental hypoxia. 49 The ability of an organism to express different phenotypes depending on the biotic and abiotic environment is denominated phenotypic plasticity, considered the acclimatization of the organisms to their environment to improve performance. 50 Moreover, there is often a lack of sufficient genetic variation in plasticity to allow evolution. 51 Consequently, the morphologic diversity verified in the populations of H. strigaticeps is likely to be related to the phenotypic plasticity of the species, which is suggested by the isolation of those three groups in CVA results.
Despite differences pointed out by the cytogenetic and morphometric data, all the populations analyzed in this study belong to H. strigaticeps, corroborating the hypothesis of dispersal of this species through the Paraná River basin, as proposed by molecular data. 24 However, it is clear that there is interpopulational variation, mainly related to heterochromatin distribution. Due to the high cytogenetic variability observed in Hypostomus, the diversity of these segments shows that the evolutionary processes acted on the differentiation of these populations. The heterochromatic polymorphism evidenced suggests that it might have an important role in the evolutionary history of Hypostomus, as has been already reported to be the case for other species of this genus. Furthermore, in spite of the phenotypic plasticity verified by the morphometric results, each population seems to be on its own path of evolution.
Footnotes
Acknowledgments
The authors are grateful to the Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio) for authorizing the capture of the fishes. The authors are thankful to the Universidade Estadual de Maringá, Universidade Estadual do Oeste do Paraná, Núcleo de Pesquisas em Limnologia, Ictiologia e Aquicultura (Nupélia), Parque Nacional do Iguassu, and the Macuco Safari for logistical support. This study was financed by the Fundação Araucária (Fundação Araucária de Apoio ao Desenvolvimento Científico e Tecnológico do Estado do Paraná), CAPES (Coordenadoria de Aperfeiçoamento de Ensino Superior), and CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico).
Disclosure Statement
No competing financial interests exist.