
Abstract
Abstract
Genetic mutations and environmental toxins are known to affect mitochondrial health and have been implicated in the progressive degeneration of dopaminergic neurons in Parkinson's disease. To visualize mitochondria in dopaminergic neurons of live zebrafish, we used the regulatory elements of the dopamine transporter (dat) gene to target a reporter, mCherry, after fusion with the mitochondrial localizing signal (MLS) of Tom20. Immunoblot analysis of mitochondrial and cytosolic fractions from Tg(dat:tom20 MLS-mCherry) larvae shows that mCherry is efficiently targeted to the mitochondria. Confocal imaging of live fish was carried out from 1 day postfertilization (dpf) to 9 dpf. We also colocalized dat mRNA expression with the mCherry protein in the olfactory bulb (OB), subpallium (SP), pretectum (Pr), diencephalic clusters 2 and 3 (DC2/3), caudal hypothalamus (Hc), locus coeruleus (LC), anterior preoptic area (POa), retinal amacrine cells (RAC), caudal hypothalamus (Hc), and preoptic area (PO). Treating Tg(dat:tom20 MLS-mCherry) larvae with the dopaminergic neurotoxin MPTP (1-Methyl-4-phenyl-1,2,3,6-tetrahydropyridine) at 2 or 3 dpf resulted in a decrease in mCherry fluorescence in the pretectum, olfactory bulb, subpallium, diencephalic clusters 2 and 3, and the caudal hypothalamus. Labeling of mitochondria in nigrostriatal dopaminergic neurons of zebrafish could allow their visualization in vivo following genetic or pharmacological manipulations.
Introduction
M
Methods of visualizing mitochondria in living animal models, especially in cells relevant to PD, are crucial to the development of an in vivo model, which will allow us to test paradigms of DA neuron death or neuroprotection. The targeting of fluorescent markers to the mitochondria of transgenic zebrafish has facilitated the study of mitochondrial function under specific physiological and/or genetic conditions.4–6 However, these transgenic models labeled the whole fish or the entire nervous system, but did not target specific neuronal population such as DA neurons.
In this study, we have developed a zebrafish model, which specifically labels the mitochondria of DA neurons by introducing a fusion protein that consists of the mitochondrial localizing signal (MLS) of Tom20 and the mCherry fluorescent protein. This fusion protein was placed under the control of the cis-regulatory elements of the dopamine transporter (dat) gene. This transgenic line allows the observation of mitochondria in real time, specifically in DA neurons, to better our understanding of the pathogenesis of PD following chemical neurotoxin exposure.
Materials and Methods
Animal maintenance
All experiments were performed according to the guidelines of the Canadian Council on Animal Care and were approved by the University of Ottawa Animal Care Committee. Zebrafish and embryos were maintained at 28.5°C according to methods described by Westerfield. 7 Wild-type adult zebrafish were kept and bred in water at 28.5°C with a controlled 14-h light cycle. Embryos were collected at the one-cell stage and were raised at similar densities at 28.5°C. Embryos were killed with an overdose of tricaine mesylate (ethyl 3-aminobenzoate methanesulfonate; Sigma-Aldrich, Oakville, Canada).
Transgenic construct and zebrafish transgenesis
To prepare the targeting construct for homologous recombination in bacteria (Liu et al., 2003), we amplified through polymerase chain reaction (PCR) and subcloned the zebrafish Tom20 MLS-mCherry sequence (MLS sequence: ctcgtcatgatgggcggcagcagcagcaggatcgcggctggtttgggcgcggcgctcttcgtcggttactgcatttatttcgacaggaagagacggagtgacccgaac) from a construct generously gifted to us by Dr. Philippe Vernier (CNRS, Gif-sur-Yvette, France). We added a floxed Neomycin resistance (NeoR) cassette downstream of the mCherry coding sequence. We used essentially the same genomic region as in the Tg(dat:EGFP) transgenic line, 8 which is a 27-kb fragment (including ∼13 kb of the dat 5′-flanking region). The targeting construct included two arms (Arm A and Arm B, about 250 bp each), which are sequences homologous to the region of the dat locus flanking the EGFP sequence in the dat:EGFP construct. Exon 1 of dat, which includes the initiator ATG, was not included in Arm A. Thus, the translational start site of the construct corresponds to the ATG of the tom20 MLS. The pBluescript II KS+ targeting construct was digested with NotI to linearize the targeting fragment of about 3 kb. To insert this targeting fragment into the dat plasmid by homologous recombination, we electroporated DY380 electrocompetent Escherichia coli, which contain a temperature-inducible RecET homologous recombination system 9 and the dat:EGFP construct, flanked by Tol2 arms for efficient transgene integration. 8
Following homologous recombination in bacteria, we selected for positive colonies using neomycin and confirmed the presence of the construct followed by PCR using primers located at the 3′ end of the NeoR and in exon 2 of dat. The purified dat:tom20 MLS-mCherry plasmid (50 ng/μL) was coinjected with transposase mRNA (35 ng/μL), when injected into embryos at the one-cell stage using a Narishige IM300 microinjector. Injected embryos were examined for the presence of mCherry in DA neurons. Primary injected fish were raised to adulthood and screened for germline transmission following outcrossing with wild-type fish.
Western blotting and mitochondria isolation
Mitochondrial and cytosolic fractions were prepared from 5 days postfertilization (dpf) zebrafish larvae using the Mitochondria Isolation Kit for Tissue (Thermo Scientific, Rockford, IL). Protein concentration was determined by bicinchoninic acid (BCA) assay, and 20 μg of protein was loaded into each lane of a 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis gel. The following antibodies were used for western blotting: rabbit anti-dsRed (1:1000) (Clontech, Mountain View, CA); mouse anti-GAPDH (1:2000) (Abcam, Boston, MA); rabbit anti-VDAC1/Porin (1:1000) (Abcam); donkey anti-rabbit-IgG-HRP antibody (1:5000) (GE Healthcare, Little Chalfont, United Kingdom); and goat anti-mouse-IgG-HRP antibody (1:10,000) (Jackson Immuno Research Laboratories, West Grove, PA).
Imaging
For two-photon confocal imaging, live embryos and larvae were embedded in 1% low melting point agarose and imaged using a Nikon A1 MP Multiphoton Confocal Microscope equipped with a 25× water dipping objective. Maximum intensity projection images are composed of z-stacks of ∼200 μm total thickness. For confocal microscopy of mitochondria, a 60× oil immersion objective was used to image 14 μm-thick sections following immunostaining. All images are dorsal facing up and anterior facing left, unless stated otherwise.
Combined fluorescent whole-mount in situ hybridization and immunostaining
For in situ and immunostaining experiments, 3 dpf larvae were fixed in 4% paraformaldehyde, washed in phosphate buffered saline with tween (PBST), dehydrated in methanol, and stored at −20°C. A dat cRNA probe was synthesized as described previously 8 and labeled with digoxigenin and developed using Tyr-Cy3 (Perkin-Elmer, Woodbridge, Canada). The following antibodies were used to detect mCherry: rabbit anti-dsRed (Clontech) and anti-rabbit AlexaFluor® 488 (Molecular Probes, Burlington, Canada). For in situ and immunostaining experiments, larvae were mounted on slides and images were acquired with a Zeiss LSM 510 upright confocal microscope equipped with a 20× air objective.
MPTP treatment
Fresh MPTP (1-Methyl-4-phenyl-1,2,3,6-tetrahydropyridine; Sigma-Aldrich) was dissolved in autoclaved Milli-Q water to prepare a 10 mM stock solution and used at a working concentration of 1 mM in 1× embryo media. 7 mCherry-positive embryos and larvae were exposed to MPTP, starting at either 48 or 72 hour postfertilization (hpf) with the MPTP-containing media changed daily. Control fish received 1× embryo media only. Live fish were imaged by confocal microscopy as described previously.
Statistical analyses of fluorescence quantification
Z-stacks comprising individual DA neuron regions were compiled, and fluorescence pixel intensity was measured using ImageJ. DA neuron regions from control fish were compared to those from fish treated with 1 mM MPTP. Differences in mean fluorescent intensity were analyzed by two-tailed unpaired Student's t-tests with three biological replicates. Predetermined p-values ≤ 0.05 were considered statistically significant (p < 0.05, p < 0.01, p < 0.001). All analyses were performed using Excel (Microsoft, Redmond, WA).
Results
Transgenic construct design for the establishment of a transgenic zebrafish line
Based on a construct we previously used to generate a Tg(dat:EGFP) zebrafish transgenic line expressing EGFP under the control of the cis-regulatory elements of the dat gene, 8 we designed a new construct (Fig. 1A) to express a fluorescent fusion protein targeted to the outer mitochondrial membrane by fusing mCherry with the MLS of zebrafish Tom20, a receptor located in the outer mitochondrial membrane. 10
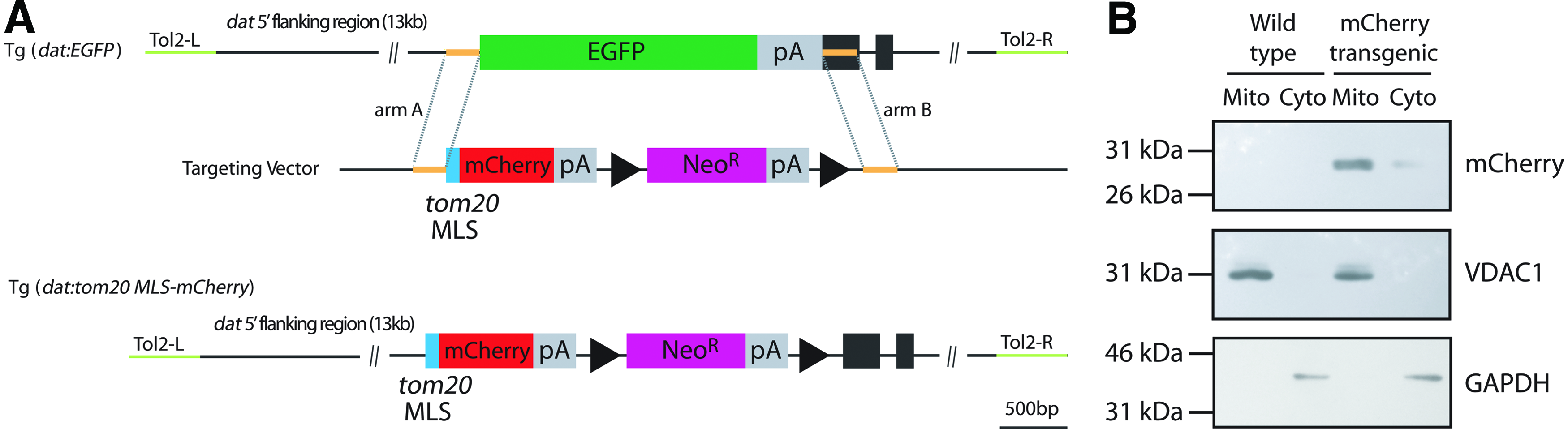
Construct to produce Tg(dat:tom20 MLS-mCherry) zebrafish line.
Primary transgenic fish were raised to adulthood (∼4 months) and screened for germ line transmission of the transgene. We obtained seven independent transgenic lines. Of these, we selected a line that expressed strong mCherry fluorescence in several DA neuron clusters (Fig. 2A, B), including the subpallial region that is considered to be involved in movement control in zebrafish. 11
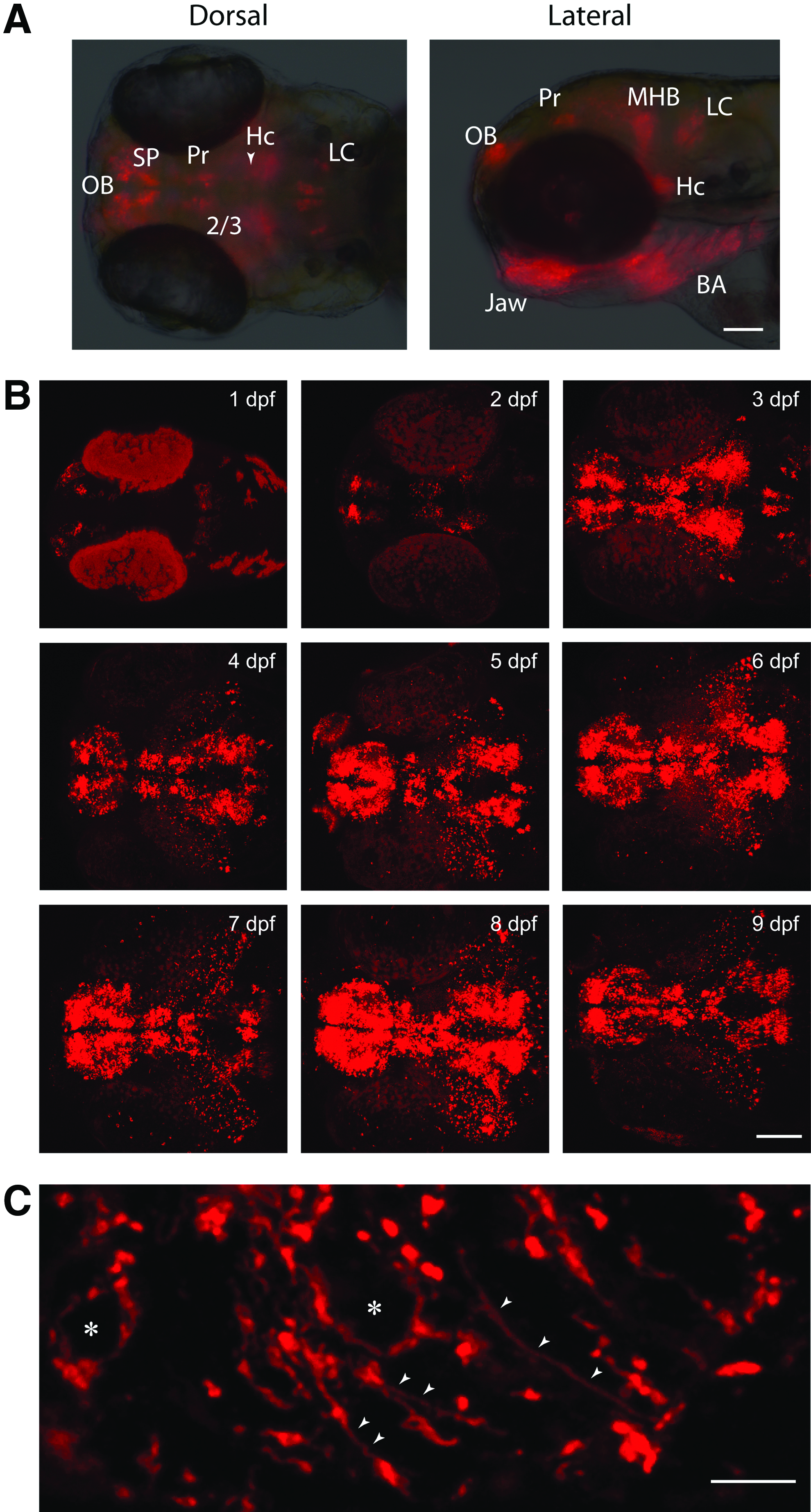
Expression of mCherry in the Tg(dat:tom20 MLS-mCherry) fish.
mCherry is properly targeted to the mitochondria in transgenic zebrafish
We isolated mitochondria from transgenic larvae at 5 dpf and compared the mCherry protein detected in the mitochondrial fraction to that in the cytosolic fraction by western blotting (Fig. 1B). A band representing mCherry is detected in the mitochondrial fraction from transgenic fish, while a low amount of mCherry is detected in the cytosolic fraction of transgenic fish, likely due to the transient transport of the Tom20 MLS-mCherry fusion protein to the mitochondria.
Colocalization of mCherry patterning with a dopaminergic neuron marker in transgenic zebrafish larvae
We examined the expression of mCherry in the transgenic Tg(dat:tom20 MLS-mCherry) zebrafish line. Fluorescence in this line was visible within the first 24 hpf, which agrees with prior observations of DA neuron development. 11 The Tg(dat:tom20 MLS-mCherry) transgenic line expressed mCherry in the olfactory bulb (OB), subpallium (SP), pretectum (Pr), diencephalic clusters 2 and 3 (DC2 and DC3), caudal hypothalamus (Hc), jaw, branchial arches (BA) (Fig. 2), retinal amacrine cells (RAC) (Fig. 3E″), and reticular astrocytes 12 of the optic nerve (data not shown). These cells of the jaw, branchial arches (BA), and reticular astrocytes of the optic nerve were seen in the Tg(dat:EGFP) line, 8 as well as a third transgenic line using the same genomic sequences from the dat locus. 13 Global dorsal and lateral views of the overall mCherry expression in live larvae were taken at 3 dpf (Fig. 2A). In addition, two-photon confocal images were also taken from 2 to 9 dpf (Fig. 2B). Examining immunostained sections of larvae imaged under higher magnification using confocal microscopy revealed mCherry immunolabeling along axonal projections (Fig. 2C, arrowheads) and in the cytoplasm (Fig. 2C) of olfactory bulb DA neurons.
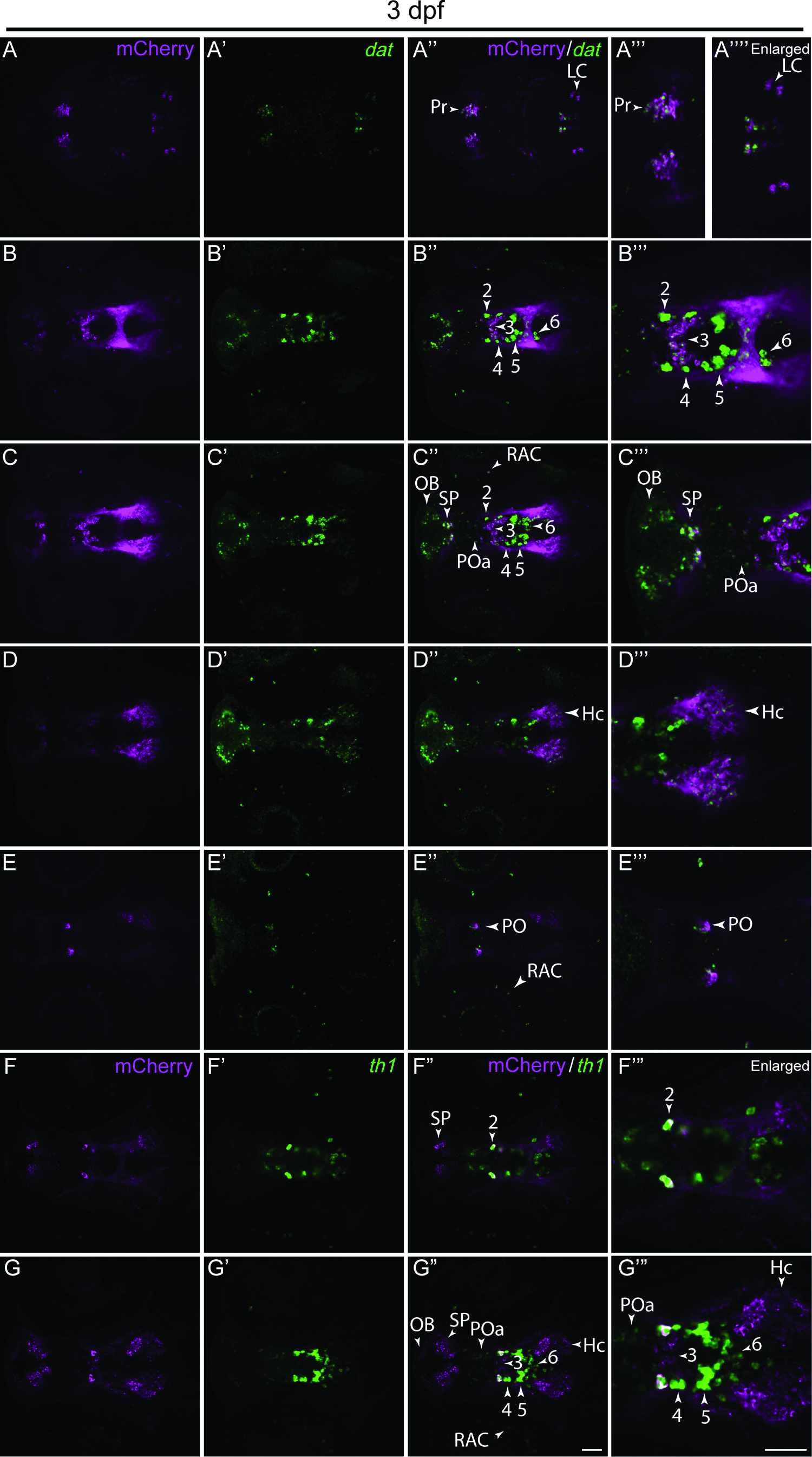
Confocal z-stacks of immunolabeled mCherry colocalization with dopamine transporter (dat) (
We colocalized the mCherry reporter by whole-mount immunostaining with the dat by whole-mount in situ hybridization (Fig. 3A–E′″) and used a th1 cRNA probe as a landmark for the identification of cells in the ventral diencephalon (Fig. 3F–G′″ and data not shown). The analysis of individual z-stacks revealed colocalization between mCherry and dat-positive cells in the pretectum (Pr), locus coeruleus (LC), diencephalic clusters 2 and 3 (DC2 and DC3), olfactory bulb (OB), subpallium (SP), anterior preoptic area (POa), retinal amacrine cells (RAC), caudal hypothalamus (Hc), and preoptic area (PO) (Fig. 3A–E′″). Using mCherry immunostaining and th1 in situ hybridization, we observed colocalization within the Pr, SP, OB, POa, DC2, DC3, RAC, and PO, but not within DC4, DC5, DC6, and Hc (Fig. 3F–G′″ and data not shown). These latter regions that have mCherry expression, but lack th1 expression, could be explained by the more predominant th2 expression in these regions. 14 Moreover, using live imaging, we observed some mCherry expression in cells, which are unlikely to be dopaminergic. For instance, we saw colocalization between dat-driven mCherry and EGFP driven by regulatory elements of the dlx5a/6a locus, even though there is no evidence that dlx genes are expressed in dopaminergic neurons. The fate of these cells is currently under investigation.
MPTP neurotoxin treatment reduces mCherry expression in Tg(dat:tom20 MLS-mCherry) larvae
We exposed embryos of the Tg(dat:tom20 MLS-mCherry) line to MPTP. Starting at 2 dpf, ∼30–50 embryos were treated with 1 mM MPTP for 72 h (Fig. 4A). A visible reduction of mCherry fluorescence in live MPTP-treated fish compared to untreated fish was observed at 120 hpf (Fig. 4B) and 144 hpf (Fig. 4C). We quantified the pixel intensity representing mCherry fluorescence in five separate regions containing DA neurons in both controls and MPTP-treated fish. At 120 hpf, a significant decrease in fluorescence was observed in the pretectum (29%, p < 0.05), olfactory bulb (33%, p < 0.05), subpallium (34%, p < 0.05), DA neuron clusters 2 and 3 (40%, p < 0.05), and caudal hypothalamus (Hc) (44%, p < 0.05). Moreover, the observed decrease in mCherry signal was more pronounced at 144 hpf in the pretectum (32%, p < 0.05), olfactory bulb (41%, p < 0.01), subpallium (55%, p < 0.01), DA neuron clusters 2 and 3 (57%, p < 0.05), and caudal hypothalamus (Hc) (62%, p < 0.01). Similar effects were observed when MPTP treatment was started at 3 dpf and continued for 48 h (data not shown). Overall, these results suggest that these DA neuron regions in the Tg(dat:tom20 MLS-mCherry) line are susceptible to the neurotoxin, MPTP, which targets mitochondria in DA neurons.
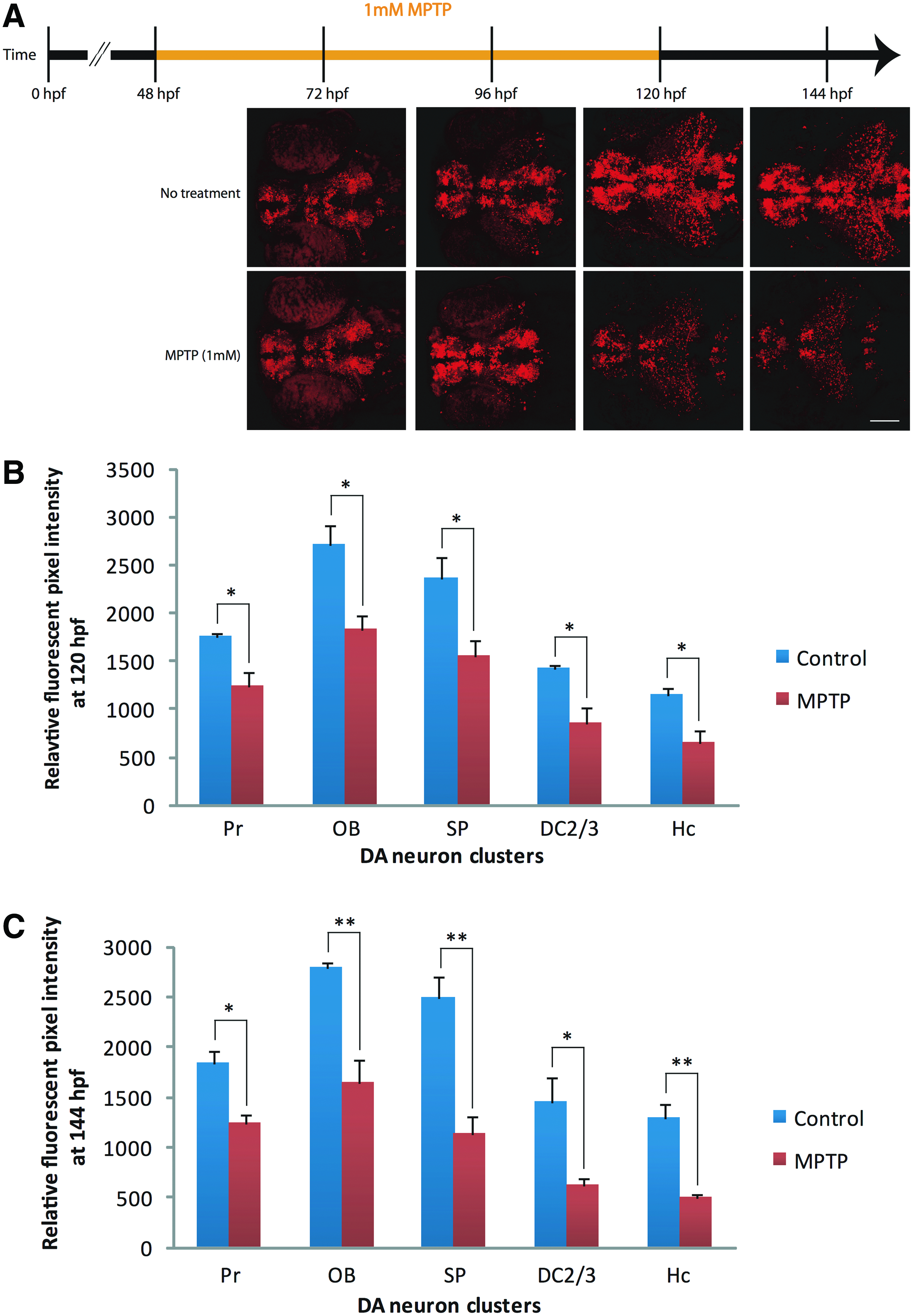
MPTP (1-Methyl-4-phenyl-1,2,3,6-tetrahydropyridine) neurotoxin exposure in transgenic larvae.
Discussion
We generated a transgenic zebrafish model for the study of mitochondria, specifically in DA neurons. The subpallial (SP) DA neurons, which are considered to be involved in movement control in zebrafish, 11 along with DA neurons present in the pretectum (Pr), locus coeruleus (LC), diencephalic clusters 2 and 3 (DC2 and DC3), olfactory bulb (OB), anterior preoptic area (POa), retinal amacrine cells (RAC), caudal hypothalamus (Hc), and preoptic area (PO), are labeled in our Tg(dat:tom20 MLS-mCherry) line. The pattern observed in the Tg(dat:tom20 MLS-mCherry) fish closely resembled the pattern of EGFP expression seen in the Tg(dat:EGFP) line. In comparison to the Tg(dat:EGFP) transgenic line, 8 the mitochondria line labels all of the same DA neuron clusters with the exception of diencephalic clusters 4, 5, and 6. The reason for this difference is unclear, but could be attributable to subtle differences in the expression pattern caused by the site of transgene integration. Despite the lack of mCherry expression in diencephalic clusters 4, 5, and 6, the mCherry expression localized to the mitochondria of subpallial DA neurons is of particular relevance to the study of motor behavior in zebrafish. The use of this transgenic model could facilitate the study of how mitochondrial dynamics correlate with behavioral analysis following DA neuron perturbations.
Data from zebrafish MPTP experiments have shown that MPTP, at doses such as 0.1 mM, affects DA neurons, but may also have an effect on other neuron populations such as gamma aminobutyric acid (GABA) neurons 15 and 5-HT cells, as previously summarized by Panula et al. 16 In this study, we opted to use a higher dose of MPTP (1 mM) to facilitate live imaging. A trend toward a reduction in mCherry (by live confocal two-photon imaging) was observed in DA neurons of the Tg(dat:tom20 MLS-mCherry) fish at lower MPTP doses (i.e., 0.1, 0.042 mM), which fall into the ranges of MPTP concentrations previously used by others.17,18 However, this trend did not reach significance and was not as easily detected by live imaging.
Previous work has shown that mitochondria in the somites and pharyngeal arch are visible through confocal microscopy in transgenic zebrafish embryos expressing EGFP targeted to the mitochondria under the control of a ubiquitous promoter, which demonstrates that live imaging of mitochondrial fusion and fission events is possible in vivo. 4 In addition, the transport and trafficking of mitochondria in zebrafish Rohon-Beard sensory nerve cells are visible in distal axon branches at the thin rim of the fin fold where the cell membrane is labeled with yellow fluorescent protein and the mitochondria are labeled with Cyan fluorescent protein. 5 Another strategy, which has been used to visualize mitochondria in live zebrafish embryos, is the transient labeling of mitochondria by injection of a construct coding for a fusion protein between the MLS of the zebrafish cytochrome c oxidase subunit VIII and EGFP. 6 Having the Tg(dat:tom20 MLS-mCherry) zebrafish model with fluorescent mitochondria in DA neurons allows the direct examination of mitochondria exclusively in DA neurons.
Using Tg(dat:tom20 MLS-mCherry) embryos and larvae, we observed a significant reduction in mCherry fluorescence in the olfactory bulb, pretectum, subpallium, diencephalic clusters 2 and 3, and caudal hypothalamus following MPTP exposure. Once the active form of MPTP, MPP+ enters a DA neuron, MPP+ targets mitochondria and inhibits complex I of the electron transport chain leading to cellular energy depletion and reduction in DA cell markers. As a consequence, MPTP is used in research to model dopamine deficiency, since it causes a 30%–40% decrease in complex I activity in the substantia nigra pars compacta. 19 McKinley et al. have previously shown that the mechanism of DA neuron toxicity induced by MPTP is conserved in zebrafish. 18 Our findings are in agreement with other MPTP studies in zebrafish, which show selectively perturbed DA neurons and result in behavioral deficits that mimic PD.8,17,18,20–23
This transgenic line is an asset for future studies aiming to screen compounds or genes thought to be involved in mitochondrial dysfunction leading to DA neuron loss. Mitochondria could be compared from toxin-treated and untreated zebrafish or genetically impaired zebrafish, such as those induced by gene knockdown (e.g., RNAi, morpholinos) and targeted genomic deletions (e.g., ZFN, CRISPRs, TALENS). The ability to follow changes in vivo will allow easier tracking of differences in real time, over several time points, using the same fish. Alternative methods would involve immunostaining or in situ hybridization, which is time-consuming and requires more biological samples. A number of endpoints could be examined, such as mitochondrial fission and fusion (e.g., counts of elongated, intermediate, and fragmented mitochondria) and mitochondrial transport along the axon. In addition, cells from the Tg(dat:tom20 MLS-mCherry) zebrafish could be dissociated and mCherry-positive DA neurons could be collected by fluorescent activated cell sorting. Mitochondria from these mCherry-positive neurons could then be isolated and used to measure mitochondrial respiration.
We described the patterning of DA neurons in the Tg(dat:tom20 MLS-mCherry) line and observed a PD-relevant phenotype following exposure of these fish to MPTP. The amenability of zebrafish to transgenesis and drug exposure facilitated our observation of mitochondria within specific DA cell populations and allowed us to examine changes following neurotoxin exposure. Since dysfunctional mitochondria may contribute to DA neuron death and mitochondria are known to possess a dynamic morphology that is regulated by mitochondrial-shaping proteins, 24 our transgenic line can be used in future studies in the context of neurodegeneration and mitochondrial dynamics (fusion and fission), specifically in DA neurons in vivo.
Footnotes
Acknowledgments
We thank Dr. Philippe Vernier (Institut de Neurobiologie, Alfred Fessard, Gif-sur-Yvette, France) for the gift of the Tom20 plasmid. Gary Hatch for help with designing the dat:tom20 MLS-mCherry construct. We thank Vishal Saxena for assistance with fish husbandry. This work was funded by grants from the Natural Sciences and Engineering Research Council of Canada and by the Canadian Institutes of Health Research (grant no. MOP-14460 and 93803). S.N. and R.G. were supported by the Ottawa chapter of the Parkinson Society of Canada (grant no. 2012–15), an Ontario Graduate Scholarship, the University of Ottawa Department of Biology Scholarship, a Government of Ontario–Graduate Scholarship in Science and Technology and the Parkinson's Research Consortium–Toth Family Fellowship in Parkinson's Research. Funding sources had no involvement in the preparation of the article.
Disclosure Statement
No competing financial interests exist.