
Abstract
Abstract
This study analyzed two Apareiodon species without available chromosome data: Apareiodon argenteus and Apareiodon davisi. Both species have 54 metacentric/submetacentric chromosomes, with centromeric blocks of heterochromatin. Nucleolus organizer regions were active in chromosome pair 2 in A. argenteus and pairs 4 and 9 in A. davisi. In A. argenteus, 45S and 5S ribosomal genes were located in chromosome pairs 2 and 18, respectively. Polymorphisms were observed in these ribosomal sequences in A. davisi, with variations in the number/position of sites, and colocalization of these genes in some chromosome pairs. The WAp repetitive fraction was dispersed along the chromosomes of the two species. The satellite DNA pPh2004 was identified in chromosome pairs 7, 8, 10, 11, and 18 in A. argenteus and in pair 24 in A. davisi. The present study describes the first case of chromosomal polymorphisms involving two ribosomal sequences in Parodontidae and discusses the role of repetitive DNAs in the genome and karyotype diversity of this family.
Introduction
P
The group has diploid number (2n) of 54 chromosomes and variations in karyotype formulas. Heterochromatin is preferentially located in the centromeric regions of the chromosomes but also occurs in terminal and interstitial blocks.5–10 The heteromorphic ZZ/ZW1W2 and ZZ/ZW sex chromosome systems are found in some species.11–17
The number and position of the chromosome sites of 45S and 5S ribosomal genes differ among species of Parodontidae, with synteny occurring in Parodon nasus Kner, 1859.16,18,19 The satellite DNA pPh2004, isolated from Parodon hilarii Reinhardt, 1866, 8 was detected in the karyotypes of Apareiodon affinis (Steindachner, 1879), P. hilarii, Parodon moreirai Ingenito and Buckup, 2005, P. nasus, and Parodon pongoensis (Allen, 1942) but not observed in Apareiodon hasemani Eigenmann, 1916, Apareiodon ibitiensis Campos, 1944, Apareiodon piracicabae (Eigenmann, 1907), Apareiodon vladii Pavanelli, 2006, Apareiodon vittatus Garavello, 1977, and Apareiodon sp. (rio Verde population). 16 The WAp repetitive fraction, obtained from the W chromosome of Apareiodon sp. (rio Verde), 20 is found in all of the species for which the cytogenetic data are available.16,17,20
Given the small number of species, in comparison to other neotropical fish groups, and the substantial karyotype diversity, Parodontidae are an interesting family for chromosomal evolution studies. Thus, the present study aimed at contributing to the knowledge on the karyotype evolution of this family through the chromosome analyses of both species Apareiodon argenteus Pavanelli and Britski, 2003 and Apareiodon davisi Fowler, 1941, without the available cytogenetic data.
Materials and Methods
Samples and collection sites
We analyzed 8 specimens (5 males and 3 females) of A. argenteus from the Araguaia River, in Aragarças, Goiás state (GO), Brazil (GPS: 15°53′32,70" S; 52°15′03,21" W) and 53 specimens (21 males and 32 females) of A. davisi from the Salgado River, in Icó, Ceará state (CE), Brazil (GPS: 06°24′25,80" S; 38°52′01,70" W). All specimens were collected with the authorization of the Chico Mendes Institute for Biodiversity Conservation (ICM-Bio; License No. 10538-1). Procedures were performed in compliance with the Ethics Committee on Animal Experimentation (process No. 13/2014) of the Universidade Estadual de Ponta Grossa (UEPG), Paraná state (PR), Brazil. The specimens were deposited in the Research Center in Limnology, Ichthyology and Aquaculture (NUPELIA) of the Universidade Estadual de Maringá (UEM), Paraná state, Brazil, and identified under the following vouchers: A. argenteus—NUP 16227; A. davisi—NUP 17080.
Chromosomal preparations and classical cytogenetic analyses
Cell suspensions containing mitotic chromosomes were obtained from the animals' kidney.21,22 Chromosomes were stained with 5% Giemsa solution to define the diploid numbers and karyotype formulas. Chromosome morphology was determined using the arms ratio. 23 The fundamental numbers (FN) were established considering metacentric (m), submetacentric (sm), and subtelocentric (st) chromosomes as having two arms and acrocentric (a) only one. Heterochromatic regions were identified by the C-banding procedure, 24 using a final staining of propidium iodide. 25 Nucleolus organizer regions (NORs) were identified using silver nitrate impregnation. 26 Classical chromosomal preparations were analyzed with an optical microscope.
Molecular cytogenetic analyses
Four sequences of repetitive DNAs were physically mapped: 45S ribosomal DNA (rDNA), 5S rDNA, satellite DNA pPh2004, and WAp repetitive fraction. The 45S and 5S ribosomal DNAs and the satellite DNA pPh2004 were amplified from clones.8,27,28 Probes were labeled by nick translation. Biotin-16-dUTP (Biotin Nick Translation Mix; Roche Applied Science, Mannheim, Germany) was used to label 18S rDNA and satellite DNA pPh2004, whereas the 5S rDNA probe was labeled with digoxigenin-11-dUTP (Dig Nick Translation Mix; Roche Applied Science), according to the manufacturer's instructions.
The WAp repetitive fraction was amplified and labeled with digoxigenin 11-dUTP (Roche Applied Science) by degenerate oligonucleotide primed—polymerase chain reactions from the microdissection of the heterochromatic part of W chromosome of Apareiodon sp. (rio Verde population). 20
Fluorescence in situ hybridization (FISH) 29 was performed under high stringency conditions (77%), with a hybridization mix that consisted of 200 ng of each probe, 50% formamide, 10% dextran sulfate, and 2× SSC (concentrated salt solution). The slides were hybridized for 16 h. Signal detection was carried out using avidin-fluorescein isothiocyanate (Sigma-Aldrich, St. Louis, MO) for 18S rDNA and pPh2004 probes and anti-digoxigenin-rhodamine (Roche Applied Science) for 5S rDNA and WAp probes.
Molecular chromosomal preparations were analyzed in an epifluorescence microscope with the appropriate filters. The selected images were captured using the DP2-BSW software, with a camera coupled to an Olympus BX51 microscope.
Results
Apareiodon argenteus specimens exhibited 2n = 54 metacentric/submetacentric chromosomes and FN = 108 (Fig. 1a). Active NORs were identified in the terminal region of chromosome pair 2 (Fig. 1a, box). Heterochromatic blocks were observed primarily in the centromeric and terminal regions of the chromosomes, with conspicuous blocks in pairs 2, 3, 4, 5, 15, 16, 18, and 20 (Fig. 1b).

Classical cytogenetics of Apareiodon argenteus and Apareiodon davisi. Karyotype of A. argenteus with conventional Giemsa staining
Apareiodon davisi specimens exhibited 2n = 54 metacentric/submetacentric chromosomes and FN = 108 (Fig. 1c). Active NORs were detected in the terminal region of the long arm of chromosome pairs 4 and 9 (Fig. 1c, box). Heterochromatic blocks were identified mainly in the centromeric and terminal regions of the chromosomes, with conspicuous blocks in pairs 3, 5, 11, 12, and 20 (Fig. 1d).
In A. argenteus, the 45S rDNA sequence was located only in the terminal portion of chromosome pair 2 (Fig. 2a), whereas the 5S rDNA was found in the interstitial region of the short arm of chromosome pair 18 (Fig. 2a). Satellite DNA pPh2004 was observed in the terminal portion of both chromosome arms in pairs 7, 8, 11, and 18 and in the terminal region of the long arm in pair 10 (Fig. 2b). The WAp repetitive fraction was present in several chromosome pairs, with blocks preferentially located in the terminal chromosome regions (Fig. 2b).
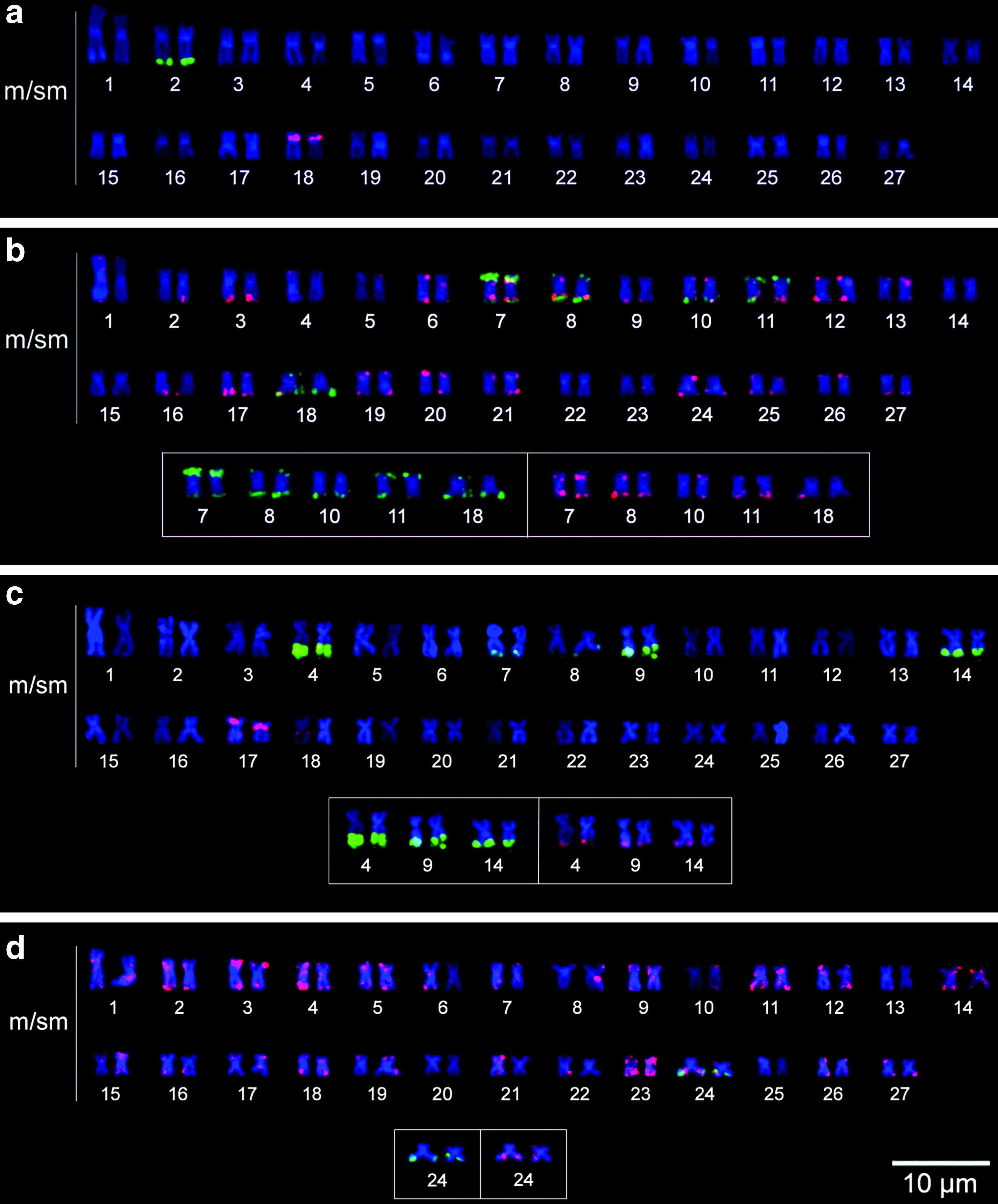
Molecular cytogenetics of Apareiodon argenteus and Apareiodon davisi. Karyotype of A. argenteus submitted to fluorescence in situ hybridization (FISH) with 18S (green signal) and 5S (red signal) ribosomal DNA (rDNA) probes
In A. davisi, FISH with 45S and 5S rDNA probes identified multiple sites of these sequences (Fig. 2c). The satellite DNA pPh2004 was found in the terminal portion of the long arm of pair 24 (Fig. 2d), and the WAp repetitive fraction was dispersed throughout the chromosomes, with blocks preferentially located in terminal regions (Fig. 2d).
Polymorphisms of the ribosomal sequences were observed in A. davisi (Fig. 3). Pairs 4, 9, and 14 contained chromosomes with colocalized blocks of 45S and 5S rDNAs in the terminal region of the long arm, as well as chromosomes without ribosomal sequences (Fig. 3). Pair 5 exhibited chromosomal variants with one 45S rDNA and two 5S rDNA sites in the terminal regions and chromosomal variants that carry no ribosomal sequences (Fig. 3). Chromosome pairs 7 and 8 exhibited variants with a terminal block of 45S rDNA in the long arm, as well as chromosomes with no rDNA sites (Fig. 3). Chromosome pair 17 contained variants with a large 5S rDNA site in the interstitial region of the short arm and colocalized blocks of 45S and 5S rDNAs in the terminal region of the long arm; a chromosomal variant with two 5S rDNA sites and chromosomal variants with the large 5S rDNA site (Fig. 3). Chromosome pair 18 showed an interstitial 5S rDNA site in the long arm and chromosomal variants without this marking (Fig. 3).

Polymorphisms of A. davisi. Chromosome pairs bearing the polymorphism of the ribosomal genes 45S (green signal) and 5S (red signal). Color images available online at www.liebertpub.com/zeb
Discussion
The specimens of A. argenteus and A. davisi showed 2n = 54 chromosomes, similar to other species of Parodontidae. Only A. affinis females have 55 chromosomes, as a result of the ZZ/ZW1W2 sex chromosomes system. 11 All the chromosomes in A. argenteus and A. davisi are metacentric/submetacentric, as observed for P. moreirai cit. Parodon sp., 13 P. moreirai, 20 and A. hasemani. 17 Almost all of the Parodontidae species have karyotype formulas with metacentric/submetacentric chromosomes 16 ; when present, subtelocentric chromosomes vary from 1 to 3 pairs. The exception is A. affinis from the lower Paraná River, for which the number of acrocentric chromosomes varies.30–33 Given the small variation in karyotype formula, the chromosomal diversity of this family is likely due to pericentric inversions and repetitive DNA accumulation and movement.16,34,35
Heterochromatin is a good chromosomal marker for the detection of ZZ/ZW heteromorphic sex chromosomes and differentiated karyotypes in Parodontidae. 16 The heterochromatin pattern identified in A. argenteus and A. davisi indicates no differentiated sex chromosomes in either species. These results corroborate those found for A. piracicabae and A. vittatus, which have no differential sex chromosomes.6,16 However, the exclusive heterochromatic blocks observed in A. argenteus corroborates the hypothesis that repeated DNA sequences play a major role in the chromosomal differentiation of Parodontidae.16,20 Overlapping active Ag-NORs sites and heterochromatic blocks were also found in the two species analyzed, as described for other species of the family, such as A. affinis,30,32 P. moreirai cit. Parodon sp., 13 A. ibitiensis, 15 and A. hasemani. 17 The single Ag-NOR detected in A. argenteus is seen in most of the species of the family. 16 However, as observed in A. davisi, multiple Ag-NORs have also been reported in A. ibitiensis. 15
In A. argenteus (present study), 45S rDNA was identified in one chromosome pair, as observed in A. affinis, Apareiodon sp., A. piracicabae, A. vladii, P. moreirai, P. hilarii, P. nasus, and P. pongoensis. 16 In A. davisi (present study), A. ibitiensis, and A. vittatus, 16 additional small sites were identified. The large 45S rDNA chromosomal site found in the multiple NOR-bearing species likely corresponds to 45S rDNA-bearing chromosome pair in the single NOR species. Because the chromosome pairs that carry this site differ among Parodontidae species, pericentric inversions, translocations, duplications, and deletions may explain the different morphology in these pairs.
In most Parodontidae species, 5S rDNA is located in a single chromosome pair, 16 as observed in A. argenteus. Additional sites have only been identified in A. vladii cit. Apareiodon sp. 14 and A. davisi (present study). All Parodontidae species exhibit interstitial 5S rDNA sites in the short arm of one chromosome pair, a subtelocentric pair in P. nasus, and a submetacentric pair in the remaining species. This is probably the ancestral 5S rDNA site for Parodontidae because it is conserved among the species.
The overlapping chromosomal sites of 45S and 5S ribosomal sequences in A. davisi are the first to be reported for this family. There is a report of synteny for these genes in P. nasus, with no overlapping sites. 19 However, overlapping of these sequences has been observed in fish species from other groups, including Channichthyidae species, 36 Pimelodus britskii Garavello and Shibatta, 2007, 37 Ancistrus species, 38 Isbrueckerichthys duseni (Miranda Ribeiro, 1907) and Parotocinclus maculicauda (Steindachner, 1877), 39 and Hypostomus species. 40 The occurrence of overlapping ribosomal genes in several A. davisi chromosomes raises questions about the integrity and functionality of these sequences. The arrangement of major and minor ribosomal sequences in different chromosomes is most commonly observed. 41 The separation of these genes is more favorable than synteny 28 because syntenic conditions can lead to disadvantageous rearrangements such as genic conversions and unequal crossing over. 42
Polymorphism of the two ribosomal sequences in A. davisi is first reported for Parodontidae. Cases of polymorphism involving only 18S rDNA are reported in the literature for A. piracicabae—NOR,5,6 A. affinis—NOR, 34 A. ibitiensis, and A. vittatus. 16 Chromosome polymorphisms involving ribosomal genes are also reported for other fish species, such as Oncorhynchus mykiss (Walbaum, 1792) 43 and Rineloricaria lima (Kner, 1853). 44 Polymorphism related to the presence/absence of one small 5S rDNA site observed in pair 17 may be the result of a pericentric inversion. This inversion would carry part of the larger conserved site located near the centromere to a subterminal region of the long arm of the pair. In fish, cases have been identified where inversions have participated in the distribution of ribosomal genes.32,45,46 The polymorphisms associated with the 18S rDNA sequence may be the result of DNA transposition events between non-homologous chromosomes due to the chromosome arrangement in the meiotic interphase and prophase. This would cause equilocal distribution of the sites, which is because these sites present preferential distribution in the terminal region of the chromosomes. 47 In fish, equilocal distribution may have participated in chromosome evolution in other groups, such as Hypostomus. 48 However, for any of these two sequences, the involvement of transposable elements such as transposons and retrotransposons cannot be ruled out.
Sequencing and analyses of rDNAs have highlighted the invasion of transposable elements, mainly in the spacer regions, which contribute to the spreading of these genic families in functional copies or pseudogenes in the genomes.49–53 In the specific case of 5S rDNA, fusion of this gene to parts of the transposable elements could result in short interspaced nuclear elements derived from 5S rDNA.54,55 It has been suggested that the diversification of rDNAs sites in a number of copies and locations in the chromosomes could affect recombination rates and cause karyotype variations in related genomes.53,56 Thus, the diversification of genic families of rDNA in A. davisi may have occurred through the involvement of transposable elements.
The WAp repetitive fraction and satellite DNA pPh2004 are repetitive elements described in Parodontidae and used as important evolutionary markers.8,10 As observed in A. argenteus and A. davisi (present study), all of the species with available data for WAp show that this sequence is dispersed throughout the karyotype, exhibiting differential clusters in species with defined sex chromosome systems. 20
To date, satellite DNA pPh2004 has been identified in the chromosomes of species from the genus Parodon and in A. affinis.8,13,16,20 In the other species of the genus Apareiodon for which there are chromosome studies, no sign of this sequence was detected by FISH.16,17,20 In the present study, the karyotypes of A. argenteus and A. davisi contained satellite DNA pPh2004 sites, suggesting that the origin of this satellite DNA lies in the early diversification of Parodontidae and that this sequence has been eliminated in some species of Apareiodon. Based on a common origin for all sites of the species, this satellite DNA may have originated from only one site, as observed in P. pongoensis 20 and A. davisi (present study). The multiple sites present in the other species may be a derived condition.
The association of repetitive DNAs pPh2004 and WAp allows the identification of proto-sex chromosomes in Parodontidae species, which have no differentiated sex chromosome systems. 20 The proto-sex chromosomes in P. pongoensis and P. nasus correspond to pair 13, which carries the two previously mentioned satellite DNAs. 20 In A. piracicabae and A. vittatus, which have no heteromorphic sex chromosomes, proto-sex chromosomes could not be identified using these probes. 20 In the present study, analyses of the position of pPh2004 and WAp sites in the species studied made it possible to identify potential chromosomes corresponding to pair 13 cited for other species, pair 10 for A. argenteus, and pair 24 for A. davisi.
This study presents the first cytogenetic data for A. argenteus and A. davisi. These results contribute to the knowledge of the karyotype diversity of the family, evident in the occurrence of chromosomal polymorphism in the ribosomal genes, and help understand the role of repetitive DNA in genome and karyotype diversity in this group.
Footnotes
Acknowledgments
The authors are grateful to Prof. Dr. Carla Simone Pavanelli for the identification of the specimens and to Luis Henrique da Silva, Pedro Luis Gallo, and Antônio Donizete da Silva for help with the sampling and technical support. This study was financed by FAPESP (Fundação de Amparo à Pesquisado Estado de São Paulo), CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior), Fundação Araucária (Fundação Araucária de Apoio ao Desenvolvimento Científico e Tecnológico do Estado do Paraná), and CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico).
Disclosure Statement
No competing financial interests exist.