
Abstract
Abstract
Among American cichlids, Geophagus stands out as a species-rich genus widespread over neotropical region. Despite their diversity and confusing taxonomy, only few and basic chromosomal reports are available in populations/species along Atlantic coast, hindering our understanding about evolutionary trends in this genus. Therefore, detailed chromosomal studies were performed in “Geophagus” brasiliensis complex from coastal rivers in northeastern Brazil, totalizing seven populations of Geophagus brasiliensis and one of Geophagus itapicuruensis. All samples shared a diploid number (2n) of 48 divided into 2 submetacentric and 46 subtelocentric/acrocentric chromosomes with (peri)centromeric heterochromatin, hypothesized as a symplesiomorphy for Geophagini. Although G. itapicuruensis and two populations of G. brasiliensis presented single NORs on short arms, multiple GC-rich rDNA sites were observed in the remaining G. brasiliensis samples, ranging from three to six NOR-bearing chromosomes. Inversely, 5S rDNA sites were invariably located at interstitial region on a st/a pair, nonsyntenic to NORs. A compilation of the data set in Geophagus shows that their chromosomal evolution has been driven by pericentric inversions and microstructural changes. Besides, the divergence found in northeastern Brazil places this region as a biodiversity hotspot. A taxonomic revision in the complex “Geophagus” brasiliensis is recommended with the support of cytogenetic analyses.
Introduction
E
Usually, the name G. brasiliensis has been generalized to several locally adapted populations of pearl cichlid found in freshwater and brackish environments along Atlantic coastal rivers of Brazil and Uruguay. 9 Consequently, interpopulation variation in both morphological and cytogenetic features is commonly reported in these fish, suggesting the occurrence of a species complex of controversial taxonomy and systematics. 13 Indeed, G. brasiliensis has been regarded as incertae sedis, 9 as corroborated by molecular analyses that placed this species apart from other Geophagus species.14–17
Several names have also been used to discriminate putative species in the complex “Geophagus” brasiliensis such as Geophagus obscurus (Paraguaçu River basin, Bahia state), Geophagus itapicuruensis (Itapicuru River basin, Bahia state), and Geophagus iporangensis (Ribeira do Iguape River basin, São Paulo state) to name a few. However, in an unpublished thesis, 18 detailed morphological data confirm G. obscurus and G. itapicuruensis as valid species as well as suggest another eight new species (Geophagus sp.n. 1 up to Geophagus sp.n. 8). These results highlight the importance of taxonomic efforts to improve our knowledge about the diversity of these cichlids.
Similarly, cytogenetic reports in the complex “Geophagus” brasiliensis have shown variation in karyotype structure, mainly involving the fundamental number of chromosomal arms (FN = 50–56) despite conserving the plesiomorphic diploid number (2n = 48) of cichlids19,20 (Table 1). Even though heterochromatin in this fish group is usually located at pericentromeric or centromeric regions,13,20,27,35,36 specimens of G. brasiliensis from Jaguaiaíva River (Paranapanema River basin, southern Brazil) diverged from the ancestor pattern once conspicuous interstitial heterochromatin blocks were observed in some pairs. This result allowed differentiating this population from others in nearby basins such as Tibagi River, 36 serving as efficient chromosomal markers.
Size heteromorphism of Ag-NORs.
FN, fundamental number; p, pericentromeric.
Despite all methodological advances in fish cytogenetics, studies using fluorochrome staining or fluorescence in situ hybridization (FISH) are scarce in G. brasiliensis complex. The few available reports show that single NORs labeled with 18S rDNA probes are coincident to CMA3 signals, that is, the major rDNA clusters are interspersed with GC-rich segments, as commonly found in fish.19,35,36 In contrast, 5S rDNA is located at interstitial position on long arms of a single subtelo-acrocentric pair.35,36
Once cytogenetic markers have been commonly used in the genetic analyses of neotropical cichlids, we performed a detailed chromosomal characterization of several populations of the complex “Geophagus” brasiliensis, including the recently validated G. itapicuruensis, from isolated and poorly known river basins through the northern limit of their distribution range. These data were used to infer the chromosomal evolution and the cytotaxonomy of this fish complex, thus contributing to their systematics and management.
Materials and Methods
A total of 95 specimens of G. brasiliensis and 13 of G. itapicuruensis were collected using gillnets in eight localities in Bahia state, northeastern Brazil, comprising Contas River, Recôncavo Sul, Una-Almada, and Itapicuru River basins (Fig. 1 and Table 2). The collection sites in Contas River and Recôncavo Sul basin are within central Atlantic forest regions close to semiarid zones, whereas Una-Almada basin is in a typical coastal Atlantic forest portion, and Itapicuru River encompasses a semiarid region. All individuals were identified and deposited in the fish collection of the National Institute of Atlantic Forest (former Biology Museum Mello Leitão) in Espirito Santo state (G. itapicuruensis: MBML 4033–4034, 4217, 4582–4589; G. brasiliensis: MBML 3998–4007, 4012–4026, 4028–4030, 4032, 4210–4214, 4216, 4365, 4545–4571, 4573–4579, 4581, 4590).
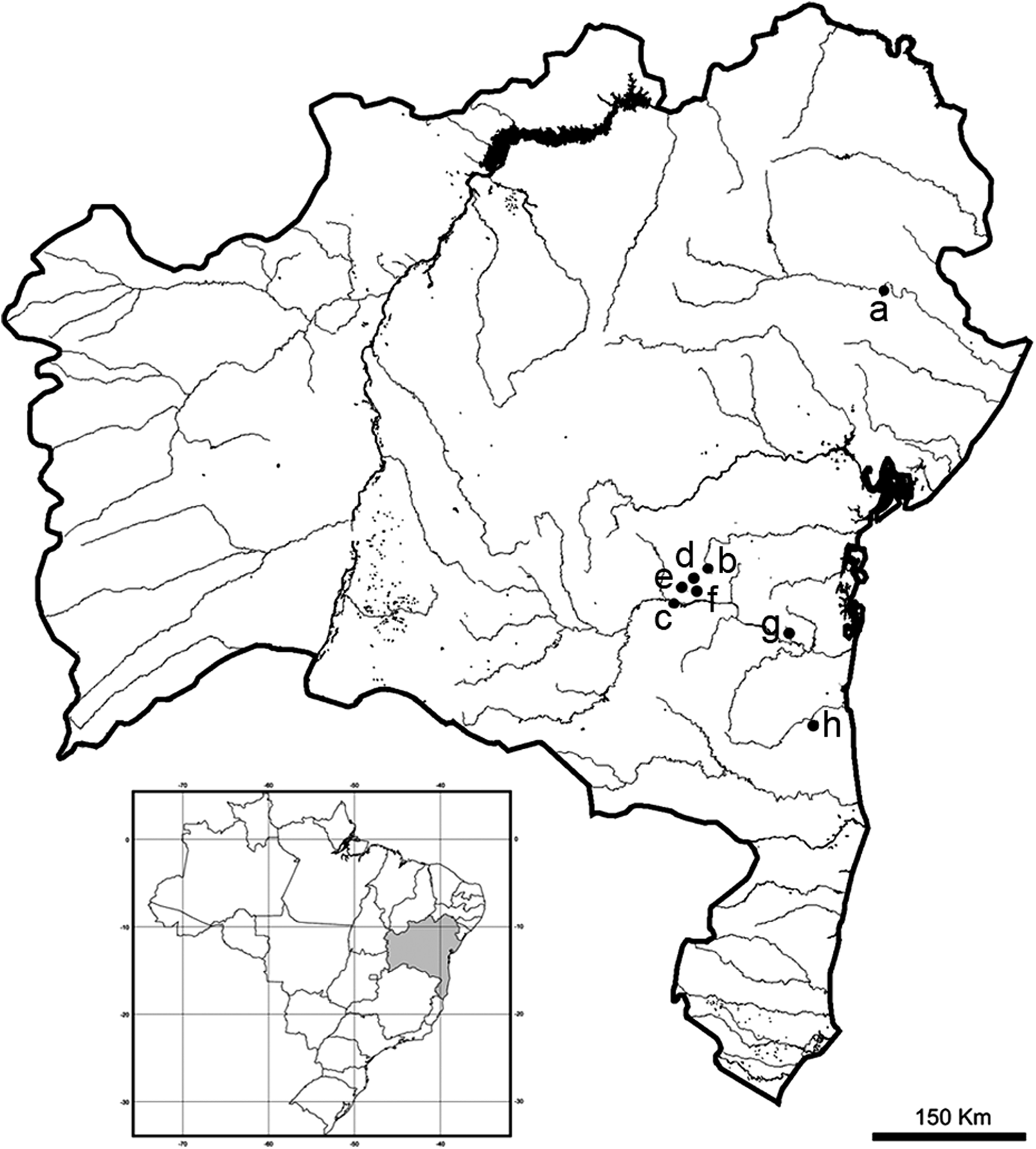
Map of collection sites of Geophagus brasiliensis and Geophagus itapicuriensis in Bahia state, northeastern Brazil: (a) Itapicuru River (Itapicuru River basin), (b) Mineiro River (Recôncavo Sul basin), (c) Contas River, (d) Pedras River, (e) Preto do Costa River, (f) Preto do Criciúma River, (g) Oricó River (Contas River basin), and (h) São Pedro River (Una-Almada River basin).
“?” refer to juveniles individuals of unidentified sex.
Euthanasia of specimens was carried out by immersion in iced water (0–4°C) for, at least, 10 min prior up to cessation of opercular movements. 40 Mitotic chromosomes were obtained from anterior kidney cells according to air-drying method 41 after mitotic stimulation for 24–48 h. 42 Afterward, the chromosomal spreads were stained with 5% Giemsa in phosphate buffer (pH 6.8) for karyotyping. Heterochromatin blocks were visualized by C-banding followed by Giemsa staining. 43 Active NORs were detected by silver nitrate staining. 44 Base-specific fluorochrome staining was performed to identify GC- and AT-rich segments using chromomycin A3 (CMA3) and 4′6-diamidino-2-phenylindole (DAPI), respectively. 45
Double fluorescence in situ hybridization (double-FISH) 46 using 18S and 5S ribosomal probes was carried out under high stringency conditions (77%). Cloned 18S rDNA probes obtained from the genomic DNA of Prochilodus argenteus 47 were labeled with 16-dUTP–biotin (Roche) for further signal detection using FITC–avidin (conjugated fluorescein isothiocyanate–avidin; Sigma-Aldrich). The cloned probe of 5S rDNA was obtained from Leporinus elongatus, 48 labeled with digoxigenin-11-dUTP and detected using anti-digoxigenin rhodamine (Roche). The slides were mounted in VECTASHIELD medium with DAPI (Vector) to counterstain the chromosomes.
Chromosomes were classified according to arm ratio, 49 and the fundamental number of arms (FN) was calculated considering metacentric (m) or submetacentric (sm) chromosomes as biarmed and subtelocentric (st) or acrocentric (a) as one-armed elements. All photomicrographs were obtained using an epifluorescence Olympus BX51 microscope equipped with a digital image system (ImagePro Plus Media Cybernetics) and processed using the software Adobe Photoshop 14.0.
For a more detailed interpretation of cytogenetical data, chromosomal features of each population/species were used to build a dendrogram based on UPGMA (unweighted pair group method with arithmetic mean) available in the software MVSP 3.21 (Multivariate Statistical Package). The data matrix in this analysis included 18 characters as follows: 2n = 48; presence/absence of Ag-NOR on pairs 5, 8, 10, 11, and 18; presence/absence of CMA3 signals on pairs 5, 6, 8, 10, 11, and 18; presence/absence of 18S rDNA signals on pairs 5, 8, 10, 11, and 18; and presence of 5S rDNA sites on pair 9.
Results
All Geophagus samples shared a similar karyotype composed of 2 sm and 46 st/a chromosomes (2n = 48, FN = 50), without differences between males and females (Fig. 2). Small heterochromatin segments (C-bands) were mainly distributed over centromeric and pericentromeric regions in chromosomes of analyzed species/populations (Fig. 3).

Giemsa-stained karyotypes of Geophagus itapicuruensis (a) and G. brasiliensis from Mineiro (b); Contas (c), Pedras (d), Preto do Costa (e), Preto do Criciúma (f), Oricó (g), and São Pedro (h) rivers. The bar equals 5 μm.

C-banded karyotypes of G. itapicuruensis (a) and G. brasiliensis from Mineiro (b); Contas (c), Pedras (d), Preto do Costa (e), Preto do Criciúma (f), Oricó (g), and São Pedro (h) rivers. The bar (bottom) equals 5 μm.
A numerical polymorphism of NOR-bearing chromosomes was identified after silver nitrate staining in the analyzed populations despite the conservative location of Ag-NORs on short arms of st/a chromosomes, comprising pair 5 in G. itapicuruensis (Fig. 4a) and pairs 8, 10, 11, 12, and 18 in G. brasiliensis (Fig. 4b–h). Single Ag-NORs were observed in G. itapicuruensis and in three populations of G. brasiliensis (Contas, Oricó, and São Pedro rivers) (Fig. 4a, c, g, h). In contrast, multiple Ag-NORs were present in the remaining populations of G. brasiliensis ranging from three (Pedras River) to four (Preto do Criciúma River) or five (Mineiro and Preto do Costa) NOR-bearing chromosomes (Fig. 4b, d–f).
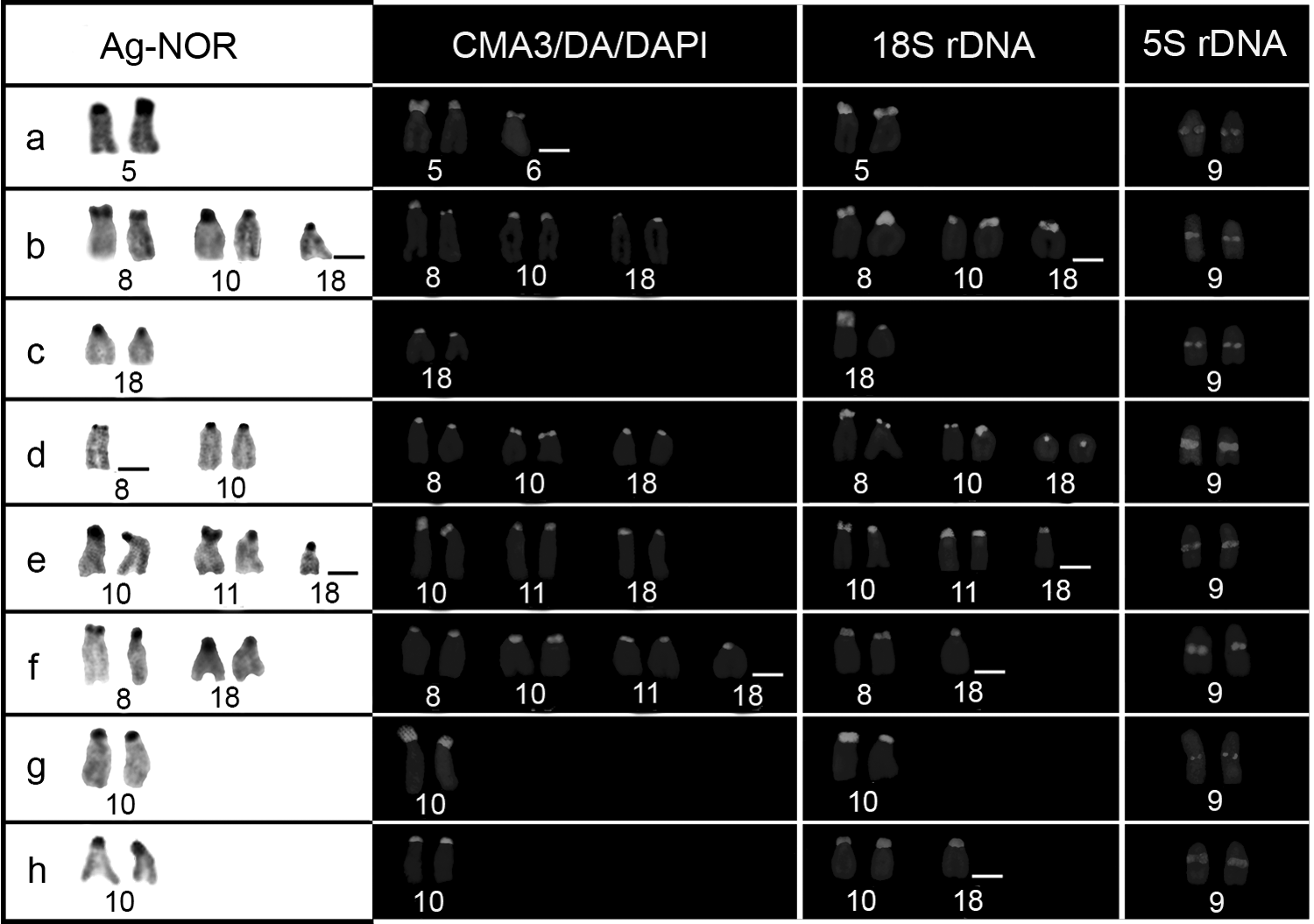
Chromosomes bearing Ag-NORs, CMA3+/DAPI− (GC-rich) regions, and 18S and 5S rDNA sites by double-FISH of G. itapicuruensis (a) and G. brasiliensis from Mineiro (b), Contas (c), Pedras (d), Preto do Costa (e), Preto do Criciúma (f), Oricó (g), and São Pedro (h) rivers.
Base-specific fluorochrome revealed CMA3+/DAPI− signals on short arms of st/a equivalent to Ag-NORs chromosomes in all samples, showing that rDNA sequences were interspersed with GC-rich sites (Fig. 4). However, other GC-rich sites unrelated to Ag-NORs were also identified in G. itapicuruensis and in the populations of G. brasiliensis from Mineiro, Pedras, Preto do Costa, and remarkably, Preto do Crisciúma rivers, comprising one or three additional signals (Fig. 4a, b, d–f).
Similar to silver nitrate staining, FISH using 18S rDNA probes showed that ribosomal loci were variable in populations of G. brasiliensis from northeastern Brazil inasmuch as two to six hybridization signals on short arms of st/a chromosomes were detected, including inactive (Ag-NOR−) rDNA sites and distinct pairs (Fig. 4). This technique proved that the size heteromorphism of NORs indicated by silver nitrate and fluorochrome staining was rather structural (Fig. 4a, e). Similarly, FISH confirmed the single NOR systems in G. itapicuruensis and G. brasiliensis from Contas and Oricó rivers (Fig. 4a, c, g). In the case of samples from Preto do Crisciúma River, the number of FISH signals was inferior to that of Ag-NORs and GC-rich sites (Fig. 4f).
Differently from 18S rDNA, the number of 5S rDNA sites remained unchanged in all populations/species, being located at interstitial region on long arms of pair 9 (Fig. 4). The simultaneous hybridization of both rDNA classes by double-FISH revealed that 5S rRNA genes are nonsyntenic to NORs.
A nonrandom arrangement of chromosomes in metaphase spreads was another common feature in the analyzed samples. Usually, these centromeric associations and bouquet formations involved the NOR-bearing chromosomes (Fig. 5).
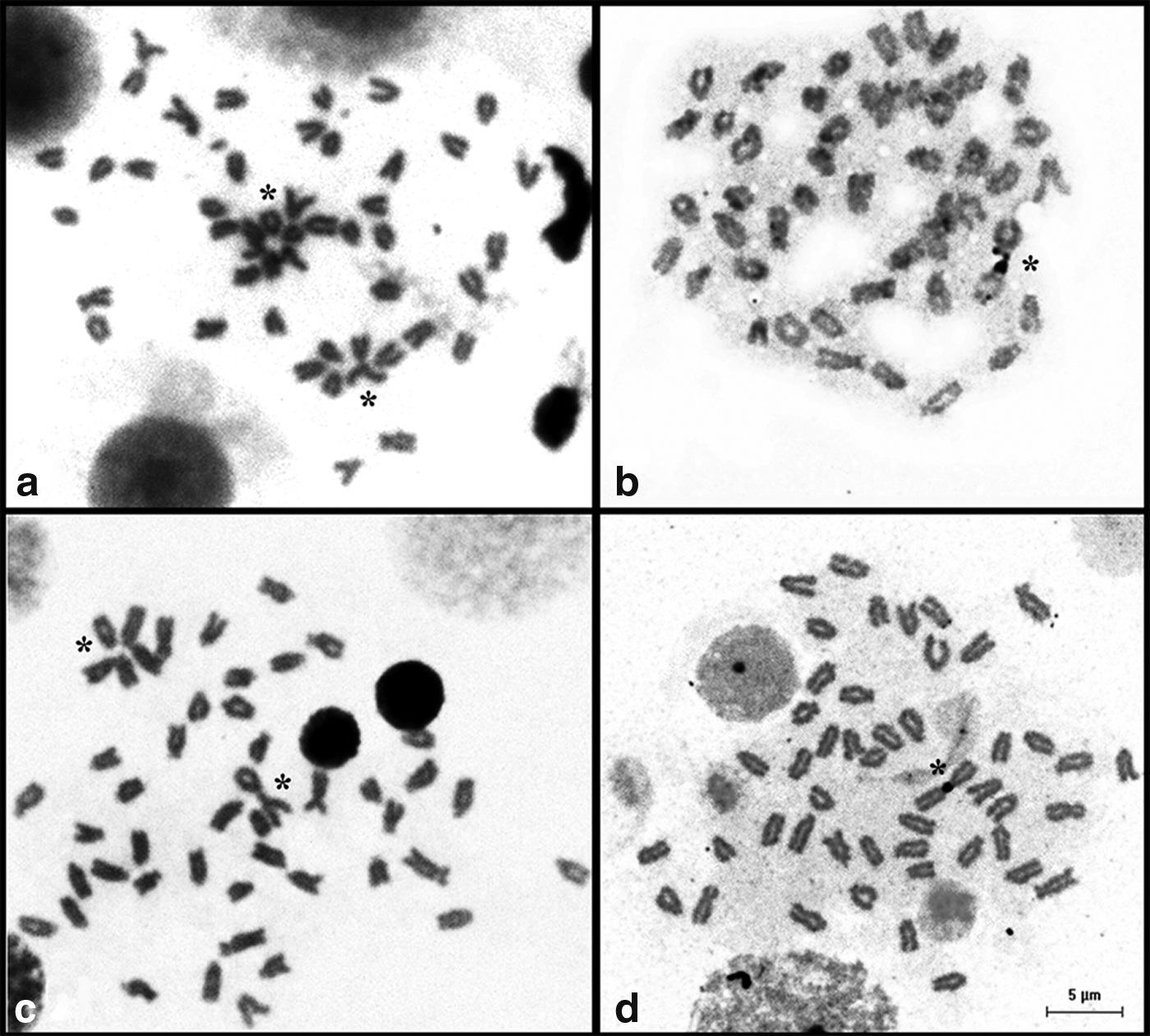
Metaphase spreads of G. itapicuruensis
The UPGMA dendrogram (Fig. 6) showed a clear distinction between G. itapicuruensis and all G. brasiliensis populations. Additionally, different rivers within Contas River basin grouped with other adjacent basins, indicating high diversity levels in this region.

UPGMA dendrogram based on chromosomal features of Geophagus from northeastern Brazil. The letters after G. brasiliensis indicate the collection sites (Co, Contas River; SP, São Pedro River; O, Oricó River; PC, Preto do Costa River; PCr, Preto do Criciúma River; Pe, Pedras River; Mn, Mineiro River).
Discussion
Cytogenetic data are available only for eight genera in Geophagini: Acarichthys, Apistogramma, Biotodoma, Crenicichla, Dicrossus, Geophagus, Gymnogeophagus, and Satanoperca. However, most chromosomal reports in “Geophagus” brasiliensis are restricted to southern populations (Table 1), whereas several northern populations and endemic species of this complex (e.g., G. itapicuruensis) remained unknown from a chromosomal viewpoint.
The presence of 2n = 48 in Cichlidae and other perciforms represents a widespread plesiomorphic feature, 7 as observed in all populations and species of Geophagus herein studied. Within Geophagini, only the genus Apistogramma is known to have diploid numbers lower than 48 chromosomes (Table 1), indicating that chromosomal fusions represent an evolutionary trend in this genus. 1
Contrary to the conservativeness of diploid numbers, distinct karyotype formulae have been reported in Geophagini (Table 1). This variation in chromosomal morphology places this tribe as structurally derived once a karyotype composed only of acrocentric chromosomes (FN = 48) is proposed as a basal condition in Perciformes. 50
All populations and species herein analyzed shared FN = 50, as also reported in G. brasiliensis from São Paulo and Minas Gerais states in Southeastern Brazil,3,4,25,37,38 Gymnogeophagus balzanii and Gymnogeophagus sp.n.21,25 Nonetheless, differentiated karyotype formulae without changes in modal diploid values have already been described in other populations of pearl cichlid (Table 1), emphasizing the role of pericentric inversions during chromosomal evolution of cichlids.7,51 Indeed, inverted chromosomes might be easily maintained in a population once they do not determine significant genetic losses/gains even though this rearrangement can potentially affect recombination and fertility and thus favor reproductive isolation or speciation. 52
The common karyotype macrostructure in populations of Geophagus from these isolated basins and other Geophagini species suggests that the presence of a single m/sm pair is an ancestor condition within this tribe. This hypothesis is corroborated by morphological studies, once Geophagus is considered the genus-type of this Geophagini, being closely related to Gymnogeophagus in phylogenetic analyses. 53 As an additional support, the species of Geophagini with highly derived body shapes, such as the dwarf cichlids of genus Apistogramma and elongated predators of genus Crenicichla bear the most divergent chromosomal formulae compared to the basal karyotype of cichlids (Table 1).
It should be pointed out that the distinguishable karyotype formulae within Geophagini species might be partially influenced by differential chromosomal condensation, thus leading to discrepancies in the classification of chromosomal types. 5 Nevertheless, other variation sources related to external morphology, color patterns, and additional cytogenetic data have been reported in allopatric populations of G. brasiliensis, reinforcing they should comprise a species complex.13,36
C-bands in analyzed samples were mostly evident at centromeric and pericentromeric regions, another simplesiomorphy in Cichlidae.7,20 However, some populations in the complex “Geophagus” brasiliensis from southern Brazil were differentiated by the amount of heterochromatin content. 36 These results reinforce the dynamic evolution of heterochromatin in apparent conserved karyotypes and can be used to track the evolutionary history of species or populations. Unfortunately, several reports in this fish family are focused on a few samples or individuals from uncertain origin (e.g., aquarium trade), which hinders the validation of a putative adaptive role of heterochromatin distribution in cichlids.
So far, single active NOR systems on short arms have been reported in nearly all representatives of the tribe (Table 1). Nevertheless, excepting G. itapicuruensis and G. brasiliensis from Contas, Oricó, and São Pedro rivers, the studied populations presented multiple (three to five) Ag-NORs on distinct pairs (Fig. 4). Until recently, only single NOR systems were described for G. brasiliensis. However, Alves-Silva and Dergam 38 reported the first multiple NOR system in populations from Doce River basin, which along with the populations herein studied reveal a unique diversity to this complex in Atlantic coastal basins.
Actually, differences in the number of Ag-NORs, CMA3+ sites and mapped 18S rDNA regions by FISH were detected in most samples of Geophagus herein analyzed. It is known that silver nitrate staining is successful to reveal only active rRNA genes. 44 This feature could account for the higher number of 18S rDNA signals by FISH compared to Ag-NORs in the populations of G. brasiliensis from São Pedro and Pedras rivers (Fig. 4d, h). Another limitation of silver nitrate staining is the possibility of unspecific marking of heterochromatic regions rich in acidic residues. 54 Even though, both Ag-NOR and FISH techniques are complementary once reduced NORs can be identified when active but remain undetectable by FISH because of the small copy number or hybridization failure of heterologous probes. 2 In this work, the four silver-stained marks and three 18S rDNA FISH signals observed in G. brasiliensis from Preto do Crisciúma River (Fig. 4f) could be explained either by inespecificity of silver nitrate staining or resolution constraints by FISH. Nonetheless, it seems plausible to infer that such incongruence is rather related to insufficient fluorescence by FISH once the signals in some pairs are quite punctiform and hybridization experiment was carried out under high stringency.
In cichlids, NORs are commonly interspersed with GC-rich (CMA3+) sequences. 51 Indeed, this correspondence was detected in all Geophagus samples in this study (Fig. 4). In contrast, the presence of additional CMA3 signals in G. itapicuruensis and populations of G. brasiliensis from Mineiro, Preto do Costa, and Preto do Crisciúma rivers (Fig. 4a, b, d–f) shows that this pattern is not ubiquitous. This result suggests a heterochromatinization process in these samples in which GC-rich segments derived from NOR-associated heterochromatin were amplified and dispersed through the genome.55,56 Such dispersal could be favored by the location of 18S rDNA clusters and GC-rich sites at terminal regions of st/a chromosomes close to pericentromeric heterochromatin.2–4 The common presence of C-bands near centromeres as well as NORs on short arms in Cichlidae5,7 usually gives rise to chromosomal associations in metaphase spreads (Fig. 5). This behavior, common in species with several one-armed chromosomes, 56 would promote the dispersal of 18S rRNA genes and/or GC-rich segments through karyotype.
Differently from 18S rDNA, the distribution of 5S rRNA genes remained invariable in Geophagus from coastal basins in Bahia. Although underrepresented, previous reports using 5S rDNA probes in Geophagini usually reveal signals at interstitial region on long arms of a st/a pair, nonsyntenic to NORs.5,35,36,38 This pattern is similar to that observed in the present study, reinforcing the conservative evolution of 5S rDNA in Geophagini.
Opposed to the conserved karyotype macrostructure, the mapping of 18S rRNA genes and GC-rich segments indicates a derived character in Geophagus from coastal basins in northeastern Brazil, characterizing them as groups with high evolutionary rates at microstructural level. Moreover, the numerical variation of 18S rDNA sites confirms that rRNA multigene families represent one of the most dynamic components in evolution of eukaryote genomes. 57 In fact, both cytogenetic markers were also useful to taxonomical inferences, allowing differentiating G. itapicuruensis from the other samples herein studied, as shown in the UPGMA dendrogram (Fig. 6).
Furthermore, the chromosomal microstructure revealed that populations of G. brasiliensis from Pedras and Preto do Costa rivers (tributaries of Middle Contas River) are more closely related to Recôncavo Sul basin (Mineiro River) (Fig. 4b, d, e) than to other populations in Contas River basin, resulting in distinct clusters (Fig. 6). Again, this pattern indicates a common evolutionary history of both basins, possibly involving headwater capture after dispersal of NORs and GC-rich sites, followed by subsequent geographical isolation by vicariance, as commonly proposed for biogeography of South America fishes.58,59 This suggestion agrees with the hypothesis inferred by the molecular data in a characid fish (Astyanax aff. bimaculatus) from Middle Contas River and Recôncavo Sul. 60
The establishment of the microstructural variation in the karyotypes of analyzed populations can be favored by the territorial behavior and parental care of G. brasiliensis 61 once these traits are likely to restrain vagility and population admixture. Therefore, the samples of G. brasiliensis from coastal basins in northeastern Brazil putatively comprise several chromosomal races of incipient speciation. Additionally, other populations along the wide distribution of G. brasiliensis display high levels of chromosomal variation,37,38 suggesting that this group possibly holds vast cryptic diversity. More importantly, the failure in recognizing these distinct evolutionary units might underestimate the local richness and eventually lead to poor conservation of regional biodiversity.
Finally, the cytogenetic diversity in this fish complex, with microstructural particularities in relation to samples from other South American regions, agrees with the refugia model proposed for drier Atlantic Forest in northern–central Brazil (e.g., Contas River and Doce River basins). According to a combined model of climate reconstruction and genetic studies, it has been inferred that these areas remained relatively stable during Pleistocene glaciation, acting as evergreen and/or species refugia to biodiversity.62,63 In these regions, high genetic variation and accentuated population structure are expected to be reported in different taxa. 64 The differentiated dispersal of ribosomal and GC-rich sites in G. brasiliensis along some of such putative refugia corroborates this association once the lack of local extinction processes would favor increased genetic diversity levels, as also observed in catfish species. 65
In conclusion, the cytogenetic reports in Geophagus highlight the necessity of a thorough taxonomic and systematic revision in this genus to provide a reliable scenario about the diversity of such fish group and their interrelationships. The chromosomal diversity in the complex “Geophagus” brasiliensis also reinforces that the often-neglected coastal basins from northeastern Brazil are a biodiversity hotspot that should be prioritized in biological conservation plans.
Footnotes
Acknowledgments
The authors would like to thank Instituto Chico Mendes (ICMBio/SISBIO) for the license to specimens' collection (no. 26752-1), the Committee of Animal Ethics (CEUA/UESB) from Universidade Estadual do Sudoeste da Bahia for approving the experiments (process 32/2013), and FAPESB and CNPq for the financial support.
Disclosure Statement
No competing financial interests exist.