
Abstract
Abstract
Tissue regeneration requires not only the replacement of lost cells and tissues, but also the recreation of morphologies and patterns. Skin pigment pattern is a relatively simple system that can allow researchers to uncover the underlying mechanisms of pattern formation. To gain insight into how pigment patterns form, undergraduate students in the senior level course Developmental Biology designed an experiment that assayed pigment patterns in original and regenerated caudal fins of wild-type, striped, and mutant, spotted zebrafish. A majority of the WT fins regenerated with a similar striped pattern. In contrast, the pattern of spots even in the original fins of the mutants varied among individual fish. Similarly, the majority of the spots in the mutants did not regenerate with the same morphology, size, or spacing as the original fins. This was true even when only a small amount of fin was removed, leaving most of the fin to potentially reseed the pattern in the regenerating tissue. This suggests that the mechanism that creates the wild-type, striped pattern persists to recreate the pattern during regeneration. The mechanism that creates the spots in the mutants, however, must include an unknown element that introduces variability.
Introduction
T
The reaction-diffusion system proposed by Turing (1952) is a good candidate to mediate the formation of a wide variety of spatially periodic structures, including pigment patterns.1–4 The reaction-diffusion mechanism is driven by two diffusible substances. The first substance is the Activator, which stimulates the production of itself and the production of the second component, the Inhibitor of this Activator. An essential characteristic of this mechanism is that the Activator diffuses more slowly than the Inhibitor. In one of the simplest examples of this system, the ratio of Activator to Inhibitor controls what type of cell or structure will form. For instance, this mechanism could produce skin with spots by regulating pigment cells. Close to the source of the Activator, the Activator/Inhibitor ratio would be high and cells with dark pigment would form. Further away, the Activator/Inhibitor ratio would be lower, and cells with light or no pigment would form. A great variety of pigment patterns can be generated through this basic mechanism, with the ultimate pattern dependent upon the rate of degradation and production of the activator and inhibitor, the rate and direction of diffusion, and the strength of the activating/inhibiting interactions.
Zebrafish have become a prominent model system for studying the generation of pigment patterns because of the availability of mutants and related species with a variety of pigment patterns.5–7 The pigment patterns of WT zebrafish are composed of black (melanophores), yellow (xanthophores), and silver (iridophores) pigment cells aligned in horizontal stripes along the body and continuing into the fins. Experiments using laser ablation are consistent with a reaction-diffusion mechanism regulating pigment pattern formation. When a laser was used to remove all pigment cells from an area of the body of a WT fish, stripes regenerated with similar width and spacing to the original stripes, although the orientation of stripes was no longer strictly horizontal. 8 The pattern of pigment in the regenerated area closely matched a computer simulation of the experiment based on a reaction-diffusion equation. 8
There are also several pathways involved in vertebrate melanocyte differentiation that include a secreted, diffusible factor, including the Wnt, BMP, and Kit pathways. 9 In zebrafish, Kit signaling is required for normal pigment pattern during fin regeneration.10,11 Further, using a mosaic approach, Tryon and Johnson demonstrated that Kit ligand A (Kitlga) can promote pigment formation as much as one fin ray away, suggesting that it is acting as a secreted, diffusible protein. 12
However, other experiments suggest that direct cell–cell interactions play a role. Many of the genes affected in zebrafish pigment mutants are not involved in a pathway involving a secreted factor. For example, the spotted phenotype is the leopard mutant that is caused by a mutation in the cx41.8 gene, which encodes a Connexin protein involved in making gap junctions that link cells. In vitro studies using isolated cells suggest that melanocytes and xanthophores respond to contact with each other with complex cell movements that could drive pattern formation. Whether these cell–cell interactions are part of a reaction-diffusion mechanism is still under debate.
The objective of this study was to test the hypothesis that the pigment patterns in original and regenerating fins would maintain the same pigment pattern. To test this, caudal fins were amputated with different amounts of fin and pigment remaining. Our results gave mixed support for our hypothesis. The original and regenerated fins of the WT fish closely matched one another. One fish that had only two stripes instead of the usual three in the original corrected to have three stripes in the regenerated fin. The pigment patterns among the original leo;lof fins varied greatly, from no spots at all to dozens of spots. Further, in almost every case, the position, density, and spacing of spots in the leo:lof mutants was different between the original and regenerated fin. These results suggest that a mechanism that acts consistently over the lifetime of the fish and from fish to fish controls formation of the normal striped pattern. However, in leo;lof mutants, there appears to be a different or extra mechanism that produces a wide variation in the pattern of spots.
Materials and Methods
Developing the experiment
The Developmental Biology course at the University of Minnesota Duluth is a combination lecture (2 × 50 min/week) and laboratory (1 × 2 h 50 min/week) course. A fin regeneration laboratory, similar to that presented in Fields et al. (2009), had been part of this course for many years. 13 Before starting this project, Dr. Liang used one of the lecture sections to present the basics of the reaction-diffusion model and the beginning idea that grew into this project. This beginning idea was to determine if the spots came back in the same place or in a similar pattern in regenerating fins. In contrast to WT fins, where part of each stripe remains even when a large region of the fin is amputated, in spotted fish it might be possible to compare regeneration in fish with differing numbers of spots/pattern remaining. The students voted in large majority to do original research starting with this basic idea.
The class as a whole agreed upon four different experimental designs and each student chose which design to use. Each student was given one WT fish and one leo:lof mutant, and then was free to either work as a team or individually during all subsequent procedures. The four designs were as follows: a straight cut orthogonal to the long axis of the fish leaving half the spots, a straight orthogonal cut removing all of the spots, a straight orthogonal cut that bisected one of the spots, and an L-shaped cut removing a corner of the fin. The student's WT fin was amputated with the same cut as their mutant. Each student had a 1-L tank for their two fish, and they were responsible for taking digital images of their fish every week or two.
All data were loaded onto a Google Drive folder, so students had access to all of the data as they wrote their papers. Each student wrote a draft of the Introduction, Materials and Methods section, and Results sections, including drafts of figures and tables. Students then divided up the work, with each student choosing to write a revised draft of two pieces of the article (for example, a student could choose to write a Materials and Methods section and one of the subsections of the results along with the corresponding figures). Dr. Liang then used these papers as the sources for this manuscript, changing as little of the original text, figures, and tables as possible. The exception was the Discussion section, which Dr. Liang wrote using detailed outlines submitted by each student.
Anesthetizing the fish, fin amputation, and recovery
Fish were either from the WT strain Zebrafish Danio rerio (Aquatica Tropical) or from the TL strain (ZIRC), which is homozygous for leot1 and lofdt2. Using a net, a fish was moved from a tank into 1 L of 0.017% MS-222 solution and kept there until swimming motion slowed and the fish began to tilt laterally, typically 1–2 min. The anesthetized fish was transferred onto a Petri dish lid so that is was positioned for a lateral picture. The fish and lid were moved onto the stage of a Leica MZ10 F Fluorescence Stereomicroscope. Water was dropped onto the caudal fin to cause it to spread out, and a picture was obtained with a Lecia DFC400 Digital Camera, with the software set at 90% brightness, 0.35 saturation, and 8× magnification. The fish was placed back into the original tank to recover. If the fish did not recover immediately, water was passed over its gills using a transfer pipet until it started swimming. A picture of a ruler was taken at the same settings for use in adding a scale bar and for measurements.
The fish was then anesthetized again, and a new razor blade was used to remove a section of the fin according to the selected experimental design. A second digital picture was taken after the surgery to document the amount of fin removed. The fish was then moved into the original tank to recover as before. Pictures of the regenerating fins were taken every 1–2 weeks after surgery using the same procedure.
The fish were maintained for the first several weeks in an Aquatic Habitats stand-alone rack at room temperature in the teaching laboratory. At this point, there was an accident where the water levels in the rack dropped too low, and many fish died due to micro bubbles forming as the water and air went together through the pump. After this, the fish were moved into the Liang laboratory zebrafish facility, where the water temperature was kept at a constant 28.5°C. To increase the numbers of fin analyzed, eight additional mutant fish were started for each of three experimental designs (straight cut with no spots left, straight cut with half of spots left, and corner cut). The surgery and regeneration of these fish was followed in the same way as the original fish.
Analyzing regeneration
To compare spot pattern among fish, a 3 × 3 mm reference square was placed over the digital image of the original fin that mapped to tissue that was removed during surgery. The same sized box was placed over the image of the regenerated fin in the matching location. The number of spots contained within each box was counted, and the data recorded in a shared table in a class Google Docs folder. ANOVA was performed with JMP 11 Pro software.
Results
Caudal fins of fish carrying the leo mutation had variation in size, shape, and density
Our first goal was to characterize the pigment patterns in WT and leo;lof double mutant caudal fins. To gain a better understanding of the extent of variation, a digital image was taken of each WT and double mutant caudal fin. Consistent with the results of others, all WT caudal fins had similar shapes and stripe patterns (Fig. 1 and Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/zeb). The fins were forked with a prominent dorsal and ventral lobe. In almost all WT fins, there were three prominent, horizontal stripes of black pigment cells, with the first stripe running along the midline between the two lobes and the other two stripes equally positioned dorsal and ventral to this stripe. Some fins had additional shorter stripes starting about halfway into the fin and running across the ventral and dorsal lobes.

Comparison of pigment patterns in striped and spotted caudal fins. WT fins have horizontal stripes of black melanocytes (black arrow) alternating with stripes of yellow xanthophores (arrowhead). In contrast, leo;lof double mutants have spots of black melanocytes surrounded by fields of yellow xanthophores. Lateral view of the fins of live adult fish. Dorsal to the top. Color images available online at www.liebertpub.com/zeb
Due to the leo mutation, the melanocytes (black pigments cells) of the double mutants formed spots instead of stripes and due to the lof mutation the fins were longer than normal (Fig. 1 and Supplementary Fig. S2). The spots of black melanocytes were located in the center of the fin, with more diffuse black pigment toward the dorsal and ventral edges. To characterize the density of spots, a 3 × 3 mm square was placed in the same relative position on each image (Fig. 2). There was great variation in the number and density of spots. The majority of fins had between four and five spots per 9 mm2 (n = 13/24 fins), with an average of 4.8 ± 3.1. However, some fins had no distinct spots, while one fin had 15 spots in the selected region (Fig. 2). There was also variation in the size of the spots. One of the largest spots was oval-shaped with orthogonal diameters of 1.7 and 0.65 mm, area of 0.9 mm2, while one of the smallest was almost a line of cells, and had diameters of 0.58 and 0.14 mm, with an area of 0.05 mm2.

Variation in the pattern of melanocyte spots among leo:lof mutant caudal fins. Lateral views of the adult leo;lof fins used in this study. In each fish, the 3 × 3 mm white box is placed in the same position relative to the border between the fin and flesh of the tail. The number of spots within the box is in the white box next to the fin. The white box in the lower left corner contains the name of the fish. The name is either the initials of the student or, in the case of fish shared among the class, the fish number. The name of each fish is indicated in the lower left corner of all subsequent figures. Three of the spots are circled with a white line to indicate how their size was measured.
Pigment regeneration occurred earlier in WT fins compared to leo;lof mutant fins
The main goal of this experiment was to observe the regeneration pattern of spots in leo;lof double mutant caudal fins and compare them to the patterns in the original fins. Based on previous work that found that WT fins regenerate with patterns very similar to the original fins, we hypothesized that the spots would regenerate with a size and spacing similar to those on the original fins. In addition, we expected fins with more pattern remaining after tissue removal would regenerate patterns more similar to the original fins than those with less pattern remaining. To test these hypotheses, we severed the zebrafish fins in four different ways. In the most severe case, all of the spots in the original fin were removed. In the middle case, approximately half of the fin was removed, leaving only the section of the fin closest to the body. In the least severe case, a corner of the fin was removed, leaving more than one half of the fin and spots intact. Finally, in some cases, a single spot was bisected to test whether the other half of the spot would regenerate.
The fins were imaged regularly during regeneration. In both WT and mutant fins, the new tissue formed first and later became pigmented. By day 7, both the WT and leo;lof double mutants experienced regeneration in the caudal fin, but this tissue was light colored and the border between original and new tissue was apparent, suggesting no melanocytes were present in the regenerating area (Figs. 3 and 4). By day 14, the WT zebrafish began showing signs of pigmentation with the caudal fin becoming darker and sometimes displaying faint stripes. As regeneration continued, the stripe formation continued with the stripes becoming darker and more defined in an anterior to posterior direction (Figs. 3 and 4). Regeneration was judged to be complete by ∼35 dpa, as there were no additional changes in the stripes after this time point.
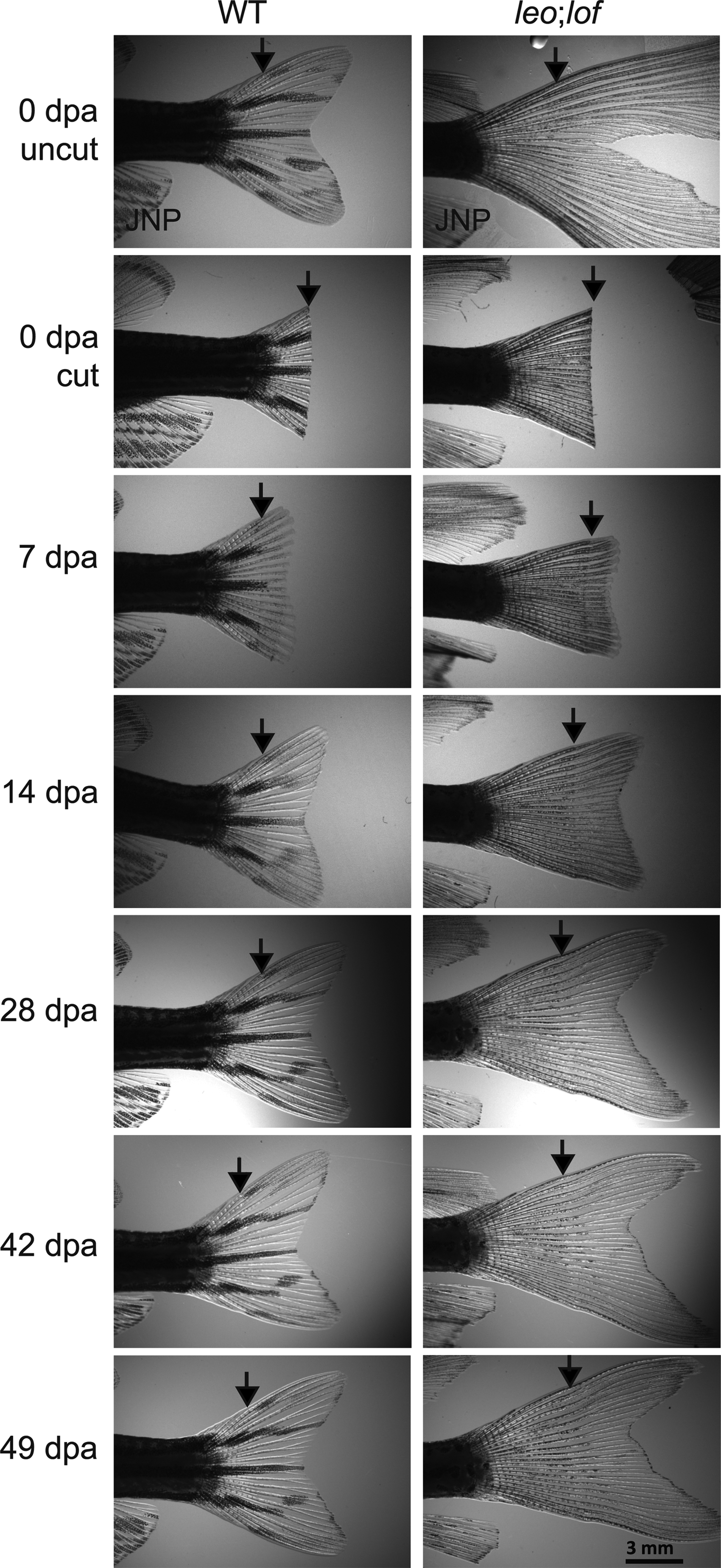
Pigment regenerates faster in WT versus leo;lof fins. Regeneration of fins amputated with a straight cut and, for the leo;lof mutant, half of the spots removed. Arrows point to the point the fin was amputated. Note that stripes are already apparent in the new tissue of the WT fish at 14 dpa and reach the caudal end of the fin by 28 dpa. In contrast, the first spots are apparent in the new tissue of the leo;lof fish at 28 dpa, and pigment has not yet reached the caudal end of the fin by 49 dpa. Lateral views of adult fins. Anterior of the fish to the left and dorsal to the top.
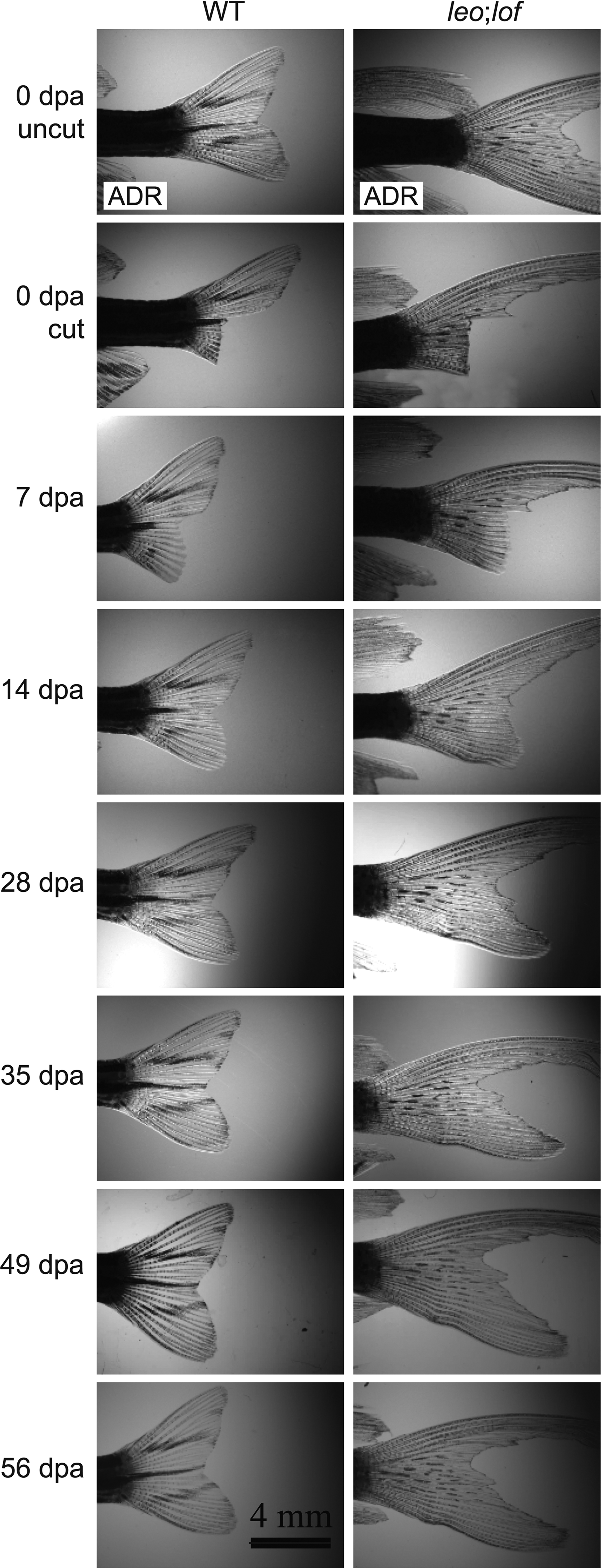
Differences in skin pigment development in regenerating WT zebrafish and leo:lof fins. Regeneration of fins amputated with a corner cut. Stripes are apparent in the new tissue of the WT fish at 14 dpa, reach the caudal end of the fin by 28 dpa, and are complete by 35 dpa. The first spots are apparent in the new tissue of the leo;lof fish at 28 dpa, and pigment has not yet reached the caudal end of the fin by 56 dpa. Lateral views of adult fins. Anterior of the fish to the left and dorsal to the top.
The regeneration time for the leo;lof fins was longer than the WT fins (Figs. 3 and 4). In the leo:lof zebrafish, there was darkening of the newly regenerated fin tissue by day 14, but no distinct spots. The overall shape of the regenerating mutant fins was sometimes disrupted by a kink or bend at the amputation site (Fig. 4). By day 28, the melanocytes in some mutant fins were beginning to develop black spots within the newly regenerated tissue and even very severely amputated fins had regenerated back to a V or lobed shape (Figs. 3 and 4). At day 35, some spots in the new tissue had a density or darkness similar to the preamputated caudal fin, while others were still very faint. At 50+ dpa, some mutant fins still had no spots or very light spots, suggesting that the final pattern was not yet complete, although the semester was ending. Because of this, we continued to take images in the summer and fall. Some fins developed additional spots during this time (Fig. 4), but others never generated any spots (For instance, see fish WLS in Supplementary Fig. S2).
Caudal fins of leo;lof mutants did not regenerate spots in same locations as original fin
As has been reported by others, in most cases the three main stripes of WT fins regenerated in a very similar location compared to the preamputation fin (Figs. 3 and 4; Supplementary Fig. S1). However, there were some cases where the match was not as close (Fig. 5). For instance, one fish began with an unusually striped caudal fin containing only two stripes and a crushed appearance. The regenerated fin was less crushed and had the three horizontal stripes typical of WT fins. In other cases, continuous stripes were replaced by stripes with gaps or vice versa (Supplementary Fig. S1, see fish MNR and JMP).
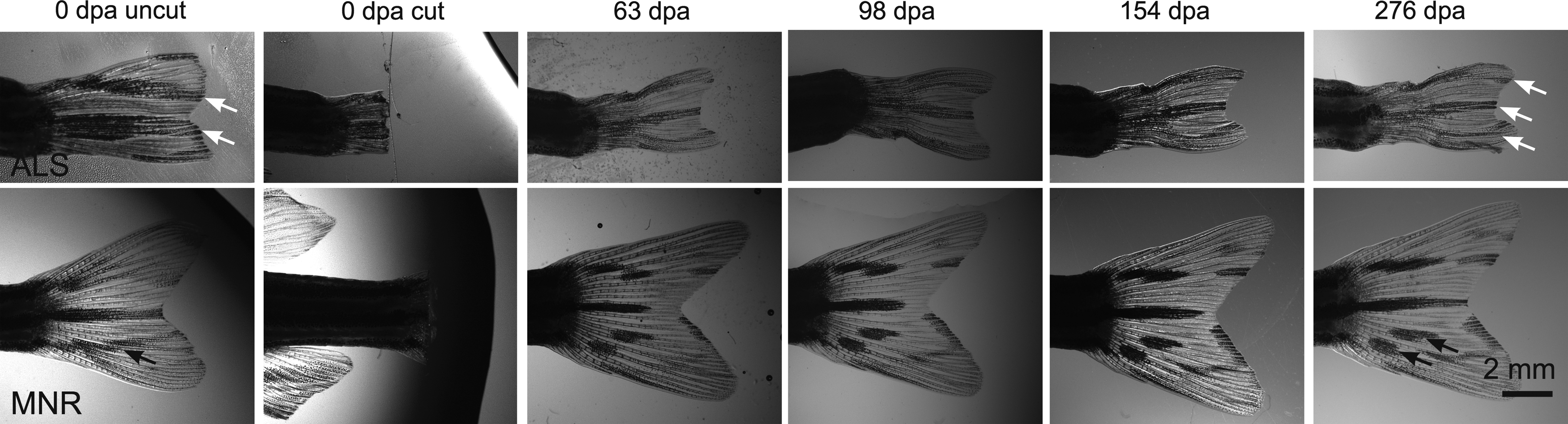
WT fins that regenerated with different pigment patterns than the original fin. The original fin of Fish ALS is abnormal compared with most other WT fins, as it has only two full black stripes (white arrows in 0 dpa uncut image) instead of three. The regenerated fin has a more normal pattern with three full black stripes (white arrows in 276 dpa image). In the original MNR fin, the ventral-most stripe is continuous (black arrow, 0 dpa), while in the regenerated fin, this stripe is interrupted (black arrows, 276 dpa). Lateral views of adult fins. Anterior of the fish to the left and dorsal to the top.
In contrast, in the mutant fish the spots regenerated not only in different locations but also with different sizes, shapes, and spacing (Figs. 3, 4, and 6, Supplementary Fig. S2). To compare the number of spots, a 3 × 3 mm square was placed over the analogous region in the original and regenerated fin and spots in this region were compared (Table 1). The variation in the regenerated fins was similar to that among the original fins, with 2–8 spots in the boxed area of the original fins and 0–12 spots in analogous regenerated area. However, the number of spots within the box was different in most cases, with the majority of the fins gaining spots (Table 1; Fig. 6, Supplementary Fig. S2). There were also other types of variations between the original and regenerated fins. For instance, in one case a spot near the border of excision that was still present after the excision subsequently disappeared (Fig. 7). There was also no consistent pattern among the three fish where a spot was cut in half (Fig. 8). In one of the fish (ADR), the spot elongated into the new tissue and became a size similar to the original spot. In the other two fish (Fish7, Fish8) the regenerating spot did not reach the size of the original spot.

Increased number of spots in a selected region of regenerated leo;lof mutant fins. leo;lof fins amputated with a straight cut, half of the spots removed. The 3 × 3 mm black square is placed in the same relative position on the original fin (0 dp, uncut) and the regenerated fin (276 dpa) and in the spot where all tissue was removed during amputation. The white box next to the black contains the number of spots within the box. Lateral views of adult fins. Anterior of the fish to the left and dorsal to the top.

Loss of spots in the original and regenerated tissue of a leo;lof caudal fin. Dotted line denotes the line of amputation. The caudal fin before excision has four spots. The most rostral spot is circled, and was retained after surgery. A and B and C denote spots in the amputated region. At 63 dpa, none of the spots are apparent. Lateral views of adult fins. Anterior of the fish to the left and dorsal to the top.

Spots cut in half may or may not regenerate back to a full spot. In fish ADR, an elongated spot was cut in half during the amputation. It grew back to almost its original size by 98 dpa. In Fish7, the spot that was bisected expanded a little bit into the new tissue, but did not reach its original size. In Fish8, the bisected spot grew very little or not at all into the new tissue. However, a new spot grew toward the original spot (white arrow). The white ovals or boxes are placed over the same relative position in the sequential images of the same fin. White arrows point to a new spot that formed in the regenerated tissue. Lateral views of adult fins. Anterior of the fish to the left and dorsal to the top.
See Supplementary Figure S2 for images of fins. Only fins that had regenerated more than 60 days are included in the Table.
There did not appear to be a pattern related to the type of amputation (Table 1). For example, in some corner cut fins, there were more spots present, while others did not have any spots regenerate. The same variation was true for the straight cut fins. ANOVA comparing the three types of amputations (corner cut, straight cut with and without spots left) found no significant difference (p = 0.68).
Discussion
Our experiments tested the hypothesis that the pigment patterns in regenerating caudal fins would closely match those in the original fin. Our results from WT fish matched this hypothesis: the stripes of black pigment were similar in number and size in almost all of the fins, whether original or regenerated. However, in leo;lof fish, we found an unexpected amount of variation. The density and size of spots varied greatly even among the original fins of the fish. Further, in most cases, the fins regenerated with a very different pattern of spots than the original fin. This suggests that there must be differences in the mechanisms that control the pattern of pigment cell differentiation or movement between the WT and mutant fish.
Possible explanations for variation in spot patterns
The reaction-diffusion model has been proposed to explain the formation of pigment pattern in zebrafish and many other systems. 4 In this model, the generation of pattern is described by a mathematical equation that describes the diffusion rates and interaction of the molecules involved along with influences from the surrounding biological environment. As many others have also found, there is little variation in the striping pattern among WT fish, and regenerating fins form stripes in places similar to the original fin. 14 This reproducibility of pattern appears to fit well with a reaction-diffusion type of mechanism, which is consistent from fish to fish and can be described by a mathematical equation.
The results in the leo;lof fish were very different. There was a great variation in the size and density of spots among the original fins and between the original fin and regenerated fin in the same fish. The source of this variation is at present unknown. We cannot rule out the idea that a reaction-diffusion type of mechanism is also acting in the mutant fins. Reaction diffusion equations have been generated that produce a spotted pattern very similar to that in the leo;lof fish. However, our results suggest that an additional element would be needed to explain the variation. The additional element could be a dynamic factor in the environment of the fish, such as nutrition, temperature, or crowdedness of the tank. There could also be differences in the genetic background or in the life history of the fish that caused variation. In support of these possibilities, these fish were obtained from the Zebrafish Stock Center, and although they were all the same strain, their life history and parentage was unknown. Further, for the first few weeks of fin regeneration, the fish were kept in a classroom without tight environmental controls.
Another, not exclusive possibility is that the aberrant xanthophore-melanophore interactions in leo mutants could account for the variation in fin spot patterns. Yamanaka and Kondo found a cellular interaction between the black melanophores and yellow xanthophores of WT fish called a run-and-chase movement. 15 When placed together in vitro, a xanthophore extended its pseudopodia toward a melanophore. The cells would then touch, and the melanophore would move away from the xanthophore. This process would repeat, causing a circular movement of the cells. It was proposed that these interactions could contribute to melanophores and xanthophores sorting out into the nonoverlapping, adjacent black and yellow stripes of WT fish. leo mutant xanthophores and melanophores were defective in both the chase and run parts of their interactions: ability of the xanthophore to repulse the melanophore was reduced, as was running away reaction of the melanophore. The authors proposed that in the leo fin, some melanophores would spontaneously aggregate, and become a spot in the middle of a field of yellow cells. 15 If the placement of spots in leo mutants depends on this kind of spontaneous event, then the aberrant cell movements could also explain the variation in spot patterns in the leo;lof fins.
Another possibility is that the presence of the lof mutation in our spotted fish in some way caused the variation in spot pattern. The most prominent phenotype in the lof mutants is a defect in growth control of the fins. 16 WT fins grow isometrically: the ratio of fin length to body length remains constant. lof fins grow allometrically: the ratio of fin length to body length increases over time, resulting in very large fins. The abnormal growing pattern could have contributed to the slower pigment regeneration and also interfered with an underlying reaction diffusion mechanism so that it no longer generated a regular pattern. Unfortunately, the gene affected in lof mutants has not yet been cloned, and so it is difficult to make a more precise hypothesis about how this interference could be occurring. 17
Studies of pigment pattern as a source of original research in laboratory courses
The text and figures created by the students in the class strongly suggest that this course exercise was a success. The text in the Abstract, Introduction, and Materials and Methods sections of this article are modified versions of text and ideas taken from student final papers. The Discussion section was built upon detailed outlines created by the students. Figures 2–8 are modified versions of student figures, and the references (with the exception of recent review articles from 2015) are from articles found and used by students. Students were also able to review and critique their experimental design. For instance, many of the students concluded that including fish carrying lof alone (long-finned, striped fish) and leo alone (short-finned and spotted fish) would have improved their ability to interpret their results.
This project was also a success compared to an earlier original research project done as part of this same Developmental Biology course. In the previous project, students designed several types of experiment that tested the hypothesis that the dark-to-light transition in the morning was the trigger for zebrafish spawning. 18 Although the spawning project also produced publication quality data, it relied on each student repeating the experiment three times over three weeks of the semester. Many students in the class reported that they would have preferred a more regular type of laboratory experience where they used this time to do a diverse set of experiments. In contrast, this project on fin regeneration used the size of the class instead of sequential repeats to validate the results. After the original experimental design and surgery, the subsequent pictures took only a few minutes every 2 weeks. Thus, students did not have to give up any other part of the laboratory experience to do original research. The approach of using the large number of students in a laboratory class to gather data could also be applied to any other project that would benefit from large numbers of parallel experiments.
One of the biggest difficulties is solving the logistical problems of doing original research in a course that meets only periodically. Fin regeneration initially appeared to be a great match for this, as monitoring regeneration every 1 or 2 weeks could capture much of the details of the project. Unfortunately, while the WT fins cooperatively finished regenerating their morphology and pigment pattern within the time frame of the semester, the mutant fins did not. Students had to write their papers with the realization that the experiment was not complete. Fortunately, enough pigment pattern had regenerated in some of the mutants and the students were able to draw the conclusion that the spot patterns were altered in the regenerating fins. However, this points out a second challenge of doing original research in the classroom, you do not know what to expect. Since there is only a limited time to work with, there may not be time to adjust and still give the students a high quality experience.
The history of introducing students to experimental science through research on pigment patterns has a long history. As early as the 1930s, Dr. H. B. Goodrich was bringing students from Wesleyan University to study the biology and genetics of pigment patterns in a wide variety of fish including the zebrafish (then Brachydanio rerio) and medaka [Oryzias (Aplocheilus) latipes].19–21 His work has had a long lasting influence on students and zebrafish research. Steve Johnson tells the story of running into the mother of a classmate who made hybrids between zebrafish and pearl danio (Danio albolineatus) as a high school student in the 1950s, almost certainly connected to the work of Dr. Goodrich. This was at the same time that David Parichy's article on danio hybrids was being reviewed 22 and, for a while, Dr. Johnson worried that his friend's mother had scooped them 50 years earlier (S. L. Johnson, personal communication). Even closer to home, one of the undergraduates who did some of the early work on fish genetics directly with Dr. Goodrich, J. P. Trinkhaus, went on to do foundational work on fish embryology, greatly influencing zebrafish research.23–32 We are honored to be a small part of this long tradition.
Footnotes
Acknowledgments
The authors would like to thank Dr. John Pastor for his excellent guest presentation on Alan Turing and the mathematics of the reaction-diffusion model. Dr. Stephen Johnson and the peer reviewers gave valuable feedback that substantially improved the article. Dr. Stephen Johnson and Dr. Steven Devoto generously shared historical perspectives on this work and information about the research of Dr. H. B. Goodrich. We thank Nicholas Lamon for his outstanding support for our experiments, and the faculty and staff of the Biology Department at the University of Minnesota for creating a learning environment that made this project possible.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.