
Abstract
Abstract
The microsporidium, Pseudoloma neurophilia, is the most common infectious organism found in laboratory zebrafish colonies. Many currently used zebrafish lines originally came from pet store fish, and the initial description of P. neurophilia came from zebrafish obtained from a retail pet store. However, as P. neurophilia has not been described from wild-caught zebrafish, whether P. neurophilia is a natural pathogen of zebrafish is an open question. The pooling of fish of different species in the aquarium fish trade is common and a generalist parasite could be transmitted to novel hosts in this scenario. We determined that P. neurophilia can infect seven species of fishes from five families by cohabitation with infected zebrafish: Betta splendens, Xiphophorus maculatus, Devario aequipinnatus, Pimephales promelas, Oryzias latipes, Carassius auratus and Paracheirodon innesi. Infections in these fishes were histologically similar to those of zebrafish. We include a case report of a laboratory population of fathead minnows with naturally acquired P. neurophilia infections. With such a broad host range, including several fish families, other laboratory fishes should be screened routinely for this and other microsporidian parasites.
Introduction
T
Zebrafish are a very popular tropical fish for home aquariums and there are numerous reports of zebrafish being obtained from retail pet stores and used in research.4,5 Indeed, virtually all zebrafish lines, even those currently maintained in laboratory fish facilities, were originally derived from retail pet store fish. In the ornamental fish industry, it is common for a variety of fishes to be kept in large tanks and allowed to intermingle. It is, therefore, important to consider the potential for infectious diseases to be transmitted from other species of aquarium fishes that are housed with zebrafish or share water within a facility. A microsporidium was initially observed infecting the hindbrain and spinal cords of zebrafish, Danio rerio, from fish that had been obtained from a retail pet store. 5 This parasite was described and named Pseudoloma neurophilia 20 years later. 6 Numerous subsequent reports of P. neurophilia in zebrafish have demonstrated this to be a highly prevalent parasite of laboratory zebrafish. 7 P. neurophilia infections are typically subclinical, making them very difficult to control and potentially introducing nonprotocol-induced variation in research using such fish.3,7
As P. neurophilia has only been described from one fish species since its initial report in 1980, it may be assumed that this parasite has very narrow host specificity. However, the observation of P. neurophilia infecting a moribund Betta splendens that was cohabitating with zebrafish in a personal home aquarium for several months led us to realize that this assumption was incorrect, and thus, we were interested in investigating the ability of this parasite to infect other laboratory fishes. We report here the observations of several fish species infected with P. neurophilia and the occurrence of a natural infection of a laboratory population of fathead minnows, Pimephales promelas. These observations expand the known host range of P. neurophilia to eight species in five families of fishes.
Materials and Methods
Experimental exposures
A total of 40 fishes representing seven species in five families were exposed to P. neurophilia-infected zebrafish and subsequently examined. These fishes were the Siamese fighting fish B. splendens (n = 3), platy Xiphophorus maculatus (n = 5), giant danio Devario aequipinnatus (n = 1), fathead minnow P. promelas (n = 8), medaka Oryzias latipes (n = 10), neon tetra Paracheirodon innesi (n = 9) and goldfish Carassius auratus (n = 4). Cohabitation was performed with 10–15 zebrafish in static aquaria or flow-through tanks at 21°C–27°C. The fathead minnows were obtained from a toxicology laboratory at Oregon State University and the medaka came from Children's Hospital, Boston, MA. The remaining fishes were obtained from a local aquarium store in Corvallis, Oregon. We also included limited unexposed fishes that were obtained from the same respective facilities as follows: medaka (n = 6), betta (n = 4), platys (n = 2), goldfish (n = 2), and fathead minnows (n = 1). No controls were available for the giant danio and neon tetras. These controls were examined by histopathology. The purpose of these controls was to determine whether P. neurophilia or other microsporidia were present in these species before exposure. However, as no rigorously tested P. neurophilia-free colonies of these fish species currently exist, it was not possible to determine whether they were infected with P. neurophilia before experimental exposure.
After cohabitation, fish were euthanized by an overdose of tricaine methanesulfonate (Argent Laboratories, Redmond, WA) and examined by wet mount microscopy and/or histology by Luna stain. 8 Except for the giant danio, a minimum of one fish from each species in which spores consistent with P. neurophilia were observed microscopically was examined using a P. neurophilia-specific assay, either by conventional polymerase chain reaction (PCR) 9 or quantitative PCR (qPCR). 10 The resulting product was sequenced on an ABI Prism®3730 Genetic Analyzer with the BigDye®Terminator v.3.1 Cycle Sequencing Kit (Applied Biosystems, Foster City, CA).
Case description of microsporidian infection in fathead minnows
Fourteen P. promelas (nine males and five females) from a toxicology laboratory population in England were euthanized using an overdose of tricaine methanesulfonate. Ten of the fish (7 males and 3 females) were from a group of 100 (50 males and 50 females) imported 2 months previously from a commercial supplier in the United States. Two weeks after arrival, the fish had bred successfully in designated breeding tanks on a recirculation system, producing a large number of viable eggs. They were then returned to their stock tank (a 100-L tank on constant flow-through using particulate and carbon-filtered water at 23°C). Between each batch of fish, tanks were emptied, cleaned, and disinfected with bleach before repopulation. Recirculating systems were not shared with any other species. Within 4 days, two male fish were found dead, and over the next 2 months deaths continued at a rate of about 2/day, even after the fish were split into smaller groups. Males were predominantly affected, with rapid wasting and color loss in the 24 h before death. By 2 months after arrival (at the time of sampling), 25 male and 2 female fish had died in this group. The four other fish sampled (two males and two females) were remnants of a separate older population at the same facility, imported from the same supplier that had experienced similar clinical problems earlier in the same year. Brains and anterior spinal cords from 10 fish (six from the initial group and four from the historical group) were aseptically collected and stored in 95% ethanol and the remaining tissues were fixed in formalin and processed for histology. Sagittal sections were cut from all fish and stained with hematoxylin and eosin or Gram stain.
The tissues were removed from ethanol for DNA extraction. Approximately 25 mg of neural tissue was extracted using the Qiagen Blood and Tissue kit (QIAgen, Valencia, CA) following the manufacturer's protocol for extraction from tissue with an initial proteinase K digestion at 56°C overnight. PCR was performed using the method described above and the products from three positive samples were sequenced for confirmation.
Results
Spores consistent with P. neurophilia were observed in the spinal cord and hindbrain of at least one representative individual in all seven exposed fishes examined by wet mount and/or histology (Table 1 and Fig. 1A–H). P. neurophilia DNA was detected by PCR and confirmed by sequencing for at least one fish of these species, with the exception of D. aequipinnatus. None of the controls was positive by histological examination. Histological examination of the various fishes except the goldfish showed a similar pattern of infection, as reported previously for zebrafish, with multiple aggregates of spores in the posterior brain and spinal cord and minimal associated tissue reaction. With goldfish (Fig. 1F, G), few spores were detected in wet mounts and histology revealed only small aggregates of spores within granulomas in the meninx adjacent to the spinal cord.

Pseudoloma neurophilia in the central nervous system in fishes exposed to infected zebrafish.
Fish were examined microscopically by wet mount of hindbrain tissue, and whole fish were examined histologically. A minimum of one representative fish from the positive wet mount samples (except for D. aequipinnatus) was examined using a P. neurophilia-specific PCR assay 9 and confirmed by sequencing.
NA, not available; PCR, polymerase chain reaction.
Case description
Microsporidian spores consistent with P. neurophilia were observed in histological sections from 11 of the 14 fish from the laboratory population of fathead minnows that were examined (8 out of 10 of the initial group and 3 out of 4 of the historical group). These were observed as clusters of weakly gram-positive, ovoid spores, ∼2–3 μm in diameter, in the brain, spinal cord (Fig 2A, B), myelinated peripheral nerve, and striated muscle tissues (Fig 2C, D). Five of the seven histology positive fish tested were positive by PCR for P. neurophilia. This was confirmed by sequencing in the three positive samples analyzed.
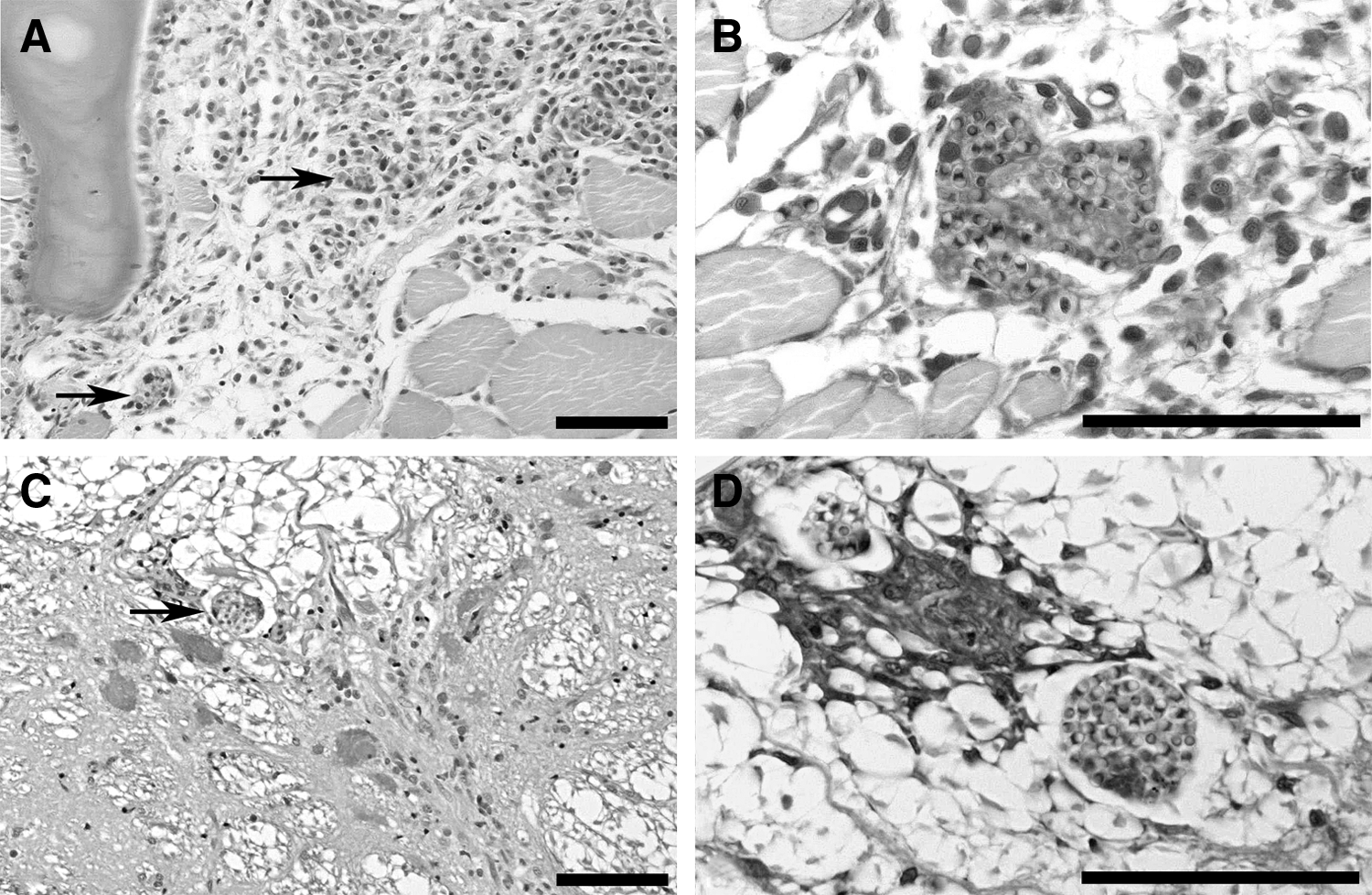
Microsporidian parasites observed in histological sections of fathead minnow, P. promelas.
Discussion
The range of hosts, which a parasite is reported to infect, is often positively correlated with the amount of study that parasite has received. 11 In contrast, there are situations in which the parasite is initially thought to have a very broad host range but is later found to actually represent an assemblage of morphologically similar species. This can especially occur in microparasites that have few morphological characters upon which to base speciation. For example, Loma is a microsporidian genus with relatively narrow host specificity, 12 with Loma salmonae only infecting Pacific salmon Oncorhynchus spp. and not other salmonids, such as Salmo spp. 13
The aquarium fish trade provides many opportunities for the transmission of generalist fish parasites, as multiple fish species are often held together at high stocking densities. Parasites with complex lifestyles, such as trematodes, will generally not survive in such an artificial environment due to the exclusion of intermediate and/or definitive hosts. Parasites with direct life cycles, however, may thrive in such conditions. This concept is best exemplified by Pleistophora hyphessobryconis, a microsporidian parasite frequently found in aquarium fishes. The causative agent of “neon tetra disease,” P. hyphessobryconis infects a broad range of fish hosts and has been reported in 20 species in four different orders of fishes,14–16 including zebrafish. 17 Infection by P. hyphessobryconis often results in clinical signs such as grossly visible lesions and high levels of mortality. In contrast, infections by P. neurophilia are often subclinical and the ability for this parasite to be transmitted vertically within the developing embryo, 18 at least in D. rerio, provides another route leading to the persistence of this parasite in both laboratory colonies and pet stores.
It is important to note that the fishes used in this study are far removed from their native habitat. Even the zebrafish used in most long-established laboratory colonies originated from pet store fish often several decades prior. P. neurophilia has not been reported from wild caught zebrafish, however, it is not known whether they have been examined or to what extent. All of the fish species examined in this study are also used in research. Indeed, the fathead minnows that were naturally infected came from a research facility. Most of the fishes used in this study originated from pet stores, and as we had limited control samples, it is conceivable that some had preexisting infections. Regardless of this alternative, we confirm here that they are susceptible to the infection.
A question remains as to why P. neurophilia infection is so prevalent in laboratory zebrafish populations and whether zebrafish are the original host of P. neurophilia. This microsporidium may demonstrate higher virulence in certain host species. This was not addressed in the current study, which was primarily observational. The lack of controls for various parameters, such as exposure history and age of the hosts, parasite dose, and small sample sizes, precluded the ability to differentiate severities of infection based on host. However, preliminary evidence suggests that goldfish may be more resistant, as the infections were lighter in this species.
We have observed no evidence that any of the host species in the current study was more sensitive to infection nor were differential tissue distributions or mortality rates observed. The nature of the laboratory facility, with fish often held at high density, could certainly play a role in the high prevalence. This could also be exacerbated by recirculating systems, used in many laboratory zebrafish facilities, which may not adequately disinfect effluent water before it is returned to aquaria. In the case study in fathead minnows, the evidence suggests that the parasite was present in fish in two separate importations from the same commercial supplier and may thus be established in commercial breeding populations.
In conclusion, we provide seven species of fishes from five families as previously unrecognized hosts for the microsporidium, P. neurophilia, demonstrated that it infects a broad range of fish hosts. While we present limited evidence for P. neurophilia infection in the giant danio, D. aequipinnatus, given the close taxonomic relatedness of this species to D. rerio, we are confident that the wet mount evidence demonstrates its susceptibility to P. neurophilia infection. In addition, we provide an example of a natural infection associated with clinical disease in a population of laboratory fish, P. promelas. Previous studies have demonstrated the impact of P. neurophilia on zebrafish on common research endpoints such as behavior 19 and fecundity. 20 Similar studies need to be performed on other fish species to determine the impact of nonprotocol-induced variation due to subclinical P. neurophilia infection when they are used in research. These results highlight the need for strict screening of fish populations used for research and the importance of obtaining fish from reputable purveyors with appropriate health and pathogen documentation to mitigate possible effects of chronic parasite infections on laboratory research. They also demonstrate that other fishes, including some that are commonly used in research, may be reservoirs of P. neurophilia.
Footnotes
Acknowledgments
This study was funded, in part, by NIH ORIP 2R24OD010998. The authors thank the Oregon State University Veterinary Diagnostic Laboratory for histological slide preparation.
Disclosure Statement
No competing financial interests exist.