
Abstract
Abstract
Gamma irradiation is commonly used as a bone marrow suppressant in studies of the immune system and hematopoiesis, most commonly in mammals. With the rising utility and popularity of the zebrafish (Danio rerio), gamma irradiation is being used for similar studies in this species. Pseudoloma neurophilia, a microparasite and common contaminant of zebrafish facilities, generally produces subclinical disease. However, like other microsporidia, P. neurophilia is a disease of opportunity and can produce florid infections with high morbidity and mortality, secondary to stress or immune suppression. In this study, we exposed zebrafish to combinations of P. neurophilia infection and gamma irradiation to explore the interaction between this immunosuppressive experimental modality and a normally subclinical infection. Zebrafish infected with P. neurophilia and exposed to gamma irradiation exhibited higher mortality, increased parasite loads, and increased incidences of myositis and extraneural parasite infections than fish exposed either to P. neurophilia or gamma irradiation alone. This experiment highlights the devastating effects of opportunistic diseases on immunosuppressed individuals and should caution researchers utilizing immunosuppressive modalities to carefully monitor their stocks to ensure that their experimental animals are not infected.
Introduction
M
Similarly, Pseudoloma neurophilia, a microsporidium of zebrafish (Danio rerio) that generally produces subclinical infections, was largely ignored as an unimportant background contaminant in many zebrafish research facilities soon after its discovery.5,6 However, as the utility of zebrafish as a laboratory animal has grown, so too has the potential for subclinical pathogens to produce clinical disease, with effects ranging from the addition of nonprotocol-induced variation in important experiments to the loss of valuable laboratory stocks. 7
Pseudoloma neurophilia is a particularly insidious microorganism in that it hides behind the veil of subclinical disease, while propagating in the form of environmentally resistant spores.8,9 These spores resist bleaching at concentrations generally used by facilities that ship embryonated eggs. 10 The parasite can also be transmitted vertically within developing embryos, 9 making the policy of bringing only embryonated eggs into a facility inadequate to avoid P. neurophilia. Between the years 1999 and 2013, up to 75% of facilities submitting fish to the Zebrafish International Resource Center diagnostic service were contaminated with P. neurophilia. 11 While that percentage has decreased in recent years, contamination remains a problem for many laboratories, particularly those that acquire their fish from pet stores or wholesale suppliers without quarantine or sentinel programs to detect subclinical disease.
It has already been established that subclinical infections can produce measurable effects on fish behavior that can adversely influence neurobehavioral experiments. 12 If even subclinical infections interfere with relatively low-impact experimental manipulations, it is logical to assume that experimentation resulting in immune suppression would produce even more damaging effects.
Ramsay et al. 13 examined the effects of stress and cortisol on P. neurophilia infections in zebrafish and found that increased cortisol levels were correlated with more severe infections, reduced weight, reduced fecundity, and increased mortality. 13 While cortisol is notoriously immunosuppressive, other experimental manipulations, such as exposure to gamma irradiation, are purposefully used to destroy hematopoietic tissue and immune function in the test subject. With the emergence and increased use of gamma irradiation protocols and the high prevalence of P. neurophilia in zebrafish research, we must pay special attention to the interactions between this experimental modality and the contamination of facilities with P. neurophilia.14–16
Gamma irradiation exposure protocols are commonly used to study hematopoiesis by elimination of the host immune system followed by transplantation of new cell lines into the depleted tissue. 15 Other laboratories utilize gamma irradiation to study the immune system by analyzing suppression followed by recovery. 17 A facility utilizing gamma irradiation protocols has occasionally observed extensive mortalities following exposure. This facility also had endemic P. hyphessobryconis infections, and it was discovered on postmortem examination that these infections were far more severe, widespread, and inflammatory than is generally observed in nonirradiated animals. 6 P. neurophilia is also endemic in this facility. As this microsporidium is so widespread, it inspired us to investigate the influence of gamma irradiation on the severity of the infections and associated mortality.
Materials and Methods
Wild-type zebrafish of the 5D strain were obtained from the Sinnhuber Aquatic Research Laboratory (SARL) at Oregon State University. This is a specific pathogen-free facility for P. neurophilia. Fish were housed in a flow-through system. The incoming municipal water source was filtered, dechlorinated with charcoal filtration, and heated to 27°C; fish were subjected to a 14/10 h light/dark photoperiod, pH 7.0–7.4, alkalinity 80 ppm, hardness 75 ppm, and conductivity 135 μs. Fish were fed twice daily, but they were not fed on weekends. They were fed the original commercial diet “master mix” used at the SARL. 18 This study comprised two experiments evaluating the interactions between P. neurophilia infections and gamma irradiation by either exposure to the infection and then irradiation (Task 1) or gamma irradiation then infection (Task 2) (Fig. 1).
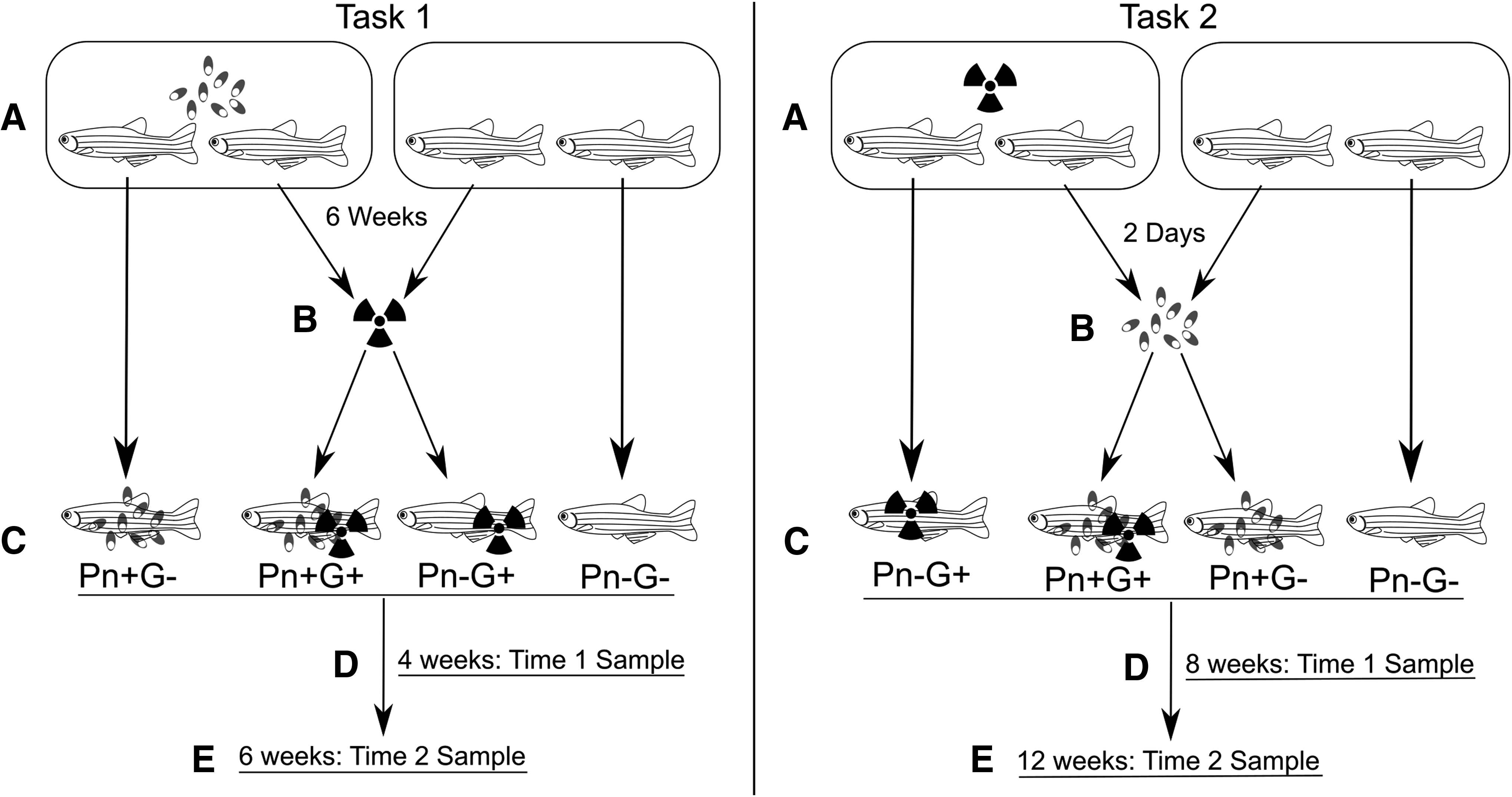
Overview of gamma irradiation/Pseudoloma neurophilia exposure of Danio rerio. Numbers of fish in each group and collection time are noted in Table 1 and in the text. Task 1:
Both of these experiments aimed to mimic potential conditions of a laboratory contaminated with P. neurophilia utilizing gamma irradiation in its research. Hence, both experiments included four groups of fish: controls, Pseudoloma infected without irradiation (Pn+G−), Pseudoloma with irradiation (Pn+G+), and irradiation without Pseudoloma (Pn−G+). Irradiation was achieved using a Gammacell 220 60Co gamma irradiator. Fish were exposed for 250 s to accomplish a dose of 16 Gray (Gy), twice, separated by 24 h. Hence, each exposure gave 15–17 Gy, resulting in a total exposure of 30–34 Gy.
Fish were first sedated with 100 ppm tricaine methanesulfonate (Aregent, Redmond, WA) and then exposed by being placed, ∼20 at a time, in square plastic dishes within the irradiator (100 by 100 mm). Dishes were 15 mm deep with 75 mL of water to just cover the fish with water. Fish were 15 months old at the time of irradiation. Fish were not fed on the day of irradiation to minimize stress. Appropriate groups of fish were experimentally infected with P. neurophilia by exposure to effluent from a tank containing about 150 infected fish maintained in a 15 L tank. A sample of infected fish used for exposure was screened by wet mount of neural tissue before the exposure. They had also been infected at least 6 months earlier and exhibited close to 100% infection.
Task 1. P. neurophilia followed by gamma irradiation
A total of 120 fish were exposed to P. neurophilia for 6 weeks in a 10 gal aquarium. Sixty fish from this tank were then irradiated and 60 were not. They were then placed into twelve 1 L tanks (10 fish/tank). Likewise, 40 uninfected fish were irradiated, and 40 controls (exposed to neither gamma irradiation nor P. neurophilia) were included. These fish were divided into four tanks/group, with 10 fish/tank. Fish were monitored daily, and moribund or dead fish were preserved for histology. Surviving fish were examined at 4 and 6 weeks postirradiation (PI) (10 and 12 weeks postexposure [PE] to P. neurophilia) (Fig. 1).
Task 2. Gamma irradiation followed by P. neurophilia
A total of 50 fish were irradiated and 30 of these fish (three tanks of 10 fish) were then exposed to P. neurophilia effluent starting 3 days after the last gamma exposure. The other two tanks of 10 unexposed fish represented the Pn−G+ group. Likewise, 30 additional fish (three tanks) were exposed to the parasite (Pn+G−) and 20 fish (two tanks) represented the Pn−G− control group. Exposure to the parasite in Task 2 was accomplished by running the effluent of the identical group of infected donors in Task 1 into six 3 L tanks (10 fish/tank). This was done by diverting the outflow of the donor tank into six separate water lines. The water flow to each tank was regulated to be similar into each tank. In addition, the tanks were rearranged every week to different outflow lines to minimize exposure variation.
Moribund and dead fish were collected and preserved for histology throughout the experiment. Many of the Pn+G+ fish had died or appeared moribund by 8 weeks PE (Fig. 1). Therefore, we collected all the remaining fish from this group and those in Pn−G− and Pn−G+ fish, and half of the Pn+G− fish. Only half of the latter were collected at this time point as the infection status was unknown, and we decided to hold some of the fish from this group for an additional 4 weeks to ascertain the development of the infection.
Fish sampling and histopathology
Sampled fish had their coeloms opened and then they were fixed in Dietrich's fixative and decalcified for 48 h using Cal-Ex II. These fish were then sectioned longitudinally and stained with hematoxylin and eosin or the Luna stain using standard methods for histopathologic analysis. 19 Two en face longitudinal sections were examined per fish and the section with the most well-exposed spinal cord was chosen as the section to evaluate. Fish were evaluated by light microscopy to determine percent of fish infected, percent of fish with nervous system infections, percent of fish with extraneural infections, percent of fish with myositis, and the abundance of parasite clusters within fish with nervous system infections.
Statistical analyses
Survival curves for each experiment were generated using Kaplan–Meier estimate. Within-treatment heterogeneity for mortalities was analyzed to determine if there was a significant difference in mortality numbers between replicate tanks within treatments using the log-rank test. If no significant difference was determined, data for the replicate treatment tanks were pooled (Task 1), otherwise mortality data were analyzed separately for each tank within that treatment (Task 2). Survival between treatments was compared for a difference from untreated controls using the log-rank test. Survival analyses were performed in GraphPad Prism version 5.1 (GraphPad Software). Significance was determined by a p value <0.01 (Bonferroni correction to correct for five multiple comparisons).
The differences in the number of P. neurophilia-exposed fish with central nervous system (CNS) infections, myositis, and non-CNS infections were compared between gamma-irradiated and nonirradiated groups using the chi-squared test. The differences in parasite abundance were compared between the two groups using the Wilcoxon–Mann–Whitney U-test. The effect of these histological endpoints on survival of P. neurophilia-exposed fish was estimated using the Cox proportional hazards model to identify independent predictors of survival. Analyses were performed using the statistical programming environment R version 3.1.2 20 and the survival package version 2.38.21,22
Results
Histological examination of control fish (Pn−G− fish) in all groups revealed no significant lesions in terms of myositis or P. neurophilia infections. Please note that in incidents of myositis within test groups, the most characteristic lesion is inflammation and muscle destruction with or without intralesional free spores. Free spores, rather than parasite clusters, are most commonly found in areas of myositis. Generally, when intact parasite clusters were visible in muscle, they were not associated with inflammation.
Task 1. P. neurophilia exposure followed by total body irradiation
No significant difference between replicate tanks within treatments was found, so fish within each treatment were pooled for survival analysis. At 4 weeks PI, which was 10 weeks postexposure to P. neurophilia (PE), there were no mortalities in untreated (Pn−G−) and P. neurophilia-exposed (Pn+G−) groups, and no significant difference was found in the survival of fish treated by total body irradiation (TBI) only (Pn−G+) compared to those exposed to P. neurophilia and subsequently treated with TBI (Pn+G+) (Table 1, 90.5% survival versus 86.2%, respectively). At 6 weeks PE (12 weeks PI), there were no mortalities in the untreated control tanks and no significant difference between survival of the Pn+G− and Pn−G+ groups compared to the untreated controls (Pn−G− fish) (Table 1, 93.3% and 95%, respectively). In contrast, survival within the Pn+G+ group declined to nearly half by 6 weeks and differed significantly from controls (Table 1, 56.7%, p < 0.0001, log-rank test).
CNS, central nervous system P. neurophilia infections; Abundance, mean numbers of P. neurophilia clusters in CNS. Note that abundance includes uninfected fish within the particular populations; Non-CNS, infections by P. neurophilia in other organs. Statistical differences at each time point between irradiated and nonirradiated fish are indicated by *(p < 0.05) using the Wilcoxon–Mann–Whitney U-test for abundance and the chi-squared test for numbers of CNS infections, myositis, and non-CNS infections. ^Marginally significant (p = 0.08).
Histological examination revealed that the prevalence of infection increased from about 20% at 4 weeks PI to about 60%–70% at 6 weeks PI in both the Pn+G− and Pn+G+ fish (Table 1; Fig. 2, Fig. 3). At 6 weeks PI, Pn+G+ fish exhibited over twice as many parasite clusters in the CNS as the Pn+G− fish (p = 0.04, U-test). Moreover, the Pn+G+ fish had five times the prevalence of parasites in non-CNS tissues (p = 0.02, chi-squared) and twice as many with myositis compared to the Pn+G− fish at this time, but the increase in myositis was only marginally significant (p = 0.08, chi-squared).
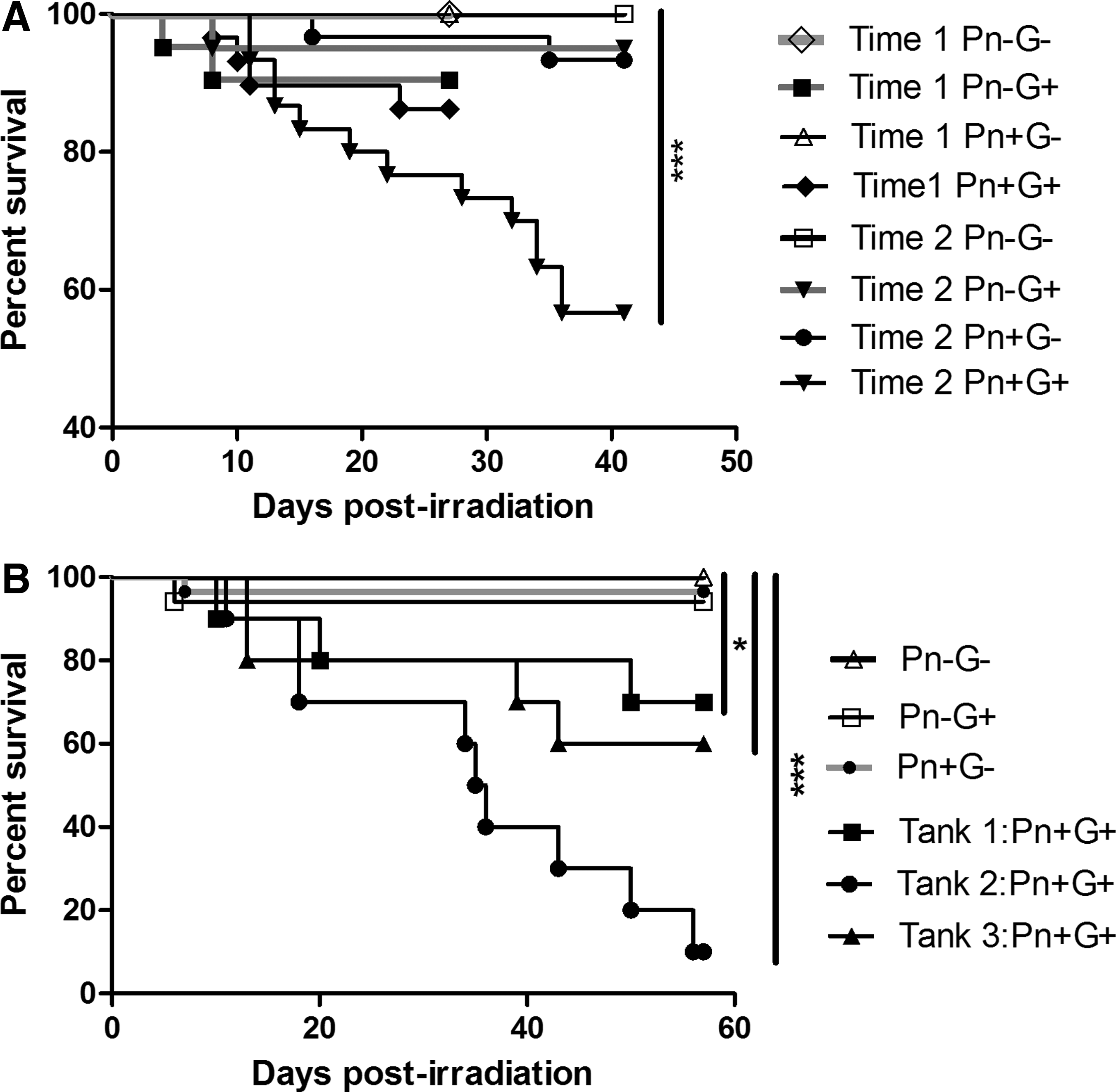
Survival of D. rerio treated with TBI and exposed to P. neurophilia.

Histologic characteristics of P. neurophilia infections in irradiated fish.
Task 2. Total body irradiation followed by P. neurophilia exposure
A significant amount of intertank heterogeneity was detected between replicate tanks of the Pn+G+ treatment group so it was most appropriate to analyze tanks separately (Fig. 2). Similar to Task 1, the Pn+G+ fish showed lower survival (overall 50% at 8 weeks PE), compared to the other groups when fish were collected at 8 weeks PI (Pn−G− = 100%, Pn+G− = 93.3%, Pn−G+ = 94.1%), and hence mortality in the Pn+G+ group was significantly greater than the other groups (Fig. 2. Tank 1 p < 0.0001, Tank 2 p < 0.0001, Tank 3 p < 0.01, log-rank test). Moreover, combining data from tanks together within each group still showed a significant difference (p < 0.001).
Histological examination also showed a similar pattern as in Task 1, with a similar prevalence of infection at 8 weeks PE in both parasite-exposed groups and about thrice the prevalence of non-CNS infections, but this difference was not determined to be significant (p = 0.08, chi-squared). However, in this experiment, both groups of infected fish showed similar levels of myositis. Half of the Pn+G+ fish died before the 8-week sample time, with a mean time to death of 4.5 weeks. Although not collected at predetermined times, we included moribund fish in our analysis to provide a larger data set. Exclusion of clinically infected fish (fish with spinal deformities, profound emaciation, or abnormal swimming behavior) from the analysis showed about 1.5 times the abundance of parasite clusters and a slight increase in myositis (64 vs. 58%), and essentially the same prevalence of non-CNS infections (43 vs. 42%).
The regression analysis performed using the Cox proportional hazards model showed that fish with myositis and extraneural (non-CNS) infections were approximately thrice more likely to suffer clinical signs and death (1.18–7.66 95% confidence interval [CI], p < 0.05; 1.23–7.16 95% CI, p < 0.05, respectively). The prevalence of P. neurophilia was similar within exposed treatment groups and thus this was found to not be a significant predictor of mortality. Overall parasite abundance in the CNS was about thrice greater in the Pn+G+-treated fish than Pn+G−, but this difference was not determined to be significant (p = 0.17, U-test). There was no difference in myositis, but non-CNS infections were about 3.5 times greater in gamma-irradiated fish, which was marginally significant (p = 0.08).
Discussion
In this study, we showed that neither infection nor gamma irradiation alone was associated with significantly increased mortality. In contrast, when combined, either infection followed by irradiation or irradiation followed by P. neurophilia exposure resulted in an increased mortality. Concurrent with increased mortality, we observed more parasites and more extensive infections outside of the CNS, and more myositis in infected irradiated fish. Myositis is particularly associated with immunosuppression with Pseudoloma-infected zebrafish 13 and is often more common in fish experiencing clinical signs associated with P. neurophilia infections. 8
Microsporidiosis has been associated with immunosuppression in a wide variety of hosts, including rabbits, fish, and dogs, and was not recognized as a significant pathogen in humans until the AIDS pandemic.2,4,6 Ramsay et al. showed that fish with higher cortisol levels due to crowding stress had more myositis, which is consistent with our finding of immunosuppression due to gamma irradiation being associated with more myositis. 13 Likewise, gamma irradiation of zebrafish with subclinical Mycobacterium marinum infections exhibited more chronic inflammation and granuloma formation in concordance with more severe infections. 17
A study by Parikka et al is particularly interesting due to the elegant exploration of the mechanisms behind activation of latent M. marinum infections with regard to irradiation-induced immune suppression. Similar to the observations regarding immune deficiency-associated microsporidiosis enhancement, human mycobacteriosis appears to be enhanced by immune suppression, 17 keeping in mind that this study utilized different radiation and sedation protocols than ours. In fact, distinct latent and active phases for M. tuberculosis and M. marinum have both been well described. 17 The Parikka study found that immune suppression in zebrafish by gamma irradiation was associated with a transition from the latent to active phase of M. marinum and an associated increase in mortality and severity of infection. 17
Ramsay et al. reported a very similar finding, in which high mortalities were induced in zebrafish exposed to crowding stress, which had underlying M. marinum infections. 23 While it is generally well known that adaptive immunity is necessary to the inducement and maintenance of M. marinum, using flow cytometry of kidney homogenates, Parikka et al. were able to determine exactly which immune cells were affected by gamma irradiation. 17 They found that leukocyte numbers reached a nadir at 6–7 days PI (at which point numbers begin to recover) and lymphocytes were depleted by ∼80%, while monocyte and granulocyte numbers were only depleted by about 47%. Specifically evaluating lymphocytes, B cells were reduced by 99%, while T cells were only reduced by 67%. Therefore, it appears that, at least in the case of mycobacteriosis, activation of latent bacteria following gamma irradiation is driven primarily by lymphocyte depletion.
Although the specific interactions between the immune system and P. neurophilia are not yet described, the fact that gamma irradiation and infection are associated with increased mortality, extra-CNS infections, and myositis compared to either irradiation or infection alone could indicate some parallels between the mechanism of immune suppression-enhanced microsporidiosis and mycobacteriosis. Both microsporidia and mycobacteria are intracellular organisms. We also see a parallel between the concept of latent and active infections and the switch that occurs during immunosuppression in both infections. Although “latent” and “active” phases have not been identified for P. neurophilia as they have for M. marinum, the general pattern of subclinical minimally inflammatory infections in immunocompetent animals appears to at least partially parallel the mycobacterium narrative.
Taking this parallel further, Parikka et al.'s finding that lymphoid depletion caused by gamma irradiation was primarily responsible for activation of latent mycobacterial infections could explain our own observations that irradiation results in an increased incidence of myositis, a severe inflammatory response microscopically dominated by granulocytes and macrophages. 17 While we do not know the exact mechanism of microsporidial myositis in zebrafish, the general reaction of the body to ruptured P. neurophilia parasite clusters resembles so-called “foreign body” responses in mammals, which are primarily mediated by innate, rather than adaptive, immunity, meaning that lymphocytes may be minimally involved with myositis lesions, at least in their acute active presentation. 24
Another possible explanation for the increased inflammatory lesions seen in our study was that most of our fish were evaluated at several weeks after irradiation. If the nadir of the immune system occurs at 7 days PI, 17 it is possible that P. neurophilia overgrowth began at the immune system's lowest point and continued to progress as the immune system recovered. As more leukocytes were produced and the immune system began to reactivate, it is possible that the severe inflammation we observed in irradiated infected fish was the result of a recovering immune system attacking overgrown ruptured parasite clusters that had been allowed to flourish during the period of immunosuppression. Because lymphocytes appear to be the primary target of gamma irradiation 17 and the incidence of myositis was increased in irradiated animals, adaptive immunity likely contributes to the initial entry of the parasite into the nervous system and the absence of myositis in most immune competent cases. As seen with chronic inflammation associated with ruptured xenomas of Loma salmonae in salmon gills, 25 the major component of myositis in P. neurophilia is likely macrophages, and this cell lineage was likely spared to a large extent following irradiation. 17
Gamma irradiation is not the only immunosuppressive experimental modality used in zebrafish. Studies of stress involving introduction of cortisol or exogenous stressors can be directly immunosuppressive and result in increased mortality and infection severity due to microsporidiosis. 13 N-ethyl-N-nitrosurea (ENU) is an alkylating agent and potent mutagen used in many zebrafish studies to produce random mutations for various genetic studies. Among the studies utilizing ENU are experiments that study immune suppression in induced mutants. 14 Furthermore, repeated, low-level ENU exposure is immunotoxic in male CD1 mice. 26 Therefore, the potential for ENU to directly induce immune suppression in zebrafish exists and may be further complicated by P. neurophilia infection. Contamination of a zebrafish colony with P. neurophilia could also potentially confound observations of embryonic lethal mutations in progeny from ENU-treated individuals, which would otherwise provide valuable information concerning genetic immunosuppression in living individuals.
Gamma irradiation and increased mortality due to secondary infections is well-documented in species other than fish, particularly in humans exposed to the horrors of the atom. Both humans and animals present at such tragedies as Chernobyl, Hiroshima, and Nagasaki have innumerable reported incidences of severe immunodeficiency and secondary infection following ionizing radiation exposure, primarily due to thymic suppression.27,28 These phenomena are so well-documented that the syndromes gain such colorful monikers as “Chernobyl AIDS”. 27 The effects of gamma irradiation on secondary infections is also well-documented in the laboratory mouse, which is susceptible to both latent infection activation and novel infection by otherwise low-virulence pathogens.29,30
There was a general trend of increased severity of infection based on abundance of CNS parasites, non-CNS infections, and myositis. However, even with 2–3 times increase on particular endpoints the change was not always statistically significant. This was likely due to the significant variability within treatment groups. Including moribund or dead fish in the analysis likely underestimates the differences in parasite burden, myositis, and extra-CNS infections as all of these would have likely had higher values if they survived to the appropriate time of collection.
This is supported by the study of Ramsay et al., which showed that parasite prevalence and burden increases with time and there is little if any recovery, at least out to 20 weeks PE. 13 Removal of the moribund or dead fish, which had a mean time to death of 4.5 weeks, resulted in an increase in parasite abundance, but not myositis and CNS infections. This was likely because examination of only healthy fish at 8 weeks represented a group that had more time to develop visible infections. In contrast, myositis and extra-CNS infections were strongly associated with mortality, and thus removing the moribund or dead fish from the analysis probably compensated for any increase in these parameters that would be expected to be related with longer standing infections. 13
In conclusion, our study showed that the combination of P. neurophilia with gamma irradiation was associated with considerably more mortality than either alone. This provides an explanation for the variable mortality observed in a facility that irradiates fish and has an underlying infection by P. neurophilia. This is yet another example of the potential impacts on research when subclinical or chronic infections are wide spread in zebrafish used for research.
Footnotes
Acknowledgments
This study was funded, in part, by NIH ORIP 2R24OD010998-11 to M.L.K. and fish were provided by the Sinnhuber Aquatic Resource Center, Environmental Health Sciences Center NIH NIEHS P30 ES000210. We thank Dr. Scott Menn, Oregon State University Radiation Center, for assistance with irradiation tasks and Dr. Thomas Sharpton for assistance with statistical analyses.
Disclosure Statement
No competing financial interests exist.