
Abstract
Abstract
The objective of the present study was to determine the effects of Roundup on oxidative status in adult Danio rerio liver and gills. Reactive oxygen species (ROS) and antioxidant capacity (ACAP) were measured in fish after exposure to Roundup (5 and 10 mg/L) for 24, 48, 72, and 96 h. Furthermore, gene expression related to antioxidant response was evaluated after 24 and 96 h. In gills, an increase in ACAP was observed after 96 h in the highest concentration. In the liver, a reduction in ROS and ACAP was observed after 24 h, whereas an increase in ACAP was observed after 48 h in the highest concentration. Exposure to the lowest concentration caused a reduction in ROS after 72 and 96 h. Regarding gene expression, a reduction in superoxide dismutase 2 (sod2) and glutathione S-transferase (gstπ) was observed. An increase in uncoupling protein 1 (ucp1) expression was observed in gills of animals exposed to the highest concentration after 24 h. Glutathione peroxidase (gpx) gene expression was reduced in the gills of animals exposed to the lowest concentration; however, it was induced in liver tissue after 96 h of exposure to the highest concentration. These results indicate that zebrafish exposure to Roundup alters oxidative status and causes a response in terms of antioxidant defense system gene expression.
Introduction
T
Roundup has been detected in aquatic environments around the world in concentrations ranging from ηg/L to μg/L.4,5 However, higher concentrations have been observed in the Brazilian southern region, where Roundup is extensively used in cultures of rice and soy. In these instances, glyphosate has been detected from 0.36 to 2.16 mg/L in ponds and rivers. 6
Several formulations of glyphosate-based herbicides are available, all of which contain base ingredients of isopropylamine glyphosate salt, surfactants, water, and other substances that are described as inert ingredients. These formulations vary in terms of glyphosate concentration and the composition of surfactants. 2 Among the surfactants, POEA (Polyethoxylated Tallow Amine) is the most common. Some studies have demonstrated that the commercial formulations are more toxic than the active ingredient per se. Tsui and Chu 7 evaluated the toxicity of the different Roundup constituents to organisms from different taxa (bacteria, microalgae, protists, and crustacean), and they concluded that the organisms presented a higher sensitivity to POEA alone, followed by Roundup, then to isolated glyphosate in its acid form, and the least so to glyphosate salt.
Studies have demonstrated that glyphosate-based herbicides can cause several effects in fish, including genotoxic effects,8–10 histological alterations,11,12 and biochemical and physiological effects.13,14 However, its mode of action is not well described. Some authors indicate that the toxicity of Roundup is based on its capacity to reduce the activity of the enzyme acetylcholinesterase.15–17 In fact, Sandrini et al. 18 have demonstrated that glyphosate is able to inhibit in vitro activity of acetylcholinesterase in Danio rerio and Jenynsia multidentata. However, other studies attribute the generation of oxidative stress to Roundup as a toxicity mechanism.
Oxidative stress is defined as an imbalance of prooxidants and antioxidants in favor of the former, leading to a redox cell-signaling disturbance and subsequent oxidative damage. 19 The antioxidant defense system, both enzymatic and non-enzymatic, is essential in maintaining redox state and in defending cells and organisms against oxidative damage.20,21 This system is highly regulated at the transcriptional level and in terms of proteins that are responsible for the synthesis of antioxidants. In this sense, the Nrf2 transcription factor (NF-E2 Related Factor 2) plays a pivotal role in the regulation of antioxidant defense genes. 22
Superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPx) are enzymes that constitute the first cellular defense against reactive oxygen species (ROS), as these enzymes actively neutralize ROS generated during cellular metabolism. 23 Other important enzymes are the glutathione S-transferases (GST). This enzyme family possesses multiple functions, such as the biotransformation of xenobiotics and the detoxification of damaging oxidative products, and catalyzes the conjugation of these electrophilic substances with the glutathione tripeptide (GSH). 21 GST and other substances, such as ascorbic acid and α-tocopherol, constitute the non-enzymatic antioxidant defense system. 23 These substances interact directly with ROS, mitigating toxic effects. Recently, other substances have been suggested to participate in the reduction of ROS generation. For example, uncoupling proteins (UCPs) may play an important role in oxidative stress regulation by reducing mitochondrial ROS production. 24
Previous studies indicate that Roundup may induce ROS generation and subsequent oxidative damage, such as lipid peroxidation. Moreover, studies have demonstrated an increase in glutathione content and in GPx and GST activities in fish that are exposed to Roundup.16,25,26 There are only a few studies evaluating the effects of Roundup on oxidative status regarding alterations in antioxidant response in terms of gene expression.27–29 The use of D. rerio as a model in toxicological studies is advantageous, as it enables inferences of higher biological complexity with respect to molecular activity, due to the complete characterization of the D. rerio genome. The objective of the present study was to investigate the effects of Roundup on the oxidative status and oxidative gene expression response in D. rerio fish. Specifically, responses related to aforementioned Nrf-2, catalase, SOD, glutathione metabolism (GCL, GPX, and GST isoforms), and UCPs were assessed in D. rerio gill and liver tissue after exposure to Roundup.
Materials and Methods
Fish maintenance and exposure protocol
The animals were obtained commercially from local suppliers and acclimated in aquaria at the Instituto de Ciências Biológicas from Universidade Federal do Rio Grande—FURG (Institute of Biological Sciences, Federal University of Rio Grande) for a minimum of 15 days. Animals were maintained in 100 L tanks (density of one animal/L) with dechlorinated tap water under constant aeration. Water conditions were maintained at 28°C, pH 7.0 with a photoperiod of 14L:10D. Nitrite levels were monitored with a commercial kit and maintained at 0 ppm. The animals were fed ad libitum every day with commercial food (Tetra–Colorbits).
After the acclimation period, juvenile fishes from both genders (234 specimens) were randomly divided into three experimental groups: control group (without Roundup), a group exposed to 5 mg/L of Roundup, and another group exposed to 10 mg/L of Roundup. These concentrations are consistent with previous literature describing oxidative status disturbances in other fish species15,25,26 within the range of glyphosate equivalents observed in environmental studies. Experiments were conducted in glass aquaria (6 L, density = one animal/L), under the same conditions as the acclimation period. The animals were exposed for 24, 48, 72, and 96 h in a static system. No mortality was observed during the experimental period. On the final day of the experimental period, animals were euthanized via immersion in metanosulfonated tricaine (500 μg/L; Sigma) and their gills and liver were dissected for subsequent analyses.
The experimental procedures were approved by the Ethical Committee for Animal Usage (Universidade Federal do Rio Grande; CEUA-FURG protocol No. 23116.007832/2013-88).
ROS quantification
The fluorescent compound dichlorofluorescein diacetate (H2DCF-DA; Sigma) was employed for ROS quantification, in accordance with LeBel et al. 30 Freshly dissected gills and the liver (pool = 2 animals/n = 6 pools per experimental group) were homogenized in 200 μL of cold homogenization buffer (100 mM Tris-HCl, 2 mM EDTA, and 5 mM MgCl2·6H2O; pH = 7.75). The homogenate was then centrifuged (2000 g for 20 min at 4°C). The pellet was discarded, and the supernatant was taken for protein measurement with a commercial kit via biuret assay (Doles Reagents LTDA). To standardize protein content between samples, the supernatant was diluted with homogenization buffer to 2 mg/L of protein. After this, 10 μL of the diluted supernatant was added to a microplate with 127.5 μL of reaction buffer (30 mM HEPES, 200 mM KCl, and 1 mM MgCl2; pH = 7.2) and 10 μL of the H2DCF-DA solution (16 μM). Fluorescence intensity was monitored at 5-min intervals over a 60 min period (25°C) via a fluorometer (Victor 2; Perkin Elmer) with excitation and emission wavelengths of 485 and 520 nm, respectively. The area between the curve of fluorescence/time was integrated, and the total area was determined for comparison. The results were expressed as percent fluorescence area × minute (%FA × min) and compared with the control group (100%).
Antioxidant capacity against peroxyradicals
The diluted supernatants employed for ROS analysis were conserved at −80°C for further antioxidant capacity (ACAP) determination. The ABAP (2,2-azobis2 methilpropionamide dihydrochloride; 4 mM; Sigma-Aldrich) were used as a peroxyl radicals generator. The ACAP of samples were measured with H2DCF-DA in the presence and absence of the ABAP. Fluorescence intensity was monitored over a 35 min period at 37°C via a fluorometer (Victor 2; Perkin Elmer) with excitation and emission wavelengths of 485 and 520 nm, respectively. The difference in fluorescence area with and without ABAP was considered the ACAP. The data were expressed as 1/% relative area compared with the control group, where an increase in ACAP values indicated an increase in the ACAP. 31
Gene expression analysis
Total RNA from liver and gill samples (pool = 3 animals/n = 5 pools per experimental group) were extracted with TRIzol Reagent (Invitrogen) according to the manufacturer's protocol. Total RNA were taken for spectrophotometrical analysis (260/280 nm) and electrophoresis gel analysis (agarose 1%) for quality and quantity determination. RNA samples were then standardized by concentration. The complementary DNA (cDNA) was prepared with the High-Capacity cDNA Reverse-Transcription Kit (Applied Biosystems). The cDNAs were diluted and taken for real time PCR following the manufacturer's protocol from the GoTaq qPCR Master Mix (Promega) and the ABI Prism 7300 Sequence Detection System (Applied Biosystems). The specific primers (Table 1) for zebrafish D. rerio genes were designed in the Primer 3 at the NCBI website (www.ncbi.nlm.nih.gov/). The efficiency of primers was tested, and only those primers with a reaction efficiency higher than 95% were used. The analyzed genes were: nrf2 (Nuclear factor [erythroid-derived 2]-like 2), cat (Catalase—hydrogen peroxide degradation), sod1 (Cu-,Zn-Superoxide dismutase–superoxide anion dismutation), sod2 (Mn-Superoxide dismutase superoxide anion dismutation), gclc (Glutamate cysteine ligase catalytic subunit—glutathione synthesis), gpx (Glutathione peroxidase selenium dependent—hydrogen peroxide degradation), gstα (Glutathione S-transferase isoform alpha—conjugation with glutathione), gstρ (Glutathione S-transferase isoform rho—conjugation with glutathione), gstπ (Glutathione S-transferase isoform pi—conjugation with glutathione), ucp1 (Uncoupling protein isoform 1—mitochondrial proton leak), ucp2 (Uncoupling protein isoform 2—mitochondrial proton leak), and ucp3 (Uncoupling protein isoform 3—mitochondrial proton leak). The tbp (TATA box binding protein) and ef1α (elongation factor 1 alpha) were employed as housekeeping genes. The relative quantification of gene expression was done by the 2−ΔCT method. The results were expressed as fold induction related to the control group (1 standard error).
Statistical analysis
Data were initially assessed to validate analysis-of-variance (ANOVA) assumptions. ANOVAs (with a significance level of 5%) were conducted on data considered normal and homoscedastic to compare groups in the same experimental time. Significant differences were further analyzed via Newman–Keuls post hoc test (with a significance level of 5%).
Results
ROS and ACAP
No significant differences were observed in ROS concentration in gills at any experimental time (Fig. 1A), although a trend of reduction was observed in the first 24 h (p = 0.058). ACAP significantly increased (80%) in animals exposed for 96 h to the highest concentration (10 mg/L) compared with the control group (p < 0.05) (Fig. 1B). A trend of ACAP induction was also observed in animals exposed to the same concentration at 72 h (p = 0.0509).
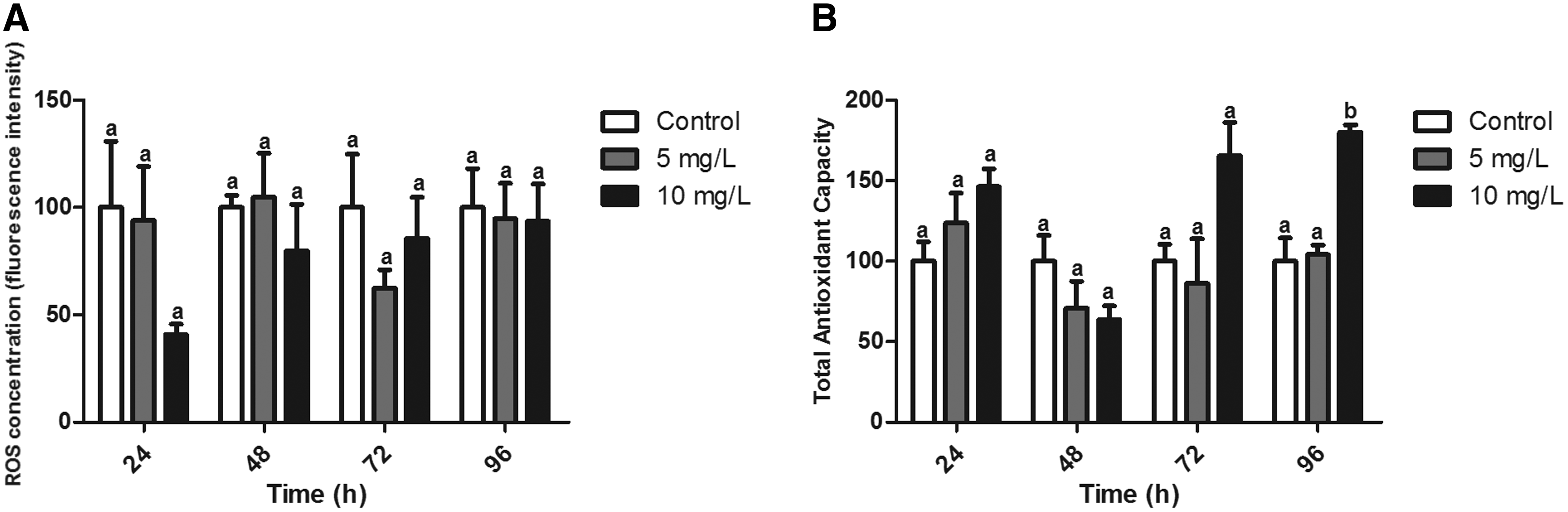
Oxidative balance in gills from zebrafish exposed for 24, 48, 72, and 96 h to Roundup (0, 5, and 10 mg/L).
ROS concentration in the liver significantly reduced in animals exposed to the highest concentration. No significant differences were observed after the 48-h exposure period, although a trend of ROS reduction was observed in both concentrations (p = 0.065). After 72 and 96 h, ROS concentration of animals exposed to the highest concentration returned to control level values. However, animals exposed to the lowest concentration presented reduced levels in both experimental times compared with both the control group and animals exposed to the highest concentration (p < 0.05) (Fig. 2A). A significant reduction in ACAP levels was observed in animals exposed to the highest concentration of Roundup after 24 h (p < 0.05). On the other hand, in a later experimental period, ACAP levels were 1.38 times higher in animals exposed to the same Roundup concentration (p < 0.05) (Fig. 2B).
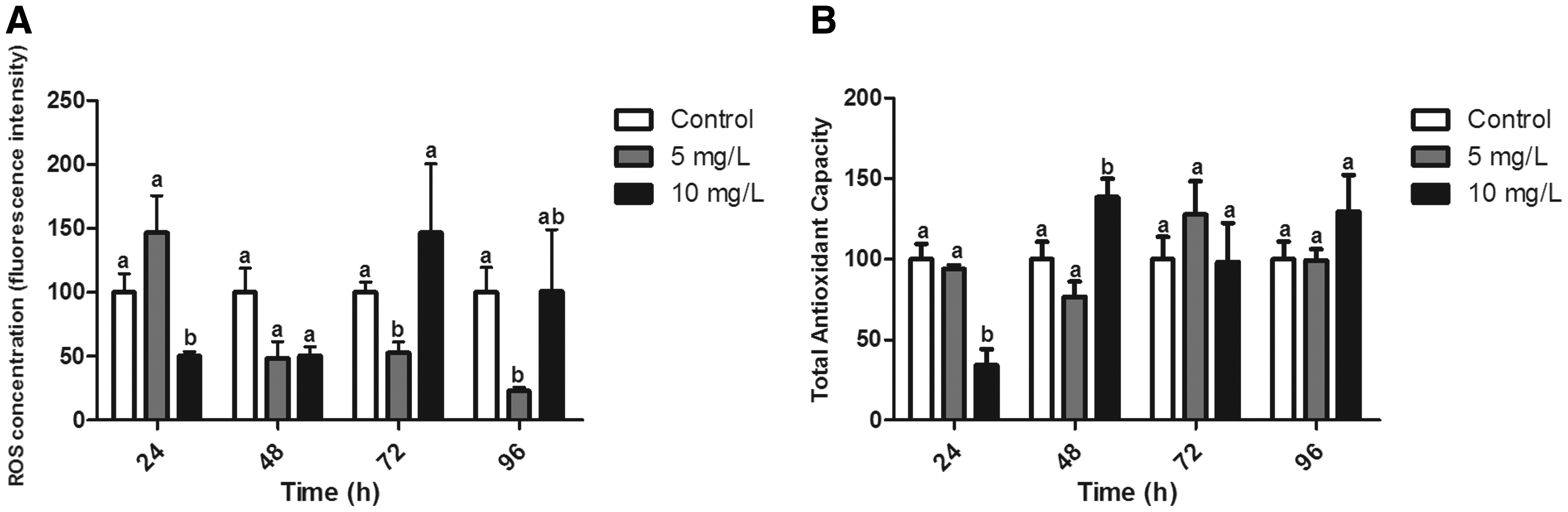
Oxidative balance in livers from zebrafish exposed for 24, 48, 72, and 96 h to Roundup (0, 5, and 10 mg/L).
Gene expression
The gene expressions profiles were evaluated in both gills (Fig. 3) and the liver (Fig. 4) after 24 (A, C, E) and 96 h (B, D, F), respectively, of Roundup exposure. No significant alteration was observed in gene expression of nrf2, cat, sod1, and gclc in gills after Roundup exposure for 24 (Fig. 3A) and 96 h (Fig. 3B) (p > 0.05). However, after 24 h, the sod2 gene expression in gill tissue presented a reduction by a factor of 0.59 compared with the control group (p < 0.05) (Fig 3A). After 96 h of exposure, gpx1a also presented significant differences in its expression (p < 0.05), with a reduction by a factor of 0.81 in animals exposed to the lowest concentration (Fig. 3B). With respect to the three gst isoforms analyzed, only gstρ presented significant differences. In gills from animals exposed to the highest concentration for 24 h, a significant reduction in gtsp concentration by a factor of 0.39 was observed in comparison with the control group and in animals exposed to the lowest concentration (Fig. 3C). A significant difference (p < 0.05) in gene expression of ucp1 was observed in gills of animals exposed to the highest concentration for 24 h. In this group, gene expression increased by a factor of 14 in comparison to the control group and in animals exposed to the lowest concentration of Roundup (Fig. 3E). No significant differences were observed for other experimental times (Fig. 3F).
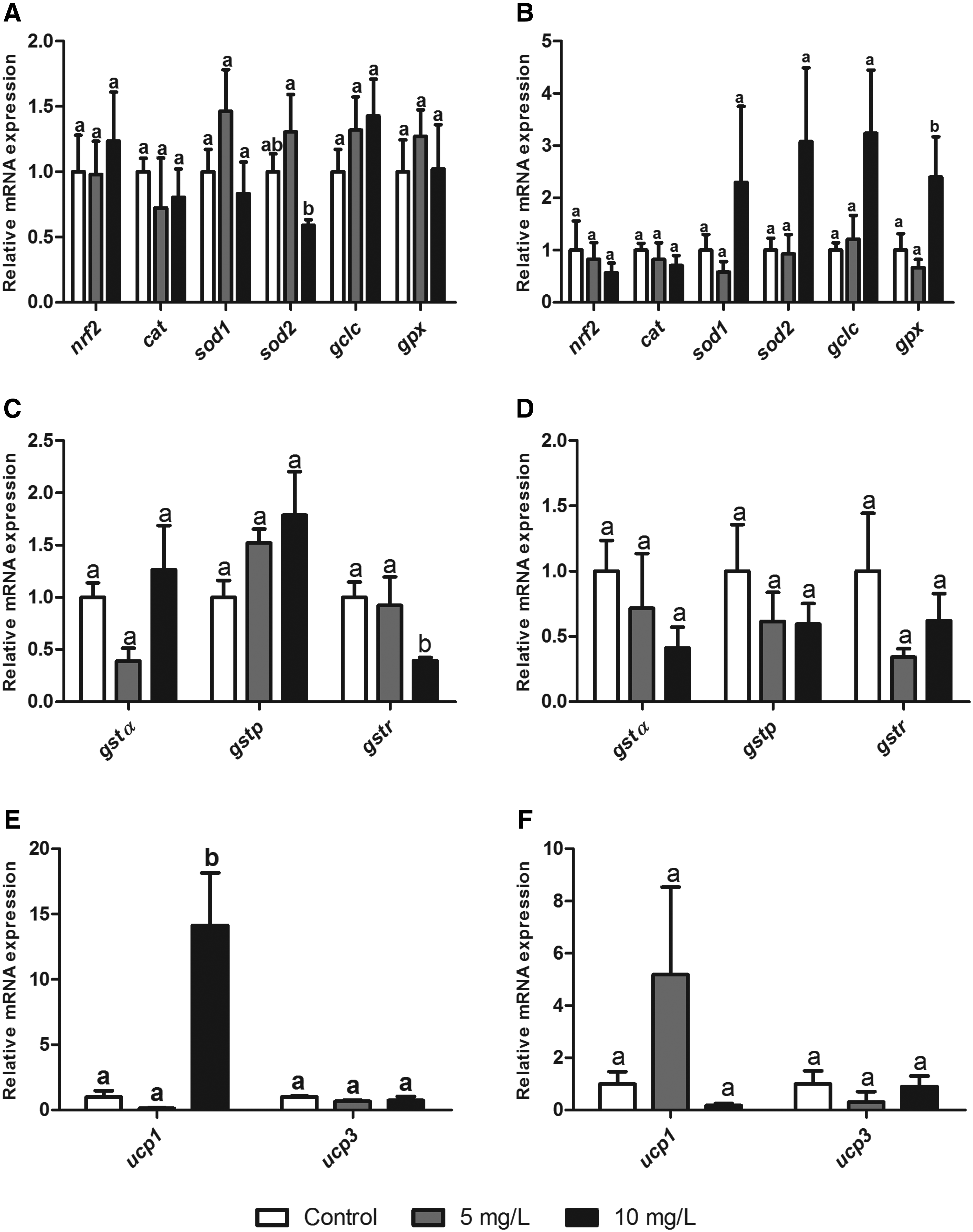
Gene expression in zebrafish gills after 24 and 96 h of Roundup exposure (0, 5, and 10 mg/L).
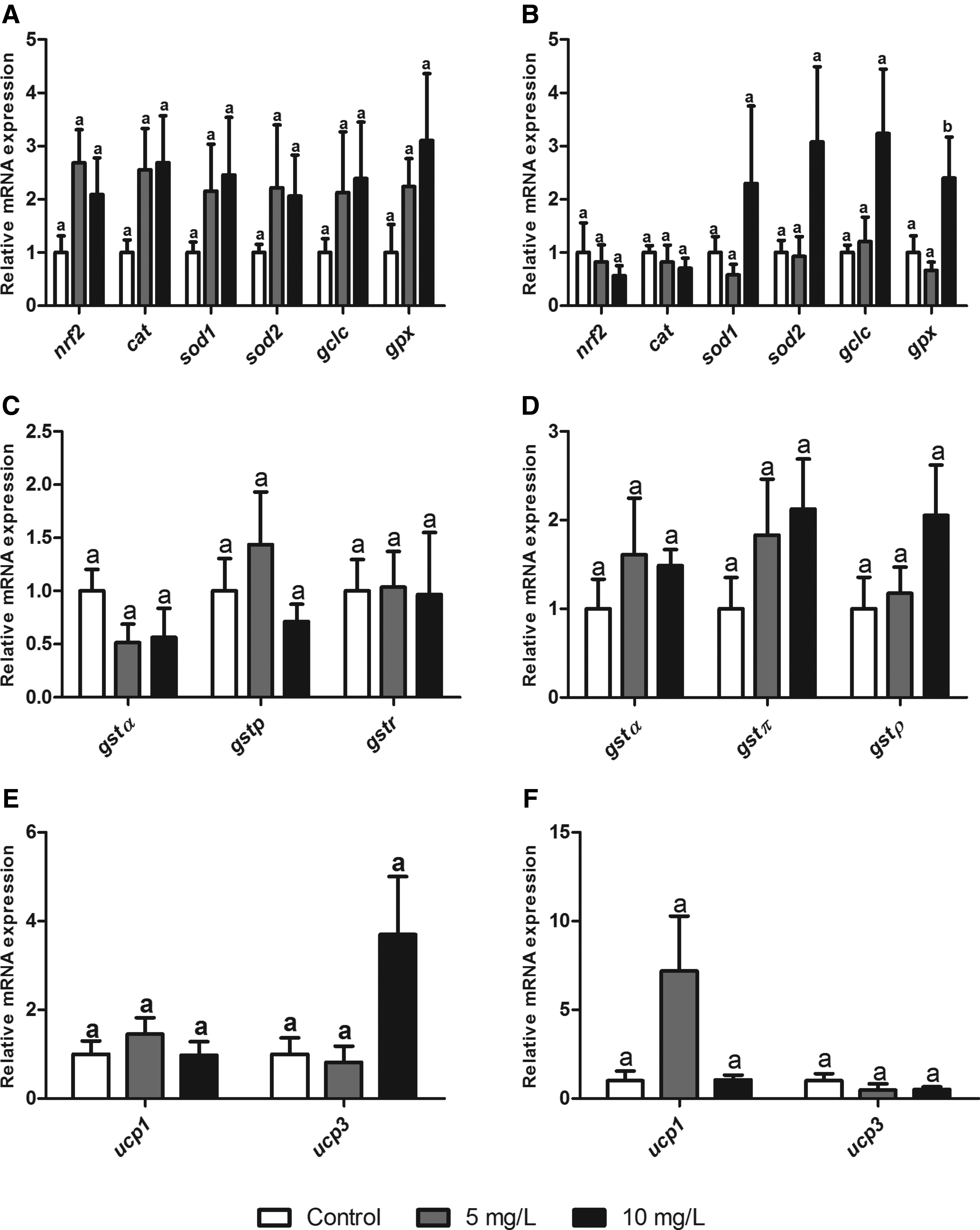
Gene expression in zebrafish liver after 24 and 96 h of Roundup exposure (0, 5, and 10 mg/L).
No significant alteration was observed in gene expression of nrf2, cat, sod1, sod2, and gclc in the liver after Roundup exposure for 24 (Fig. 4A) and 96 h (Fig. 4B) (p > 0.05). However, gpx1a expression presented a significant increase by a factor of 3.23 in animals exposed to the highest concentration after 96 h (Fig. 4B) compared with the control group. No significant differences were observed in gst isoforms in the liver of animals from both experimental periods (Fig. 4C, D). No significant differences were observed for ucp isoforms in both experimental times (Fig. 4E, F).
Discussion
Roundup was recognized in previous literature as a herbicide that can induce harmful effects on aquatic animals. 25 Some studies have demonstrated that its mechanism of toxicity is based on its capacity to reduce or inhibit activity of the enzyme acetylcholinesterase.15–18 Meanwhile, other studies have demonstrated that the mechanism of toxicity of Roundup and other glyphosate-based herbicides is via the generation of oxidative stress. These studies have demonstrated that this herbicide may alter components of the antioxidant defense system, such as enzymes and low-molecular-weight substances, and may cause oxidative damage in biomolecules.25,26,32 However, a global analysis of ROS generation and total ACAP after in vivo exposure to Roundup is not available.
The present study demonstrates that Roundup exposure causes an alteration of the antioxidant status in zebrafish gills and liver. Liver, the main detoxifying tissue against xenobiotics, appeared to be more sensitive and responsive than gills, as significant differences for ROS and/or ACAP were observed in liver tissue across all experimental times. Other studies corroborate this result, demonstrating that the liver is more responsive to Roundup effects when compared with other tissues.15,16,25 This response is due to the fact that enzymes in the liver that act in xenobiotic detoxification processes may become bioactivated to highly toxic forms. 33 On the other hand, gills are a target of pollutants in the water due to their function, anatomical location, and direct contact with the external environment, leading to the rapid absorption of contaminants in water. 34
The gills represent the main entrance for pollutants present in the water column, and, as such, may suffer from oxidative stress. However, ROS generation was not significantly altered in the present study, although a trend of reduction was observed after 24 h of exposure. ACAP presented a later response with a tendency to increase after 72 h. This tendency was confirmed with a significant increase after 96 h in animals exposed to the highest concentration. Harayashik et al. 35 also measured ACAP in gills of Poecilia vivipara exposed to lower concentrations (130 and 700 μg/L) than those employed in the present study, observing no significant differences. Species-specific differences and the concentrations employed may be responsible for this discrepancy.
A reduction in ROS levels in liver tissue was observed after 24 h of exposure to the highest concentration; however, this reduction was observed only after 72 and 96 h of exposure to the lowest concentration. This indicates temporal response patterns between the two concentrations studied. At the highest Roundup concentration, ROS levels returned to control group values after 72 and 96 h. In the lowest Roundup concentration, this return was not observed. This effect of reducing ROS levels may be associated with a mitochondrial dysfunction caused by Roundup, as the mitochondrion is the main site for ROS production. 36 Peixoto 37 observed an uncoupling of oxidative phosphorylation in mitochondria exposed in vitro to Roundup. This effect is likely related to a non-specific mitochondrial membrane permeability, which, in turn, may lead to effects on ROS generation.
A reduction in ACAP levels was observed after exposure for 24 h to the highest concentration, with a subsequent increase response after 48 h. After this, ACAP concentration returned to levels similar to those observed in the control group. Modesto and Martinez 16 have demonstrated that the fish Prochilodus lineatus presented a reduction in the main components of the antioxidant defense system when exposed to Roundup transorb (1 and 5 mg/L), including reductions in enzymes (SOD, CAT, and GST) and glutathione levels in liver tissue. These data corroborate our result of ACAP reduction after the initial exposure period. In fact, another aspect of the study by Modesto and Martinez 16 is that extended exposure to Roundup caused an induction of GSH as a response to the oxidative stress situation, supporting the observed induction of ACAP in the present study. Sinhorin et al. 32 also observed this pattern of response in fish from the genus Pseudoplatystoma. This recuperation in terms of ACAP in zebrafish liver may be related to the non-enzymatic antioxidant defense system, since previous works demonstrate that enzymatic activities in fish liver tissue were, in fact, reduced after Roundup exposure.25,26,32 Taken together, these results reinforce that a broader evaluation such as ACAP presents an advantage in understanding the status of the antioxidant defense system over specific evaluations of individual enzymatic activity.
A concomitant reduction in ROS levels and ACAP was observed in liver tissue after Roundup exposure. As ROS are important factors in cell signaling, this significant reduction in ROS levels would alter cell redox state and lead to disturbances in the antioxidant defense system of animals. 38 ACAP levels increased significantly after 48 h of exposure, although ROS levels remained reduced. Webster and Santos 29 showed that Roundup alters the expression of genes involved in cell cycle, apoptosis, hepatic metabolism, immune response, and antioxidant responses, thus demonstrating that Roundup would alter cell-signaling pathways.
Peixoto 37 demonstrated that Roundup may interfere with mitochondrial bioenergetics, leading to a strong reduction in mitochondrial membrane potential and a reduction in both membrane selectivity and cell respiration. Webster and Santos 29 also demonstrated that Roundup interfered with the expression of genes related to components of the electron transport chain. These described situations would contribute to disturbances in mitochondrial bioenergetics and ROS production, which, in turn, would affect the antioxidant defense system.
The present study tested the hypothesis that alterations in oxidative status in organisms resulting from exposure to Roundup can cause an alteration in the gene expression profile of genes involved in the antioxidant response. The exposure periods of 24 and 96 h were selected, since they represent the exposure periods with the most prominent alterations in ACAP and ROS generation in both gills and the liver. The gene nrf2 codes to Nrf2 transcription factor, and this protein controls and coordinates the expression of genes involved in cell defense such as detoxification enzymes and enzymes/proteins of the antioxidant defense system.22,39 No significant differences were observed in the present study regarding nrf2 concentration between gill and liver tissue after exposure to Roundup at any time. However, the function of this protein is associated with its cellular presence rather than with its gene expression, so the non-significant nrf2 gene expression result does not mean that enzymes and proteins governed by this transcription factor are not regulated.
A reduction in sod2 gene expression was observed in gills after 24 h of exposure to the highest concentration. The protein isoform coded by this gene, Mn-SOD, is located in the mitochondria, the main site for ROS production. This correlates with the reduction in ROS levels, which, in turn, would be associated with a mitochondrial dysfunction caused by Roundup. 37
Another antioxidant enzyme that presented alterations in its gene expression was GPx. This enzyme catalyzes the transformation of H2O2 to H2O using a glutathione molecule as a co-substrate in the process. 23 In gill tissue, a reduction in gpx expression was observed in the lowest Roundup concentration; whereas in liver tissue, an induction in the highest Roundup concentration was observed. Topal et al. observed significant alterations in GPx expression and activity in Oncorhynchus mykiss exposed to a range of glyphosate concentrations (2.5, 5.0, and 10.0 mg/L). 40 Modesto and Martinez 16 observed that GPx activity was enhanced in liver tissue of P. lineatus exposed to 5 mg/L Roundup, corroborating results from the present study. Based on these results, GPx should be considered a good indicator regarding Roundup exposure.
The expression profile of glutathione S-transferase isoforms was observed and in particular, the expression of gst1α, gstπ, and gstρ in both gill and liver tissue from zebrafish was observed. GSTs conjugate with electrophilic substances such as xenobiotics and damaging oxidative products, subsequently producing the less toxic tripeptide glutathione and facilitating excretion.9,41 Some studies have evaluated total GST activity after Roundup exposure. In P. lineatus, GST activity was induced in liver tissue of fish exposed to Roundup (10 mg/L) for 24 and 96 h. 16 In another study with the same species, the authors described an initial reduction of GST activity after 6 h of exposure to Roundup. 15 The same trend was observed in Carassius auratus exposed to various Roundup concentrations (2.5, 5, 10, and 20 mg/L). 25 These results corroborate the reduction of gstρ observed in the present study for gill tissues after Roundup exposure for 24 h. Glisic et al. 42 analyzed the various functions of the GST enzyme family in D. rerio. The authors suggested that isoforms not assessed in the present study were more related to biotransformation of xenobiotics. Further, the authors indicated that isoforms such as gstρ are more involved in other physiological process, including conjugation of lipid peroxidation products. This role is also attributed to other GST isoforms, such as those analyzed in the present study.
Control of ROS production at the mitochondrial level represents an important aspect of oxidative status. UCPs are present in the inner mitochondrial membrane and uncouple the electron transport chain from the oxidative phosphorylation process. 43 Five isoforms have been described in mammals, with different degrees of identity, tissue distribution, and functions. 44 These proteins are involved in a range of physiological processes, including non-shivering thermogenesis and cell signaling; whereas a more recent approach has suggested a role in the attenuation of ROS production at the mitochondrial level and subsequent mitigation of oxidative stress. 45 The present study produced an interesting result, wherein an increase in expression (by a factor of 14) of ucp1 was observed in zebrafish gills exposed to Roundup for 24 h at the highest concentration. This UCP isoform is classically associated with adaptative thermogenesis in mammal tissues. 46 However, the presence of this protein in fish and other animals that do not regulate body temperature suggests another function. 47 There are no data describing ucp1 gene expression after xenobiotic exposure. However, the result observed in the present study may be related to the reduction in ROS levels after 24 h of Roundup exposure to the highest concentration.
Conclusion
The results of the present study demonstrate that exposure to Roundup leads to a disturbance in the oxidative status of zebrafish gill and liver tissues. This effect characterizes an alteration in cellular redox state, which, in turn, would cause disturbances in cellular signaling pathways related to antioxidant response. These results are corroborated by previous studies regarding enzyme activities and gene expression profile related to components of the antioxidant defense system. Further, Roundup exposure interferes with genes that are not classically involved in the antioxidant defense system, such as ucp1, which, in turn, may contribute to an indirect restoration of the oxidative balance.
Footnotes
Acknowledgments
R.R.V., Fabio de Mello Tarouco, and Amanda Silveira Guerreiro are graduate students and fellows to the Brazilian agency CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior). The authors would like to thank CNPq (Conselho Nacional de Pesquisa process 480919/2013-5) and the National Institute of Science and Technology—Aquatic Toxicology (INCT-TA) for their financial support.
Disclosure Statement
No competing financial interests exist.