
Abstract
Abstract
Comprising a large number of species, the genus Astyanax has been intensively studied by several approaches to elucidate its evolutionary relationships. Such studies have demonstrated that many nominal species are artificial clusters where distinct taxa are grouped under the same denomination. Astyanax aff. fasciatus stands out due to its high karyotypic diversity, since cytogenetic studies have reported three standard cytotypes (2n = 46, 48, and 50), as well as cases of sympatry between cytotypes, variant cytotypes, and B chromosomes. In this study, we attempted to evaluate the reliability of the chromosomal differences in relation to the analysis of the ATPase6/8 mitochondrial DNA (mtDNA) sequence, thereby providing subsidies to the evolutionary reconstruction of this group. Nine populations from four distinct hydrographic basins along Southeastern Brazil were analyzed. These are the first cytogenetic data collected for four of them. Fluorescent in situ hybridization with 5S rDNA probe evidenced the presence of a standard phenotype for the group and the existence of a new arrangement in the individuals from Ribeira de Iguape River. Besides the karyotypic variation, the genetic distance was low among the studied populations and some aspects of the evolutionary relationships among distinct cytotypes/populations could be ascertained by phylogeographic studies. The incipient molecular structuring of certain cytotypes in different hydrographic basins indicates the role of different evolutionary processes on the diversification of the group.
Introduction
F
Cytogenetic studies allowed the identification of cryptic species in distinct fish groups, particularly in Astyanax,2–4 showing the great plasticity of the group, which seems to be followed by molecular diversity. 5
The species of Astyanax are known as tetras and correspond to small fishes distributed throughout Central and South Americas. The Astyanax fasciatus group inhabits several hydrographic systems and its type locality is referred to as “Rivers from Brazil,” which makes it difficult to determine the characteristics of the species type. The wide distribution and biological preferences of the species can intensify evolutionary processes on its diversification. Several species of the genus were previously considered a subspecies of A. fasciatus. One of them, Astyanax mexicanus, popularly known in the aquarium trade as blind tetra, is one of the most important model for the studies of developmental biology.
Regarding cytogenetic data, three standard cytotypes are identified in A. aff. fasciatus according to the chromosomal pairing (2n = 46, 48, and 50), with slight karyotypic differences within each cytotype. The cytotype with 2n = 46 chromosomes is predominant in the Upper Paraná River basin, mainly in the Paranaíba River basin, 6 Middle Tietê River,7,8 Paranapanema River basin, 9 and Mogi-Guaçu River. 3 The cytotype with 2n = 48 chromosomes is found in the Grande River 10 and Mogi-Guaçu River,3,9 which belong to the Upper Paraná River basin, as well as in the Tibagi River (a tributary of Paranapanema River), 11 in the coastal drainages along the Ribeira do Iguape River basin 12 and Northeastern Brazil.13,14 On the other hand, the cytotype with 2n = 50 chromosomes was detected only in the Tibagi River. 11 Besides the distinct standard cytotypes, other variant forms, not related to B chromosomes, have already been reported in A. aff. fasciatus, bearing 2n = 45, 46, 47, 3 and 49 chromosomes. 11
Studies based on chromosomal and morphometric data have shown some degree of structure among different populations/cytotypes,11,13,15 although molecular markers (RAPD and ISSR) have indicated a recent differentiation and likely introgression, particularly within specimens from the Mogi-Guaçu River. 16 The approach to associate both (chromosomal and molecular data) is rare for Neotropical fishes and absent for A. aff. fasciatus. The aim of this study is to test if the chromosomal evolution in A. fasciatus is consistent with molecular evolution and biogeography and if the different major cytotypes could represent different species.
Materials and Methods
The analyzed specimens identified as A. aff. fasciatus were collected in nine different sites along several hydrographic basins (Fig. 1 and Table 1). After the cytogenetic procedures and tissue sampling for DNA extraction, the individuals were fixed in 10% formaldehyde and stored in 70% ethanol. They were identified by specialists and deposited in the fish collection of the Universidade Federal do Rio Grande do Sul (UFRGS, Brazil).
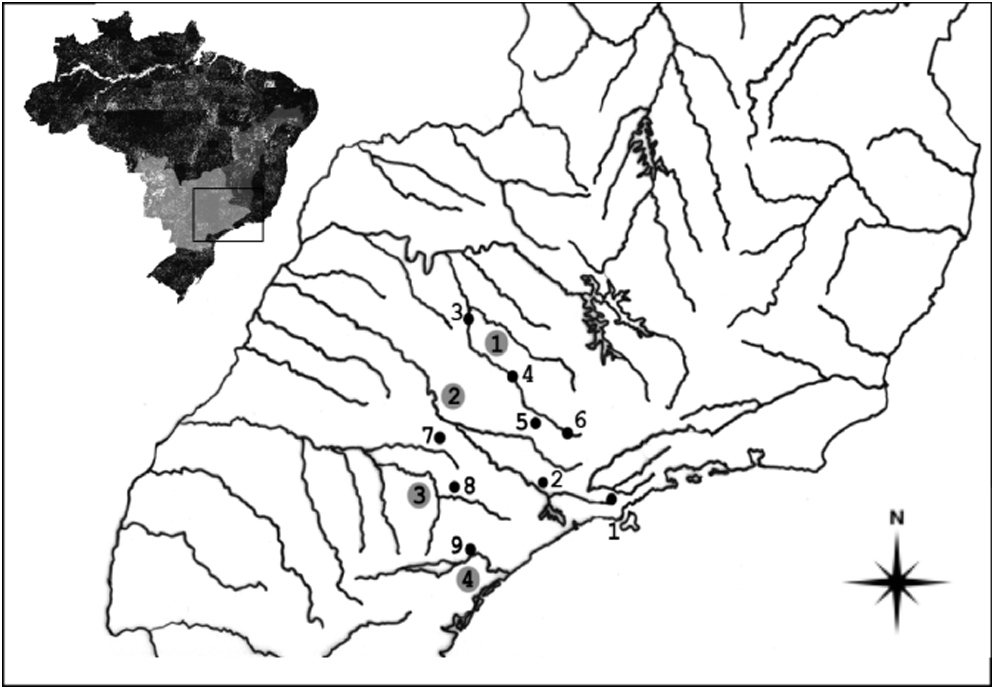
Map showing the populations of Astyanax aff. fasciatus under study. The numbers in the circles indicate the main hydrographic basins in Southeastern Brazil: (1) Mogi-Guaçu River; (2) Tietê River; (3) Paranapanema River; and (4) Ribeira de Iguape River. The numbers without circles indicate the collection sites: (1) Salesópolis/SP; (2) Indaiatuba/SP; (3) Barrinha/SP; (4) Cachoeira de Emas/SP; (5) Araras/SP; (6) Ouro Fino/MG; (7) Angatuba/SP; (8) Pilar do Sul/SP; and (9) Sete Barras/SP.
Cytogenetic data from this work.
Previously analyzed populations by: Pazza et al. 15
Lack of sequencing data.
Previously analyzed populations by: Pazza et al. 3
Chromosomal studies
Mitotic chromosomes were obtained according to Gold et al. 17 Silver nitrate staining (Ag-NOR) was performed according to Kavalco and Pazza. 18 The direct localization of ribosomal cistrons was accomplished by fluorescent in situ hybridization (FISH)3,19 using 18S rDNA 20 and 5S rDNA 21 probes. The probes were marked with Biotinylated Uridin (BdUTP) using the Nick Translation Bionik Labeling System Kit, Invitrogen®. In the FISH procedure, high stringency washes were preferred, using 20% Formamide in 0.1× SSC. The signals were amplified using a FITC-avidin and anti-avidin conjugated with biotin solution and incubated in the NFDM buffer. The slides were set up with 25 μL of Vectashield Mounting Medium antifade, Vector® with Propidium Iodide (1.5 μg/mL).
The chromosomal preparations were analyzed using an optical microscope and captured (resolution of 5 Mp) using the image system CoolSnap Pro and the software Image Pro Plus (Media Cybernetics). The chromosomal types were classified according to the arm ratio (AR) proposed by Levan et al., 22 as follows: M = metacentric (AR = 1.00–1.70), SM = submetacentric (AR = 1.71–3.00), ST = subtelocentric (AR = 3.01–7.00), and A = acrocentric (AR >7.00).
Sequencing and analysis of mtDNA
The DNA extraction was accomplished using the Wizard kit (Promega), following the manufacturer's instructions. The DNA samples were quantified in agarose gel using a Ladder Low Mass (Invitrogen) and diluted to a working solution of 10 ng/μL.
Five representatives from each collection site were used, when possible. One mitochondrial segment corresponding to the subunits 6 and 8 of the ATPase gene was chosen. The following primers were used in polymerase chain reaction (PCR) experiments to amplify a segment of nearly 910 bp: ATP8.2-L8331 (5′-AAAGCRTTRGCCTTTTAAAGC-3′) and CO3.2-H9236 (5′-GTTAGTGGTCAGGGCTTGGRTC-3′). 23 This segment of mtDNA was chosen because it was used in previous studies, proving useful to distinguish groups in the complex species Astyanax aff. bimaculatus. 24
The amplification reactions were carried out in a thermocycler Mastercycler Gradient Eppendorf, totalizing a volume of 25 μL, containing 15 ng of template DNA, Tris-KCl (20 mM Tris-HCl pH 8.4 and 50 mM KCl), 1.5 mM MgCl2, 2.5 μM of each primer, 0.1 mM of each dNTP, and 2.5 U Taq-polymerase (Invitrogen). The PCR conditions comprised a first denaturation step at 94°C for 4 min, hybridization at 56°C for 30 s, and extension at 72°C for 2 min, followed by 40 cycles of 15 s at 94°C, 30 s at 56°C, and 2 min at 72°C, and a final extension step for 10 min at 72°C. After checking the amplification results in 1% agarose gel, the PCR products were purified using the ChargeSwitch® PCR Clean-Up kit (Invitrogen). This DNA was used as a template in the sequencing reactions, using the BigDye™ Terminator Cycle Sequencing kit v. 3.1 (Applied Biosystems), according to the manufacturer's instructions. About 10 ng of template DNA and 5 pmol of each primer were added in each sequencing reaction. The samples were submitted to 25 cycles of 30 s at 96°C, 15 s at 50°C, and 4 min at 60°C, and precipitated for further sequencing.
The sequences were visualized and edited using the software Chromas Lite v2.01 and compared with the GenBank database (www.ncbi.nlm.nih.gov) using BLASTn to confirm their identity by comparison with the mitochondrial sequences of other fish species. Next, the DNA sequences were aligned based on pairwise and multiple alignments, with the following penalties: gap opening = 20 and gap extension = 6.66, using the algorithm ClustalW v1.6 25 available in the software Mega v5. 26
The phylogram of maximum likelihood (ML) was performed based on the best evolutionary model that explains the data, HKY+G, according to the test of the evolutionary model performed in MEGA v526 software. The statistical data were obtained using the software packages DAMBE v5.3.19, DnaSP v5, 27 and MEGA v5. 28
Results
Chromosomal analyses
The population sampled in Salesópolis, Upper Tietê River basin, presented four distinct cytotypes: 2n = 46, 12 M+20 SM +10 ST +4A, fundamental number (FN) = 88 (Fig. 2a); 2n = 48, 8 M+22 SM +12 ST +6 A, FN = 90 (Fig. 2b); 2n = 49, identical to the cytotype with 2n = 48 plus a middle-sized SM B chromosome in all metaphase spreads, FN = 92 (Fig. 2c); and 2n = 50, 8 M+16 SM +14 ST +12 A, FN = 88 (Fig. 2d), which were identified as a new species. The population from Sete Barras, Ribeira de Iguape River basin, presented a karyotype with 2n = 48 chromosomes, composed of 10 M+20 SM +12 ST +6A, FN = 90 (Fig. 2e). The populations from Indaiatuba (Middle Tietê River basin) and Pilar do Sul (Paranapanema River basin) presented a karyotype with 2n = 46 chromosomes, so that 12 M+20 SM +10 ST +4A, FN = 88 (Fig. 2f, g, respectively). Besides the same karyotypic formulae, cytotypes with 2n = 46 chromosomes presented slight differences in relation to the size of the second acrocentric chromosomal pair; they were similar to the first acrocentric pair from Salesópolis individuals and remarkably smaller in the populations from Indaiatuba and Pilar do Sul.
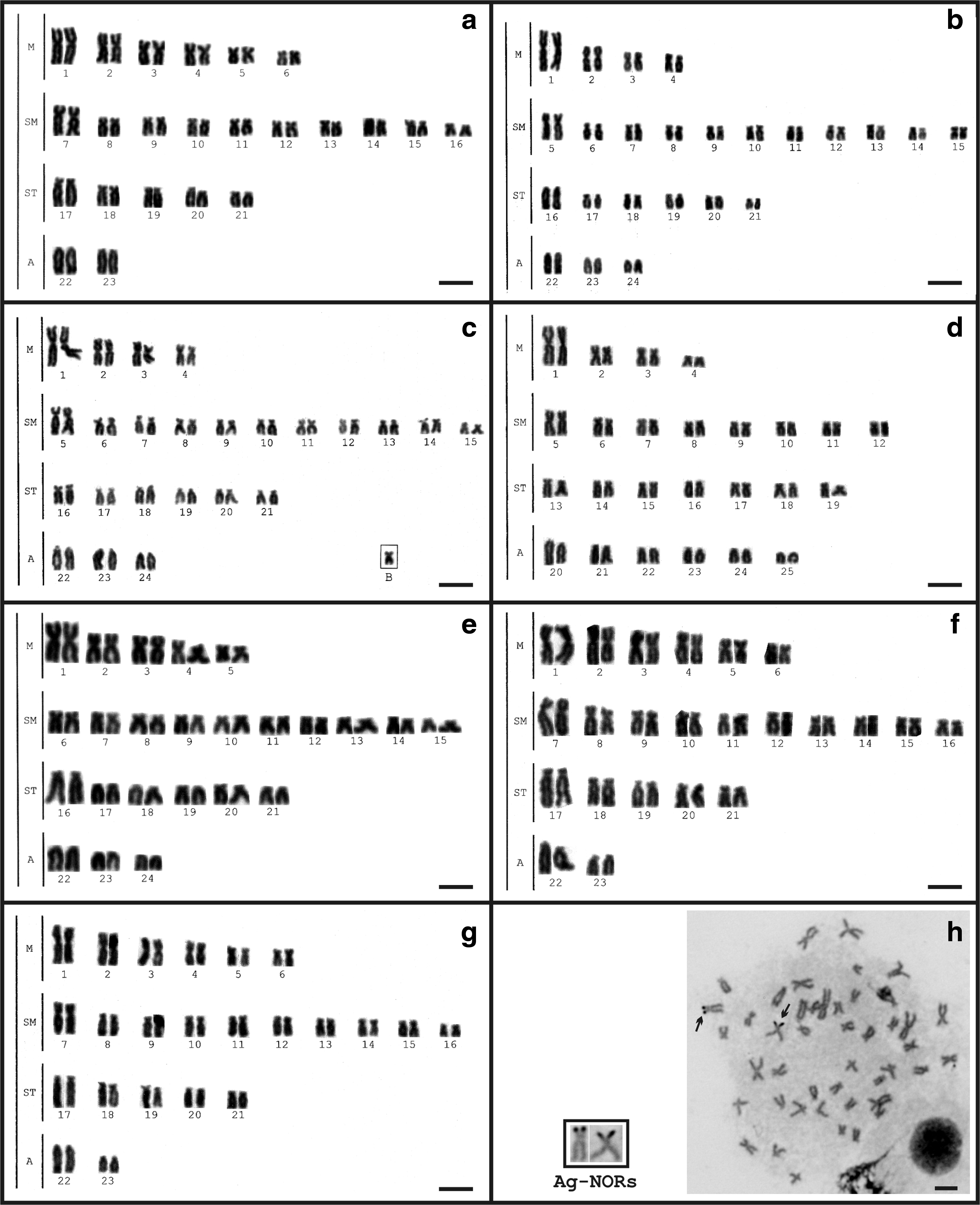
Karyotypes of A. aff. fasciatus after Giemsa and Ag-NOR staining from Salesópolis/SP-Upper Tietê River basin [2n = 46
Multiple Ag-NORs were detected by silver nitrate staining, although marks on short arms of a subtelocentric pair were more frequent in all analyzed cytotypes (Fig. 2h).
Two up to seven chromosomes bearing terminal signals were observed after FISH with 18S rDNA probes, ranging from conspicuous blocks in some populations to minute marks, as observed in Sete Barras sample (Fig. 3a–g). After 5S rDNA FISH, two chromosomal pairs were marked, one metacentric pair carrying pericentromeric fluorescent signals and one acrocentric pair with pericentromeric marks. The only exception was detected in the population from Sete Barras, which presented a site at a pericentromeric region in a pair of acrocentric chromosomes, besides the same four chromosomes bearing 5S rDNA also observed in the other populations.

Metaphases of A. aff. fasciatus showing the 18S
Molecular studies
Out of the 801 bp analyzed in the sequenced fragment, 47 were variable. The following nucleotide frequencies were found: 0.298 (A); 0.294 (T/U); 0.279 (C); and 0.129 (G). Transition/transversion deviation was R = 4.54. The evolutionary distances within and among different populations and cytotype groups (p-distance) are shown in Table 2.
Differences are based on 801 bp of the mitochondrial DNA (mtDNA) ATPase6/8 region. The analysis involved 28 nucleotide sequences. Codon positions included were first+second+third+Noncoding. All positions containing gaps and missing data were eliminated.
ML phylogram allows the identification of six major groups based on their hydrographic basin (Fig. 4). Salesópolis individual with 2n = 50 chromosomes appears as a distant outgroup in the ML tree based on the best evolutionary model with bootstrap (500 replicates), which indicates that it must pertain to a different species or species group. Posterior extensive analyses about its morphology were performed, and these specimens were considered to be a new species that belongs to the Astyanax aff. scabripinnis cryptic group. However, this sample was maintained in the analysis due to the convergent chromosomal features presented, similar to karyomorphs described in other populations from Southeastern Brazil. Therefore, we decided not to include it in the molecular analysis, aiming to avoid long branch attraction.
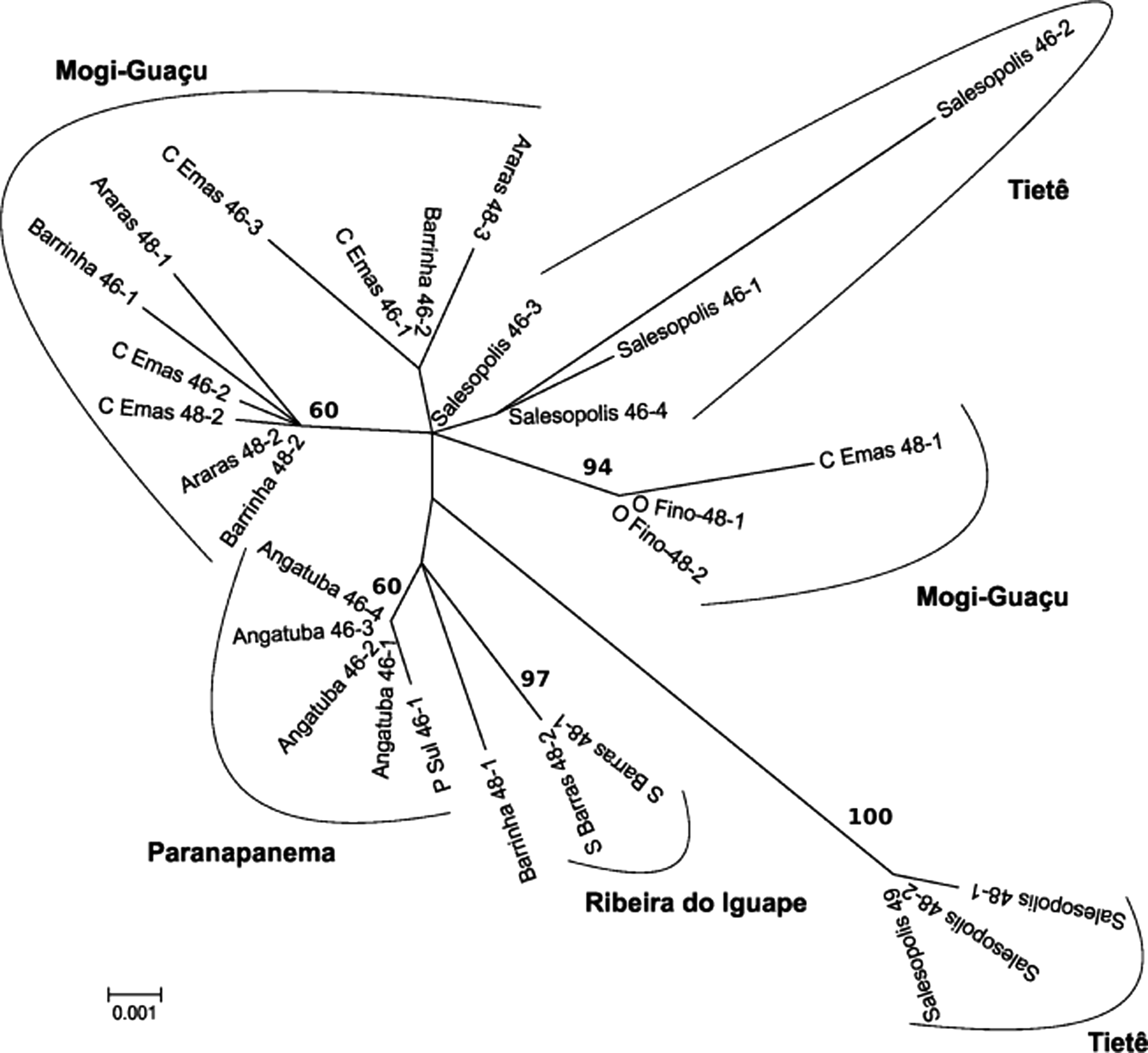
Molecular Phylogenetic analysis by maximum likelihood method. The evolutionary history was inferred by using the maximum likelihood method based on the Hasegawa-Kishino-Yano model. The tree with the highest log likelihood (−1446.0844) is shown. Initial tree(s) for the heuristic search were obtained automatically by applying Neighbor-Join and BioNJ algorithms to a matrix of pairwise distances estimated by the Maximum Composite Likelihood approach. Then, the topology with superior log likelihood value was selected. A discrete Gamma distribution was used to model evolutionary rate differences among sites (five categories [+G, parameter = 0.2128]). The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. The analysis involved 28 nucleotide sequences. The following Codon positions were included: first+second+third+Noncoding. Bootstrap values from 500 replicates (values <60 is not shown). All positions containing gaps and missing data were eliminated. There were a total of 801 positions in the final dataset.
Discussion
Our results show that in A. fasciatus, the chromosomal evolution is faster than molecular evolution and it is not consistent with the phylogeographic distribution of the species, which is different to that which was observed in the A. bimaculatus—Astyanax altiparanae group. 24
Several approaches have been used to elucidate the extension of the group A. fasciatus, recognized after a review by Eigenmann. 28 Based on taxonomic revisions, some subspecies have been reanalyzed and reached a species status, such as Astyanax parahybae 29 and Astyanax henseli. 30 In the same way, some authors have identified the mistakes in the identifications in Astyanax, 31 increasing the number of possible new species of the genus.
There has been much debate about the proper methods to define a species, based on different “species concepts,” 32 as well as concerns about the increasing number of species and their effect on conservationist practices. Most conservation programs rely on species richness, endemism, diversity, endangered status, etc., for comparison between distinct sites or taxa. 33 In this study, we attempt to use the information obtained from analysis of mtDNA and chromosomes to explain the natural history of the A. aff. fasciatus fish group.
The occurrence of sympatric cytotypes in A. aff. fasciatus has been previously reported, particularly in Cachoeira de Emas, on the Mogi-Guaçu River, where both standard 2n = 46 and 48 cytotypes are found in sympatry along with other five variant cytotypes with diploid numbers ranging from 45 to 47. 3 In Tibagi River, sympatric standard cytotypes with 2n = 48 and 50 and a putative hybrid (2n = 49) were also detected. 11 Three cytotypes, 2n = 46, 48, and 50, were also observed in the Tietê River basin. 34 In this work, three sympatric cytotypes of A. aff. fasciatus were detected along the Tietê River, close to its headwaters in Salesópolis (SP): the standard cytotypes with 2n = 46 and 48 chromosomes and a variant with 49 chromosomes, apparently derived from a standard 2n = 48 karyotype plus a B chromosome (Fig. 2a–c). B chromosomes have already been reported in another population of A. aff. fasciatus, and they are commonly found in other Astyanax species, 35 particularly in A. aff. scabripinnis. 36
Despite some karyotypic differences among these three sympatric cytotypes in Salesópolis (Tietê River), they share remarkable common features, such as the distribution of 5S and 18S ribosomal genes (Fig. 3a–c). These features are also shared with a fourth karyomorph found in Salesópolis and preliminary identified as A. aff. fasciatus, with 2n = 50 chromosomes (Fig. 2d).
The priori analysis indicates great proximity among all sampled individuals from this collection site, due to their morphology as well as the physical mapping of the rDNA genes (Fig. 3). Similarly, the distribution of heterochromatin and satDNA As-51 is quite similar among the individuals with 2n = 46, 48, and 50 chromosomes. 37 However, the analysis of the ATPase gene showed great divergence in the haplotypes of individuals with 2n = 50 chromosomes, and an additional morphological analysis was performed. This approach allowed the recognition of a new Astyanax species of the A. scabripinnis group. Indeed, the sequences of these individuals seem to be closely related to the Astyanax paranae, a species derived from the A. scabripinnis group and sampled in the Paranaíba River—Upper Paraná River basin (data not shown). The association of molecular and cytogenetic analysis could determine the real status of the sympatry of Astyanax genus at the Salesópolis region (Upper Tietê River basin), which highlights the importance of multidisciplinary approaches in evolutionary biology studies.
Notoriously, another population from the Tietê River basin (Indaiatuba/SP) presents a karyotype with 2n = 46 chromosomes and a karyotype formula (Fig. 2g) similar to that verified in Salesópolis. However, a remarkable size difference in two acrocentric chromosomal pairs can be observed. The specimens from Salesópolis (Fig. 2a) have these chromosome pairs with similar size, whereas in Indaiatuba, the 23rd pair is about 50% smaller than the 22nd pair (Fig. 2g). This feature, found in individuals from Indaiatuba, is shared with other populations bearing 2n = 46 chromosomes, such as those from Pilar do Sul/SP (Fig. 2f). It had been previously reported in populations from the Tietê,7,8 Mogi-Guaçu, 3 and Paranapanema 15 River basins. Another common characteristic of such populations is a dispersed distribution of the As-51 satellite DNA over several chromosomes,9,37 while the population from Salesópolis presents the same satellite DNA in a single pair. 37 These results suggest that such differences reveal more than just interpopulation divergence, but rather independent taxonomic units.
Among the cytotypes with 2n = 48 chromosomes, found in different hydrographic basins, the most divergent one refers to the population from Sete Barras (Ribeira de Iguape) (Fig. 2e), characterized by a very differentiated karyotypic formula in relation to those observed in the Upper Paraná River3,9 (this work) and in the coastal populations from Northeastern Brazil.13,14 In addition, although the chromosomal number remains the same, the karyotype structure can be distinctive, as observed by Morelli et al. 12 in another population from Ribeira de Iguape River basin. The karyotypes and other cytogenetical features of A. aff. fasciatus from Ribeira de Iguape River basin suggest a vicariance event in this hydrographic system, probably as result of a founder effect. This population also shows an incipient molecular structuring (Fig. 4) and can be considered an independent evolutionary unit.
Besides the divergence in the karyotypic macrostructure, the distribution of 5S rDNA genes is also noteworthy. The presence of four 5S rDNA-bearing chromosomes with pericentromeric marks on a metacentric pair is the most frequent pattern reported within Astyanax,24,38,39 just as observed in the populations from Tietê and Paranapanema Rivers (Fig. 3h). On the other hand, the population from Sete Barras (Ribeira do Iguape basin) presented six chromosomes bearing 5S rDNA sites (Fig. 3i), including the marker metacentric pair, a feature that has been previously reported in populations from coastal northeastern rivers of A. aff. fasciatus 14 and A. mexicanus from North America. 40 It is important to reinforce the existence of a marker chromosome bearing 5S rDNA site in the Astyanax species from the continental drainages, while in the species spread exclusively on the coastal rivers, this marker is absent and the amount of chromosomes bearing 5S ribosomal genes is greater. 24 This feature is a symplesiomorphy of the species of Astyanax from the continental drainages, and it seems to be a significant marker for chromosomal studies of the group. Probably due to the dynamics of chromosomal rearrangement in the group, Medrado et al. 14 observed a lack of this marker in one population from northeastern Brazil, representing an autapomorphy.
The distribution of 18S rDNA sites followed a common trend for Astyanax, which is location at terminal sites of several chromosomes3,38,41 and high occurrence of a single active Ag-NOR bearing pair. Indeed, studies with silver nitrate staining have shown a pattern of preferential Nucleolar Organizing Region (NOR) activation in these fish.2,40 The population from Sete Barras (Ribeira de Iguape River basin) presented a more divergent distribution of 18S ribosomal genes, comprising six minor discrete sites (Fig. 3e).
Despite the wide karyotypic variation observed in this work and in other reports, the mitochondrial ATPase6/8 gene sequence was poorly variable, which suggests a recent divergence in this fish group (Table 2). Nonetheless, some interesting clusters can be noted. Both Salesópolis (Upper Tietê River) and Mogi-Guaçu River populations were subdivided in ML phylogram (Fig. 4). In Salesópolis, the cytotypes with 2n = 46 and 48 are properly structured; meanwhile, the individual with 2n = 50 chromosomes was so distant that it must be a different species or group of species. Cytotype 2n = 48 from Salesópolis shows the highest genetic distances compared with any other cytotypes or populations (Table 2). As for Mogi-Guaçu River fishes, three main groups were formed in the ML phylogram (Fig. 4). The first one (Mogi-Guaçu) is composed of specimens with 2n = 48 from Emas waterfalls and the individuals from Ouro Fino (2n = 48). The second and wider group comprises specimens with 2n = 48 and 46 chromosomes, but with a point of divergence that splits them into two groups where one is formed primarily of individuals with 2n = 46 chromosomes and the other is formed primarily of individuals with 2n = 48 chromosomes. However, the presence of individuals with different karyotypes suggests an incipient divergence in the group, perhaps with some degree of interbreeding, at least in the remote past. These data are similar to those obtained by RAPD and ISSR molecular markers, 16 although the nuclear markers have provided high values of genetic distance among the populations/cytotypes. Pazza et al. 3 suggested that the cytotype with 2n = 46 chromosomes found in Mogi-Guaçu River is an invader, formerly from the Tietê River basin. This hypothesis was reinforced by the genetic distance between both cytotypes as estimated by molecular markers, although the gene flow is still present, 16 which might account for the variant cytotypes and for the haplotype sharing demonstrated by both RFLP 42 and mtDNA sequencing. These data can suggest that the invasion by the 2n = 46 cytotype from the Tietê River basin is responsible for the admixture crossings and gene flow seen in the distinct cytotypes.
The population structure found in Angatuba (Paranapanema), particularly in relation to the specimens from Araras, is in agreement with the results obtained by the analysis of chromosomal and morphometric traits. 9 Morphometry seems to be a good tool for studying different populations of Astyanax.2,9,11,43 Such structure occurs despite their small genetic distance (Table 2) and similarity with the cytogenetic data in the standard 2n = 46 cytotype observed in the Emas waterfalls. 3
Despite the remarkable divergence in either the karyotype structure or the distribution of 5S rRNA genes (Fig. 3), both the Sete Barras (Ribeira de Iguape River basin) and Angatuba (Paranapanema River basin) samples presented low genetic distance (Table 2). These collection sites present considerable proximity despite the presence of an important water divisor in the region, the Paranapiacaba ridge.
Sequences of mtDNA have been used to infer phylogeographic patterns in several fish species. 23 Using mtDNA, Völker et al. 44 reported a low haplotype differentiation on phylogeographic analyses associated with high karyotypic differentiation in Chromaphyosemion bivittatum (Cyprinodontiformes, Nothobranchiidae), similar to that observed among the populations of A. fasciatus. However, the comparative approach between phylogeography and molecular cytogenetics seems to be resolutive for other species of the genus Astyanax, and the study of gene regions with a greater accumulation of evolutionary changes can generate better results in the future.
Matoso et al. 45 found indications of a possible introgression event between A. fasciatus and A. altiparanae at the Paraná River basin. They found that hybridization between different and structured Astyanax species could be a common phenomenon since natural populations show it. This feature has also been used in crossbreeding between close species, such as the epigean and cave forms of A. mexicanus. The phylogeography of the group seems to be punctuated by natural introgression events. 46
These and previously reported karyotypic data, coupled with evidence of higher genetic distance among distinct cytotypes based on nuclear markers, suggest that the rate of chromosomal mutation is higher in this group than the mitochondrial variation. This fact may be related to introgressions caused by secondary contact that minimize the differences in the mitochondrial genome once it lacks recombination and presents maternal inheritance. A better explanation is perhaps that the intrinsic genomic features in these species may lead to karyotypic evolution.
Footnotes
Acknowledgments
The authors are thankful to Dr. Luiz R. Malabarba and Dr. Vinicius A. Bertaco for the taxonomical identification, Carlos E. Lopes for assistance in collecting the samples, and the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP), and Universidade Federal de Viçosa for financial support.
Disclosure Statement
No competing financial interests exist.