
Abstract
Abstract
The taxonomy of Satanoperca spp. is still unresolved, especially because coloring, one of the main diagnostic characters, is variable among species of this genus. Thus, the aim of this study was to elucidate the relationship between the genome and the organization of the chromosome in two Satanoperca species. Our main goal was to develop a method to better differentiate taxa and understand the evolution of Satanoperca jurupari and Satanoperca lilith karyotypes, which we analyzed with classical and molecular cytogenetics. Both species have the same diploid number (2n) of 48 and location of 5S rDNA sites on pair 5. Nonetheless, the distribution of heterochromatin and 18S rDNA sites followed a species-specific pattern. The interstitial telomeric sites were not highlighted in either species. Regardless, a single B chromosome was identified in some metaphases of S. lilith. These data show that Satanoperca species harbor chromosomal features that can be used to identify the two species of Satanoperca studied here, allowing for the use of cytogenetic markers to make taxonomic inferences within the genus.
Introduction
C
The subfamily Geophaginae belongs to Cichlidae and is a problematic clade.8–11 Molecular phylogenetic analyses shows Geophagus is a polyphyletic group closely related to Mikrogeophagus (Geophagus brasiliensis) and Gymnogeophagus (Geophagus stricto sensu and Geophagus steindachneri). On the other hand, Satanoperca is more closely related to Apistogramma and Taeniocara in the “Satanoperca clade.” Satanoperca includes a group of species widely distributed throughout South America, with eight that are currently valid and three additional ones in the process of being described. The valid species include: S. daemon, 12 S. jurupari, 12 S. acuticeps, 12 S. papaterra, 12 S. leucosticte, 13 S. mapiritensis, 14 S. lilith, 15 and S. rhynchitis. 16 The additional three undescribed Satanoperca spp. include one found in the Araguaia–Tocantins river basin, another in the Madeira and Jamari rivers, and the final one located in the upper and middle Negro river. 17
Despite the recent review of this genus, the taxonomy of Satanoperca remains unresolved because morphological characters are often used to delineate most species. This is problematic because one of the main characters used is coloring, which is variable within the genus. Thus, studies aimed at establishing links between the genome and chromosomal organization can potentially improve the differentiation of taxa and aid in understanding karyotype evolution among groups. The focus of the present study was to cytogenetically characterize two species of Satanoperca (S. jurupari and S. lilith) found in the Central Amazon region using classical and molecular techniques.
Materials and Methods
Twelve S. jurupari (8 males and 4 females) individuals were collected from Catalão Lake (3°09′25″S, 59°54′49″W), located at the confluence of the Negro and Solimões rivers. In addition, five S. lilith (3 males and 2 females) specimens were collected from the Anavilhanas archipelago (2°23′41″S, 60°55′14″W), located in the Negro river. All samples were collected with the permanent license from the Brazilian Institute of Environment and Renewable Natural Resources (IBAMA) (permanent license No. 22984-1/2010). Fish were anesthetized to death with clove oil diluted in water, and subsequently numbered, recorded, fixed in formaldehyde, stored in alcohol, and deposited in the fish collection of the Instituto Nacional de Pesquisas da Amazônia (INPA), Manaus, Amazon, Brazil (voucher numbers INPA-ICT 048816 and INPA -ICT 048817).
Mitotic chromosomes were obtained from kidney cells, 24 h after application of 1 mL of an aqueous solution of yeast and 0.0125% colchicine per 100 g live weight for 45 min. 18 Nucleolus organizer regions (NORs) were identified after impregnation with silver nitrate 19 and detection of the distribution of heterochromatin (C-band) using barium hydroxide. 20 Genomic DNA was extracted from the muscle tissues of the two Satanoperca species following the protocol of Sambrook and Russell 21 and then used for amplification of the 5S rDNA and 18S rDNA probes, with the polymerase chain reaction 22 using the following primers: 5Sa (5′-TACGCCCGATCTCGTCCGATC-3′) and 5Sb (5′-CAGGCTGGTATGGCCGTAAGC-3′) 23 as well as 18Sf (5′-CCGCTTTGGTGACTCTTGAT-3′) and 18Sr (5′-CCGAGGACCTCACTAAACCA-3′). 24
Telomeric sequence amplification was performed without DNA using the following primers: (TTAGGG)5 and (CCCTAA)5. 25 Isolated probes were labeled with the nick translation method using biotin 14-dATP (BioNick Labeling System; Invitrogen) and/or digoxigenin-11-dUTP (Dig-Nick Translation Mix; Roche). The Avidin-FITC antibody (Sigma-Aldrich), anti-Avidin–Biotin (Sigma-Aldrich), and anti-digoxigenin rhodamine (Roche) were used for detection and signal amplification. After analysis and numeration of chromosomes with a light microscope, the modal diploid number and relative frequencies for each individual were determined. The best representatives of metaphase obtained with classical techniques (conventional, NORs, and C band) were photographed using an oil immersion objective and the Canon Power Shot A 650 IS digital camera. Cells in metaphase were subjected to molecular techniques (FISH) and epifluorescence was assessed after images were captured using an Olympus Bx-51 light microscope and an oil immersion objective. Adobe Photoshop 7.0 software was used for karyotype analysis (version CS4) of chromosomes in mitotic metaphase, which were tentatively cut and annealed.
Chromosome morphology was determined according to the arm ratio criteria (RB = BM/Bm, where BM and Bm are the bigger and lower arms, respectively), according to Levan et al. 26 Owing to the peculiar difficulty of chromosome pairing in the cichlid, since the chromosomes are similar in size and morphology, chromosomes were classified into meta-acrocentric, subtelocentric, or submetacentric according to a previous study by Thompson. 4 B chromosomes were easily identified due to their distinct morphology from the chromosomes of the A complement and reduced size. To establish the fundamental number (FN), the first individuals were considered to have two chromosome arms, and the last carriers, only one arm.
Results
Both S. jurupari and S. lilith had diploid numbers of chromosomes equal to 48 (4 m-sm + 44 st-a), and the FN was equal to 52 (Fig. 1a, d). Nevertheless, B chromosome was present in some metaphase of all males and females of S. lilith. Males and females of both species had the same karyotypes. Constitutive heterochromatin is distributed in the pericentromeric regions of most chromosomes of both species, however, the markings in S. jurupari are conspicuous and those in S. lilith are tenuous. Furthermore, markings on the terminal region of the long arms of the homologous from pair 15 were observed in S. jurupari and the B chromosome found in S. lilith was completely heterochromatic (Fig. 1b, e).
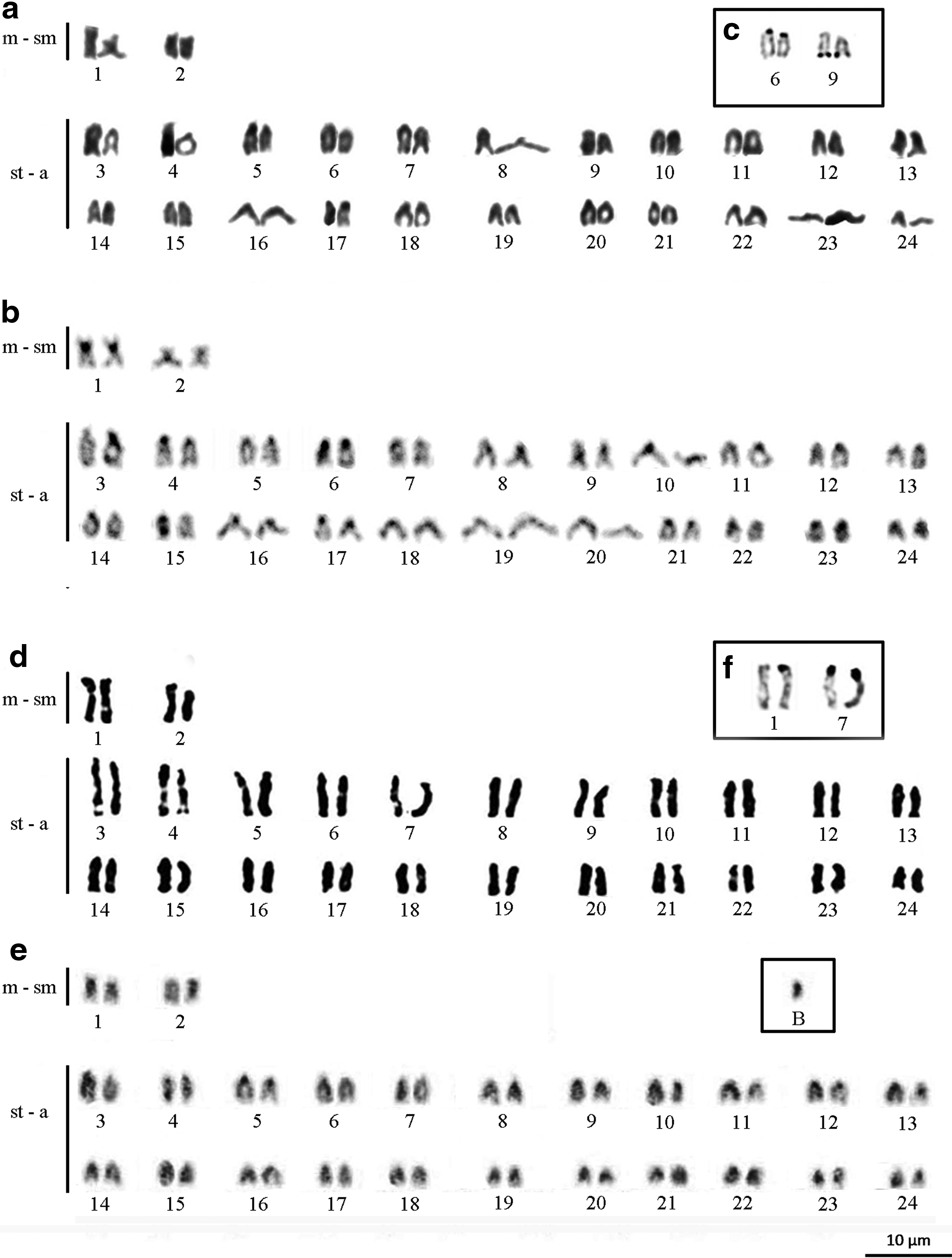
Karyotypes of Satanoperca jurupari
Multiple NORs were identified by Silver Nitrate staining. These NORs were located on the short arms of both homologous chromosomes from pair 6 (st-a), in both end regions of the long arm from the chromosome pair 6 (st-a) in S. jurupari (Fig. 1c), and in the terminal region of the long arms of both homologous pairs 1 and 7 in S. lilith (Fig. 1f). The 18S ribosomal DNA sites were also mapped in these chromosomes (Fig. 2c, f), whereas the 5S ribosomal DNA sites were located, interstitially, on the homologues from pair 5 in S. jurupari and S. lilith (Fig. 2b, e). Telomeric sites were evident only in the terminal regions of the chromosomes in both species (Fig. 2a, d).

Karyotypes of S. jurupari and S. lilith showing an absence of interstitial telomeric sites in both species
Discussion
Karyotypic reports in Satanoperca are available for only two species (S. papaterra from Paraná River basin, Southeastern Brazil, and S. jurupari from Araguaia River Basin, Central Brazil). Both these species were shown to have a diploid number of chromosomes equal to 48 and to lack differentiated sex chromosomes, but harbor distinct karyotypes.27–29 In this study, both S. jurupari and S. lilith, from the Amazon Basin, showed the same diploid number described for most Neotropical cichlids (48 chromosomes) and similar chromosomal formulas, corroborating previous evidence for the conservation of chromosomal macrostructure, which is considered to be a baseline characteristic among Neotropical cichlids.28–31
The presence of a B chromosome has been reported in S. jurupari from the Araguaia River, Mato Grosso, Brazil. 32 However, in this study, this extra chromosome was not observed in individuals belonging to S. jurupari, although the karyotype formula was similar.28,29 On the other hand, in this study, B chromosomes were observed in S. lilith.
Within the family Cichlidae, B chromosomes were previously identified in 6 Neotropical species and 13 African species,33–35 raising some concerns related to its origin, evolution, and function. Recent studies have provided insights into these concerns within the cichlids.33,36 In Astatotilapia latifasciata, an African species from Nawampasa Lake, repetitive DNA sequences were found in B chromosomes,37,38 which were composed of fragmented sequences and transcriptionally active genes, suggesting that the origin of these chromosomes is involved with the replication and amplification of sequences, subsequently forming isochromosomes and leading to the introgression of genome segments by transposable elements (TEs).33,36,39
However, in populations of Lithocromis rubripinnis, another African cichlid in Lake Victoria, evidence for B chromosomes was found only in females, suggesting that sex determination is directly linked to the presence of B chromosomes within this African group.35,39,40 However, in the present study, the presence of a B chromosome in S. lilith was observed in both males and females, indicating that this chromosome is not associated with the sex in this species.
Furthermore, Pires et al. 34 analyzed the mitotic and meiotic behavior of B chromosomes in populations of Crenicichla lepidota, from Guaiba Lake, (RS). This lake is located close to the industrial centers of Rio Grande do Sul, which is a city known to receive domestic and industrial waste, and found that these chromosomes were present in males and females. However, as few species of Neotropical cichlids harbor B chromosomes, the authors suggested that the origin of these chromosomes in the family is recent, and is possibly attributed to exposure to certain environmental conditions. A probable origin was reported for three species of Amazonian cichlids by Feldberg et al., 41 who suggested that the B chromosomes originated due to chromosomal breaks caused by the bioaccumulation of heavy metals in the natural environment in Amazon. Thus, the naturally present mercury in the Amazon possibly acts as an environmental stress and leads to the creation of B chromosomes in S. lilith.
The two species studied here had distinct heterochromatic patterns, where S. jurupari presented markings predominantly in the pericentromeric region with blocks in the telomeric region from pair 15. The tenuous markings of S. lilith were located in the pericentromeric region. However, distribution patterns of heterochromatin were preserved within species, with markings in the pericentromeric regions of almost all chromosomes reported for most Neotropical cichlids.3,29,30,42–45
Mapping the 5S rDNA site revealed it was located in the interstitial region of pair 5 in both species. The location of these sites in the interstitial region has been observed in most Neotropical cichlids, as well as other fish groups,45–48 which is considered to be a characteristic related to the ancestral organization of this gene in the genome.48–51 Furthermore, the interstitial location of these genes on cichlid chromosomes may be a preventative measure, inhibiting gene dispersion events from occurring throughout the genome.48,52 Additionally, this could indicate that this default location on the chromosome could correspond to the ancestral version in this genus.
However, the presence of chromosomal rearrangements in Satanoperca is evident when we compare the number and position of 18S rDNA sites. In particular, S. jurupari from the Araguaia River (MT) has 18S sites in only a few acrocentric chromosomes, 25 whereas both Amazonian species have two pairs of chromosomes carrying these sites. This discrepancy may be related to the dynamic evolution of these ribosomal genes, which can generate different standards as to its chromosome distribution. 47 In addition, both 18S rDNA carrier pairs are involved with the NORs and are different between species.
The presence of simple NOR located in the largest chromosome pair is considered a standard feature of cichlids.28,30 However, the results showed that S. lilith had only one 18S rDNA site on the largest chromosome pair of the complement with additional sites from the homologous pair 7, whereas in S. jurupari the 18S rDNA is located in pairs 6 and 9. These results indicate that there is large margin in karyotype variability, which could be related to TEs.48,53–55
Analysis of the distribution of 18S rDNA in the genome of Oreochromis niloticus has shown that some copies of the 18S are consistently flanked by TEs, indicating that these factors may be operating in the dispersed copies of 18S, thereby generating variability and a large number of sites. 48 In Satanoperca, mapping of the Rex1, Rex3, and Rex6 retroelements revealed markings on the pericentromeric region and interstitial and terminal regions of some chromosomes, 56 indicating that association with the 18S rDNA can cause karyotype variability in the genus.
Thus, although S. jurupari and S. lilith harbor conserved karyotype macrostructure, which is considered a basal feature in the cichlid family, karyotype organization in these species revealed that chromosomal rearrangement events have acted on the evolutionary history of the genus. This study revealed that the genus Satanoperca has characteristics specifically regarding genomic organization that allow for the delineation of species, allowing for the use of cytogenetic markers to make taxonomic inferences within the genus.
Footnotes
Acknowledgments
This work was supported by the Universidade Federal do Amazonas (UFAM), the graduate program of INPA Genética, Conservação e Biologia Evolutiva, Conselho Nacional de Pesquisa e Desenvolvimento Tecnológico (grant number: 474617/2013-0), FAPEAM (020/2013), and CAPES (Pró-Amazônia–grant number 23038.009447/2013-45, 3295/2013). FAS received funding from the Fundação de Amparo a Pesquisas do Estado do Amazonas. The English language was revised by Cactus Communications Pvt. Ltd.
Author Contributions
All of the authors performed the experiments, analyzed the data, wrote the article, and read and approved the final article.
Disclosure Statement
No competing financial interests exist.