
Abstract
Abstract
Techniques based on nuclear magnetic resonance (NMR) for imaging and chemical analyses of in vivo, or otherwise intact, biological systems are rapidly emerging and finding diverse applications within a wide range of fields. Very recently, several NMR-based techniques have been developed for the zebrafish as a model animal system. In the current study, the novel application of high-resolution magic angle spinning (HR-MAS) NMR is presented as a means of metabolic profiling of intact zebrafish embryos. Toward investigating the utility of HR-MAS NMR as a toxicological tool, these studies specifically examined metabolic changes of embryos exposed to polymethoxy-1-alkenes (PMAs)—a recently identified family of teratogenic compounds from freshwater algae—as emerging environmental contaminants. One-dimensional and two-dimensional HR-MAS NMR analyses were able to effectively identify and quantify diverse metabolites in early-stage (≤36 h postfertilization) embryos. Subsequent comparison of the metabolic profiles between PMA-exposed and control embryos identified several statistically significant metabolic changes associated with subacute exposure to the teratogen, including (1) elevated inositol as a recognized component of signaling pathways involved in embryo development; (2) increases in several metabolites, including inositol, phosphoryl choline, fatty acids, and cholesterol, which are associated with lipid composition of cell membranes; (3) concomitant increase in glucose and decrease in lactate; and (4) decreases in several biochemically related metabolites associated with central nervous system development and function, including γ-aminobutyric acid, glycine, glutamate, and glutamine. A potentially unifying model/hypothesis of PMA teratogenicity based on the data is presented. These findings, taken together, demonstrate that HR-MAS NMR is a promising tool for metabolic profiling in the zebrafish embryo, including toxicological applications.
Introduction
M
Most recently, various NMR-based techniques for imaging and chemical analysis (including HR-MAS NMR) have been reported for the zebrafish as an increasingly important animal model.2–6 To date, however, NMR has been largely applied to adult stages of zebrafish, and has enabled in vivo imaging, and localized chemical analyses, in this system. With respect to the former, NMR-based microimaging (μMRI) was recently demonstrated with live zebrafish and specifically applied to the detection and characterization of melanomas.2,3 Localized in vivo high-resolution spectroscopy, likewise, was achieved in the brain of adult zebrafish (with a spatial resolution of voxels as small as 3.3 μL) and specifically utilized, as an example, for metabolic profiling of spatially defined regions of the brain including, most recently, effective lipid profiling.4,5 Although the past studies have largely focused on adult stages of the zebrafish, one such study applied NMR, and HR-MAS specifically, for chemical analysis of intact embryonic stages and, in particular, was able to quantify ethanol levels as part of exposure studies. 6
In the present study, HR-MAS was investigated as a means of metabolic profiling of intact zebrafish embryos. As embryonic stages of the zebrafish represent a particularly versatile system with numerous practical advantages, including capacity for high-throughput analysis, the development of HR-MAS-based metabolic profiling would represent a potentially powerful metabolomics tool that could, indeed, be applicable to a wide range of scientific investigations. Among the potential applications of HR-MAS NMR to the zebrafish model is toxicological utility for understanding metabolic changes related to environmental toxins toward both understanding toxicity (i.e., targets, mechanism of action) and identifying potential biomarkers of exposure. Bolstering this potential, the zebrafish has, indeed, been utilized extensively as a toxicological model for a wide array of toxic environmental contaminants, including heavy metals, endocrine disruptors, and various organic pollutants.7–9 In particular, embryonic and subsequent larval stages have proven particularly useful for characterizing teratogenicity (i.e., developmental toxicity).8,9 With respect to the current study, the zebrafish embryo teratogenicity assay (ZETA) has been specifically used in the investigation of toxic, and specifically, teratogenic metabolites from marine and freshwater algae, including cyanobacteria, 10 and has enabled both characterization of known algal toxins11–13 and identification (through screening, bioassay-guided isolation, and chemical/toxicological characterization) of otherwise unknown toxic metabolites.14–16
In one such study, ZETA was used to identify a family of teratogenic secondary metabolites, namely, the polymethoxy-1-alkenes (PMAs), and subsequently demonstrate a taxonomically widespread distribution of these metabolites among both prokaryotic cyanobacteria and eukaryotic algae (i.e., Chlorophyta or green algae).14,16 Accordingly, it was suggested that PMA may represent an emerging class of environmental toxins specifically associated with increasingly frequent and intense blooms of so-called harmful algae worldwide. In fact, PMAs have been previously described from a wide range of cyanobacteria for more than three decades.17–20 However, only recently have they been linked to biological activity and specifically observed teratogenicity in the zebrafish embryo model. 14 As such, no data currently exist with respect to possible mechanisms, or biochemical, molecular, or cellular targets, of these compounds.
Elucidating the effects of toxic compounds, such as the PMAs, at the biochemical and molecular level is not only obviously fundamental to understanding the mode of action but also potentially informative with respect to the assessment of toxic potential through identification of measurable biomarkers of exposure. Accordingly, in the current study, HR-MAS NMR was applied to metabolic profiling of zebrafish exposed to PMAs, at specifically subacute concentrations and exposure times, as a means of identifying relevant metabolic changes toward generating hypothesis regarding mechanisms/targets of these compounds, as well as potential biomarkers, and as a model class of compounds demonstrating, more generally, the potential of the technique as a metabolomics tool for environmental toxicology.
Materials and Methods
Isolation of PMA
The teratogenic PMA variant, 4,6,8,10,12,14,16,18,20-nonamethoxy-1-pentacosene (Fig. 1)—as the typically most abundant congener—was isolated from cultures of the cyanobacterial species, Cylindrospermopsis raciborskii, as previously described.14,16 Briefly, freeze-dried biomass was twice extracted in chloroform and pooled extracts (following filtration) concentrated to dryness in vacuo. Subsequently, the PMA was isolated by silica gel (60 Å, 40–63 μm) with a gradient of ethyl acetate in hexane (eluting with 100% ethyl acetate) and subsequent reverse-phase HPLC (Phenomenex Luna 5 μm C18 100 Å, 250 × 4.6 mm), using a gradient of 50%–100% acetonitrile (and water).

Structure of 4,6,8,10, 12,14,16.18,20-nonamethoxy-1-pentacose as a common and representative PMA. PMA, polymethoxy-1-alkene.
Exposure of zebrafish to PMA
Zebrafish were exposed to PMA by a method modified from previously published protocols.10,11,13–16 Briefly, 100 embryos (12 h postfertilization [hpf]) per replicate (n = 4) were exposed in 35-mm-diameter polystyrene dishes to 20 μg/mL of PMA (in E3 medium 21 ) for 24 h. Exposure concentration was based on previously determined teratogenicity of PMA and specifically represent concentrations that are not acutely toxic (i.e., teratogenic or lethal) to minimize metabolic changes associated with mortality or severe development deformities. 14 Likewise, exposure times <36 hpf (i.e., 12 hpf embryos and 24 h treatment) were selected as previous studies with PMA-determined teratogenicity, in the form of morphological deformities, was observed only after 72–96 hpf. 14 Control embryos were exposed to the solvent carrier (i.e., methanol) alone at corresponding (v/v) concentrations. Following exposure, treated and control embryos were washed (to remove any residual PMA) and carefully transferred to a 4-mm zirconium oxide rotor (Bruker BioSpin AG), to which 10 μL of deuterated phosphate buffer (100 mM; pH 7.0) containing 0.1% (w/v) 3-trimetylsilyl-2,2,3,3-tetradeuteropropionic acid (TSP), as a lock solvent and NMR (i.e., chemical shift) reference, was added. The rotor was immediately transferred to the NMR spectrometer.
HR-MAS NMR studies
All HR-MAS NMR experiments were carried out on a Bruker DMX 600 MHz NMR spectrometer operating with a proton resonance frequency of 600 MHz. The instrument is equipped with a 4-mm HR-MAS dual inverse 1H/13C probe with a magic-angle gradient. All measurements were carried out at a magic-angle spinning rate of 6 kHz and a temperature of 277 K. Temperature was controlled by a Bruker BVT3000 control unit. Bruker TOPSPIN software (Bruker BioSpin AG) was used to acquire and process the NMR data.
One-dimensional 1H HR-MAS NMR spectra were recorded using rotor synchronized Carr–Purcell–Meiboom–Gill (CPMG) pulse sequence with water suppression. Each one-dimensional (1D) spectrum was acquired by applying a spectral width of 8000 Hz, domain data points 16k, number of averages 512 with 8 dummy scans, constant receiver gain of 2048, an acquisition time of 2 s, and a relaxation delay of 2 s. Since NMR measurements were done on intact embryos, the relaxation delay was set to a small value to remove nascent short T2 components due to the presence of lipids. All spectra were processed by an exponential window function corresponding to a line broadening of 1 Hz and zero-filled before Fourier transformation. 1H HR-MAS NMR spectra were phased manually and automatically baseline corrected using TOPSPIN 2.1 (Bruker BioSpin AG). The total analysis time (including sample preparation, optimization of NMR parameters, and data acquisition) of 1H HR-MAS NMR spectroscopy for each sample was ∼20 min.
Two-dimensional homonuclear correlation spectroscopy (1H-1H COSY) was performed in magnitude mode. The parameters used for COSY were 2048 data points collected in the t2 domain over the spectral width of 4k; 512 t1 increments were collected with 16 transients, relaxation delay of 2 s, acquisition time of 116 ms, and presaturated water resonance during relaxation delay. The resulting data were zero-filled with 2048 data points and were weighted with the sine bell window functions in both dimensions before Fourier Transformation.
Data analysis
For quantification of metabolites, NMR data analysis was performed using MestReNova software version 6.0.3-5604 (Mestrelab Research S.L.). The concentrations of the various metabolites in the spectra were determined by comparing the integral peak intensity of the metabolite of interest with that of the TSP peak, after correcting for the number of contributing protons and for tissue weight. All statistical analyses (t-tests and ANOVAs) of the NMR quantification results were performed with OriginPro v. 8. F-values were calculated, and F-values larger than 2.8 (p < 0.05) were considered significant.
Multivariate statistical analysis (i.e., principle component analysis [PCA]) of primary metabolites in the spectra was performed using Bruker software package AMIX (version 3.8.6). The CPMG spectra, collected from embryos, were subdivided in the range between 0.3 and 9 ppm into buckets of 0.04 ppm (total 218 buckets), using Bruker AMIX software (version 3.8.7; Bruker GmbH). The region of 4.80–6.00 ppm was excluded from the analysis to remove the water signal. To compensate for the differences in the overall metabolite concentration between individual samples, the data obtained were mean centered, scaled to unit variance, and then normalized by dividing each integral of the segment by the total area of the spectrum. The resulting data matrix was exported into Microsoft Office Excel (Microsoft Corporation). This was then further imported into SIMCA software (Umetrics AB) for PCA. Correlation coefficients with p < 0.05 were considered statistically significant.
Results
A representative 1D 1H HR-MAS NMR spectrum obtained directly from intact, control zebrafish embryos (12 hpf) is shown in Figure 2. Highly resolved spectra were obtained, and the assignment of the metabolites was further reinstated by comparing the 1H spectra of reference compounds to existing literature values.22,23 The reference compounds in the Biological Magnetic Resonance Data Bank 24 were specifically used for the characterization of the metabolites. For further unambiguous assignment of the metabolite resonances in zebrafish embryos, a two-dimensional (2D), homonuclear (1H-1H), dipolar-correlation NMR spectrum was measured from intact embryos (Fig. 3). The 2D spectrum allowed separation of most overlapping peaks for the coupled spin systems that gave rise to off-diagonal peaks. The 2D NMR clearly reveals separate correlation networks of several metabolites and, on the basis of cross-peaks, unambiguous assignments were made (Fig. 3). The detailed description of all the peaks' assignment of metabolites is given in Table 1.
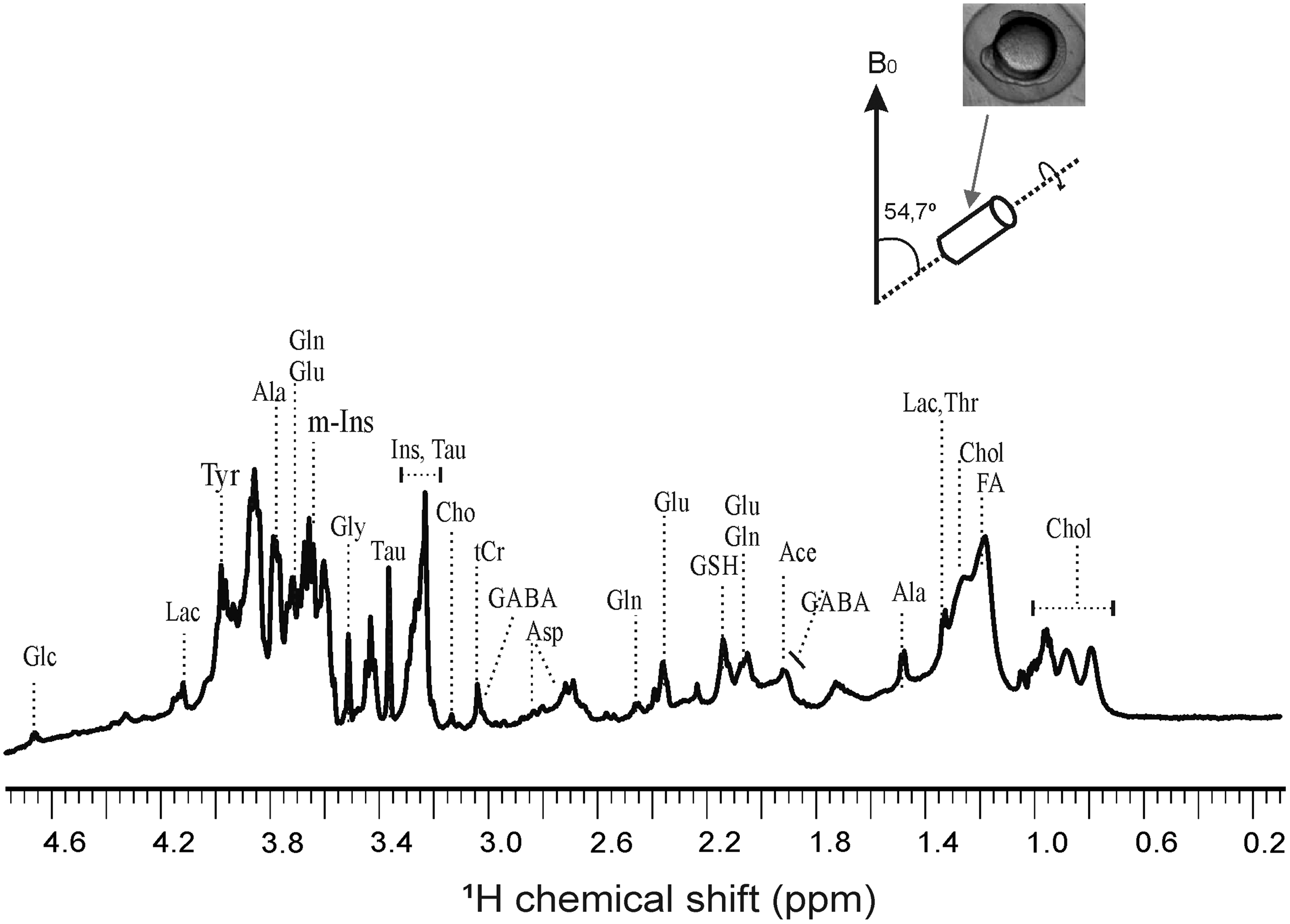
One-dimensional 1H HR-MAS NMR obtained from intact zebrafish embryos (12 h postfertilization). All HR-MAS NMR measurements are done at 4°C with a spinning speed of 6 kHz, cycle delay of 2 s, and total number of scan 256. Water suppression was done for obtaining enhanced signals from metabolites. 1H shift was calibrated using TSP as an internal standard. Glc, glucose; Lac, lactate; Ala, alanine; Gln, glutamine; Glu, glutamate; Gly, glycine; Tau, taurine; m-Ins, (myo)-inositol, Cho, choline; tCr, total creatine; GABA, γ-aminobutyric acid; Asp, aspartate; GSH, glutathione; Ile, isoleucine; Chol, cholesterol; FA, fatty acid; HR-MAS, high-resolution magic angle spinning; NMR, nuclear magnetic resonance; TSP, 3-trimetylsilyl-2,2,3,3-tetradeuteropropionic acid.
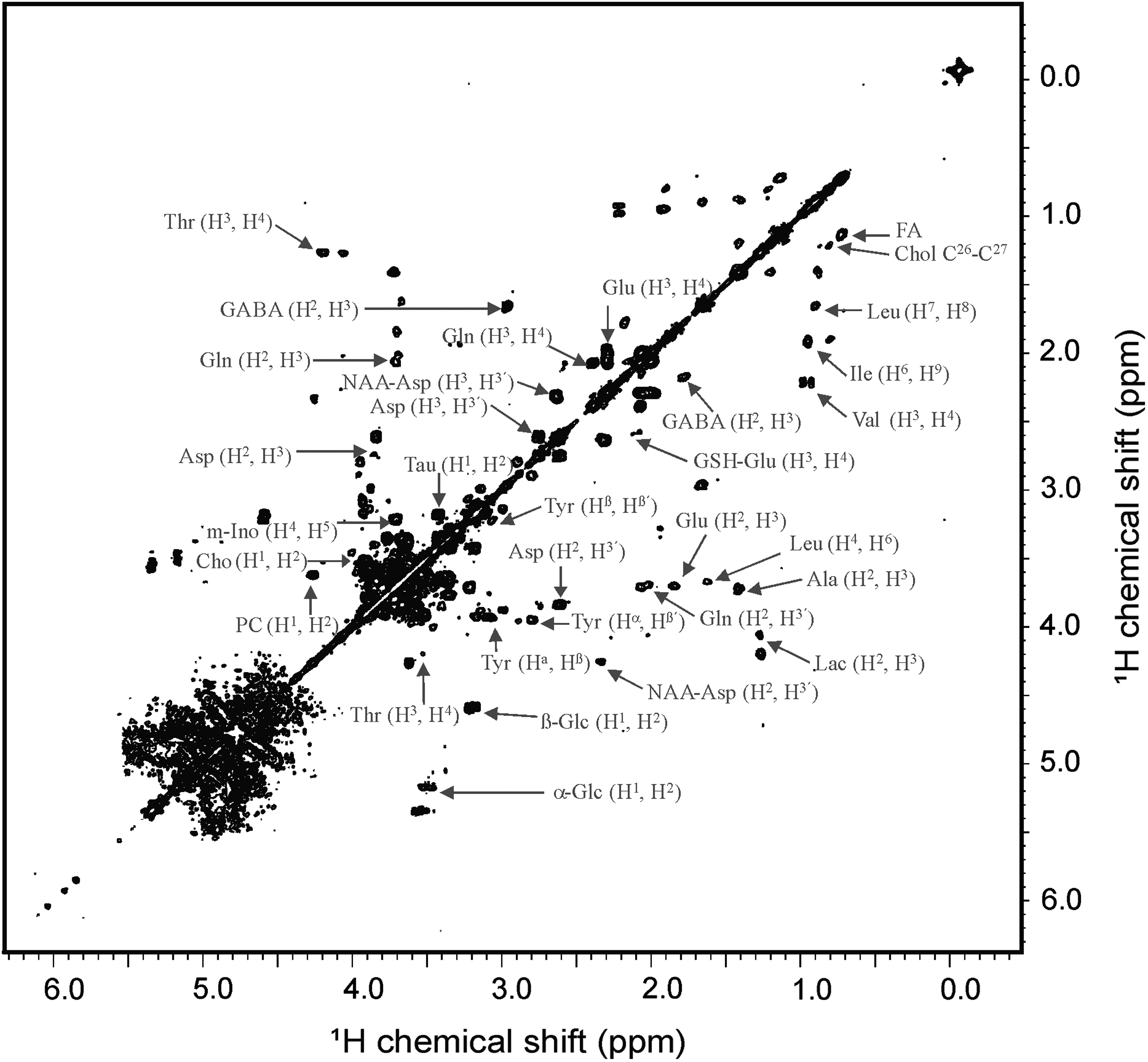
Two-dimensional [1H-1H] homonuclear correlation HR-MAS NMR spectrum obtained from intact zebrafish embryos (12 h postfertilization). The 1H shift was calibrated using TSP as an internal standard. PC, phosphoryl choline; Leu, leucine; Thr, threonine.
1
Comparison of 1D HR-MAS spectra obtained from control and PMA-treated zebrafish embryos is depicted in Figure 4. Quantitative analysis of metabolites in embryos exposed to PMA when compared to control embryos shows statistically significant changes in several identified metabolites in exposed embryos (Fig. 5). The major changes include (1) increased levels of glucose (Glc), (myo-)inositol (m-Ins), cholesterol (Chol), fatty acids (FA) and phosphoryl choline (PC) and (2) decreased levels of glutathione (GSH), lactose (Lac), γ-aminobutyric acid (GABA), glycine (Gly), glutamate (Glu), and glutamine (Gln). There were no observable effects (data not shown) of the PMA at the concentration and exposure time tested, with respect to mortality, developmental deformities, or other relevant endpoints (e.g., hatching rate) that were previously observed at higher concentrations (≥50 μg/mL) and longer exposure times (>72 hpf) for the compound. 14
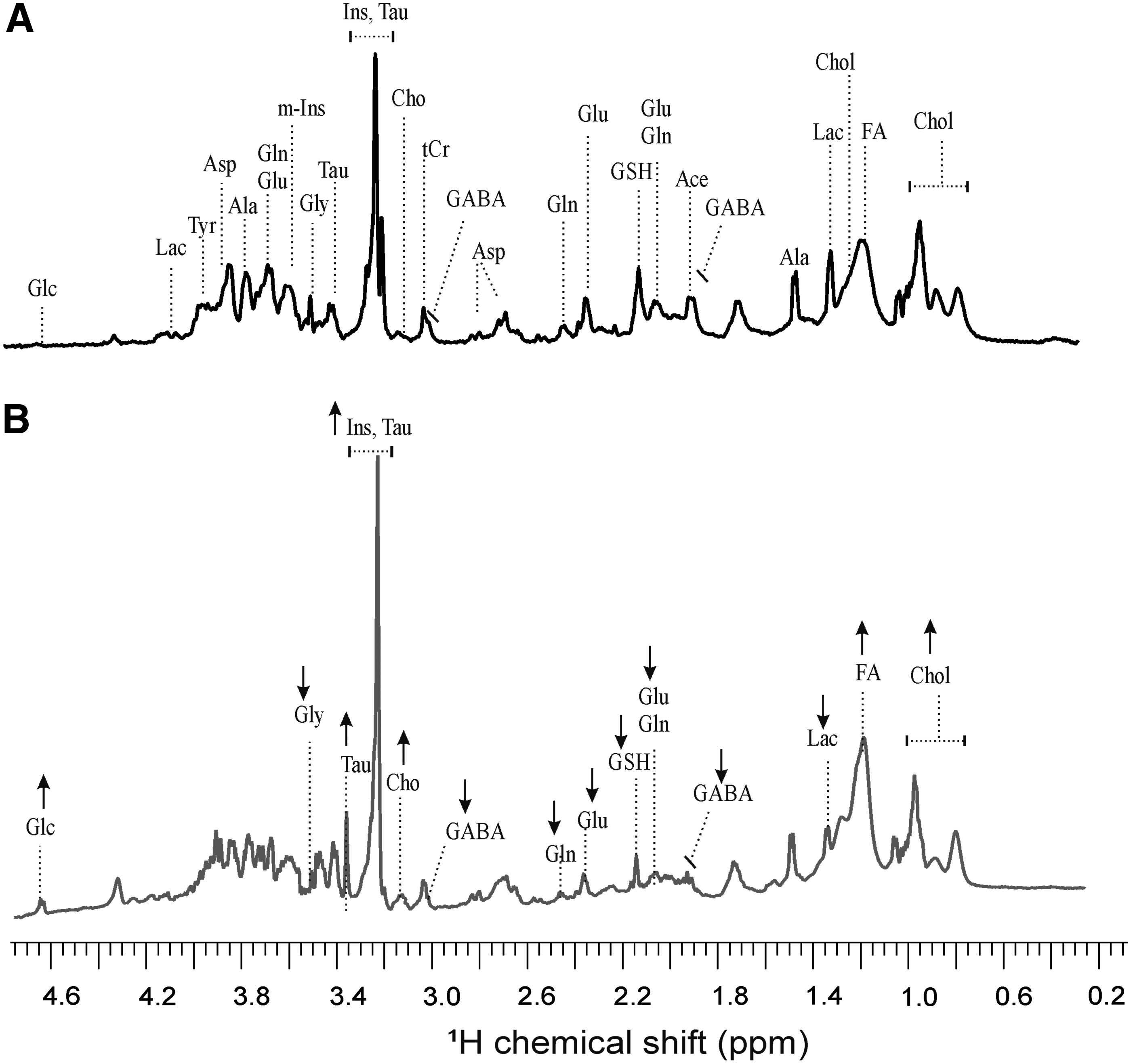
Effect of PMA treatment on the metabolic profile of intact zebrafish embryos measured by one-dimensional 1H HR-MAS NMR. Representative spectra were obtained after treatment of zebrafish embryos (12 h postfertilization) for 24 h with
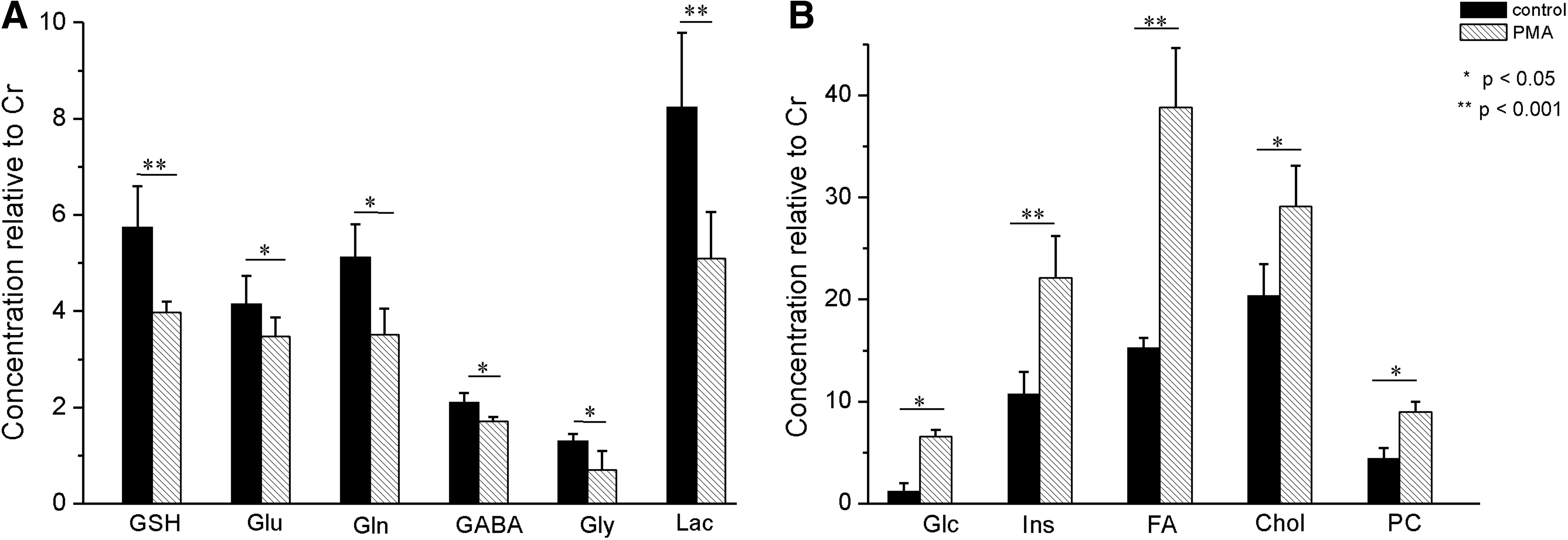
The concentration of metabolites in the zebrafish embryos treated with or without PMA. Zebrafish embryos (12 h postfertilization) were treated for 24 h with either solvent carrier (control) or PMA, followed by washing three times. Representatives of metabolites that show decrease
The 1H HR-MAS spectra were investigated by PCA, and subsequent partial least square discriminant analysis (PLS-DA) modeling (R2 = 0.995, Q2 = 0.999) to probe if control and PMA-treated embryo can be discriminated, and to determine the spectral regions and corresponding compounds mainly responsible for separation. The PLS-DA scores plot and loading plot are presented in Figure 6. The score plot explains 82.4% of total variance of control sample clustering in the negative PC1 scores and PMA-treated samples in the positive PC2 scores (Fig. 6A). To determine the variables, that is, metabolites assigned to the corresponding buckets that are mainly responsible for the separation of two groups, the load values or weights of the PC1 were analyzed (Fig. 6B). Signals coming from m-Ins, PC, Glc, Chol, and FA have a positive score in the PC1 loading plot within 95% significance interval (p < 0.05), and signals coming from GABA, Glu, and Lac have a negative score in the PC1 loading plot. All of these metabolites have loading within 95% confidence level indicating them to be the most significant metabolites for group separation in the score plot.

Discussion
The current study is the first report of HR-MAS NMR as a means of metabolite profiling in the intact zebrafish embryo system (i.e., without any extraction) and generally demonstrates the technique to be an effective and potentially powerful tool for metabolomics studies of early (e.g., ≤36 hpf) developmental stages of this particularly versatile system. Building on previous work and prior NMR-based characterization of the most abundant metabolites found in zebrafish embryos, 2D homonuclear (i.e., COSY) HR-MAS NMR was capable of identifying a wide range of relevant metabolites. Subsequent quantitation of peak area, coupled to PCA, enabled statistical analysis of changes in metabolite profiles.
To explore the applicability of HR-MAS NMR metabolite profiling as a toxicological tool, the technique was specifically applied to the algal-derived PMAs as taxonomically widespread potential contaminants of aquatic systems. Although teratogenicity (i.e., morphological deformities) of PMAs in the zebrafish embryo model has been described, 14 the mechanism of this toxicity and possible biochemical, molecular, and cellular targets remain currently unknown. Consistent with subacute concentrations and exposure times, no discernible effects on development (e.g., mortality, deformities, hatching rates) were observed. However, embryos exposed to PMA were, indeed, characterized by statistically significant changes in several relevant metabolites. Moreover, the changes in metabolic profile, associated with PMA-exposure, notably demonstrate a possible convergence of several interrelated biomarkers, which point to a potentially unifying model of the teratogenicity of these compounds.
As one of the most salient observations, particularly in light of the teratogenicity of the PMAs, inositol levels were found to increase significantly in exposed embryos. Derivatives of m-Ins, and phosphatidylinositol derivatives, in particular, have an established role in Wnt-mediated pathways of embryo development (including zebrafish).25,26 Wnt pathways are a well-described group of signal transduction pathways that regulate crucial aspects of cell-fate determination, cell migration, cell polarity, morphological patterning, and organogenesis during embryonic development. Phosphatidylinositol-based signaling is recognized to serve as a key intracellular function in this regard. 27 Consistent with a possible role of the Wnt pathway is not only PMA teratogenicity in general but also specific inhibition of organogenesis observed in prior studies; more specifically, exposure to PMAs was found to inhibit the formation of both the eye and heart in developing embryos, 14 and both cardiac and ocular development in the zebrafish are, indeed, linked to Wnt.28,29 It is tempting, therefore, to hypothesize a role for inositol through Wnt pathways in the observed teratogenicity of PMAs.
An alternative hypothesis conversely, however, might implicate a more general interaction of PMA with lipids present in cell membranes. Consistent with this hypothesis, a concurrent increase in FA, PC, and Chol was observed for PMA-exposed embryos. These latter metabolites are clearly associated with lipid composition of cell membranes: PC as the “polar head” of many phospholipids and FAs as “non-polar tails,” whereas cholesterol has an established role in maintaining cell membrane integrity and fluidity, and can comprise, in fact, as much as 50% cell membranes. Similarly, inositols (and m-Ins specifically) are components of—and, in turn, are derived from hydrolytic cleavage of—several phospholipids of membranes, including phosphatidylinositols involved in the Wnt pathways.
As such, the combined observation of elevated levels of these four metabolites by HR-MAS may represent a convergence of indicators reflecting possible interactions of PMA with membrane lipids and subsequent disruption of membranes. It has, in fact, been previously proposed based on 3D modeling that PMAs—and the repetitively polymethoxylated backbone, in particular, which forms a hydrophobic “corkscrew”-like conformation—might interact with lipids and consequently disrupt membranes. 30 Accordingly, disruption of cell membranes could be postulated to potentiate consequent hydrolysis of phospholipids (through access to phospholipases) and consequent release (i.e., intracellular elevation) of free PC, m-Ins, and FA. Simultaneously, increased synthesis or, alternatively, release from lipoproteins of cholesterol might, in turn, serve a compensatory response to restore membrane integrity. The increased synthesis of cholesterol as a mechanism to maintain homeostasis has, in fact, precedence in the study of alkylphospholipids which, as proposed for PMA, interact with, and consequently, disrupt membranes. 31
Another compelling observation in our study is the significant decrease in several interrelated metabolites associated with neuronal function and development. Among these, there is a decrease in Lac concurrent with observed increases in Glc. Numerous studies suggest that lactate produced from glucose through glycolysis in supporting glial cells (i.e., astrocytes, oligodendrocytes)—and supplied, in turn, to neurons through the so-called astrocyte-neuron lactate shuttle—is the primary energy source for neurons.32–35 It has been, furthermore, shown that consumption of energy, in the form of lactate, is particularly important during neuronal development. Also, similarly, several lines of evidence suggest that supply of lactate, as a carbon source (as well as energy source), is essential to myelinogenesis in glial cells during embryo development. 36
Consistent with reduced glycolysis is decreased levels of GSH. As a key detoxifying molecule, glutathione is particularly important for removing reactive oxygen species and other cytotoxic species (e.g., methylglyoxal 37 ) generated during glycolysis. The reduced levels of GSH could, therefore, reflect a decreased cellular demand, and thus synthesis, due to the reduced levels of glycolysis. However, the observed reduction in GSH could alternatively, or additionally, be explained (as discussed below) by depletion due to decreased levels of key biosynthetic precursors (i.e., glutamate, glycine) of this tripeptide.
Aligned with proposed effects of PMA on central nervous system (CNS) development, simultaneous decreases in Gly, Glu, Gln, and GABA (as key biomarkers of neuronal function and development) were observed. Glutamate is the most abundant and important excitatory neurotransmitter in the brain, whereas GABA and glycine are the most abundant inhibitory neurotransmitters in the brain and spinal cord/brain stem, respectively. Accordingly, the three metabolites are essential mediators of neuronal activity in the adult brain, but known to play, in addition, a key role in CNS development.38–40 Glutamine, conversely, is a well-known intermediary of GABA and glutamate recycling by way of the so-called glutamate/glutamine cycle between glia and neurons. 41 Similarly, glial cells are recognized to play a key role in the cycling of glycine; specifically, astrocytes function both in the uptake of glycine from synapses and, in turn, supply glycine and L-serine (as a biosynthetic precursor of glycine) to neurons. 42 Furthermore, aside from their well-known roles in CNS function and development, glutamate and glycine are biosynthetic precursors of glutathione, and decreases in the two could, therefore, additionally explain the observed GSH decrease. Indeed, it has been well established that reduction in the precursors (i.e., glutamate, glycine, and cysteine) can, in fact, lead to cellular depletions of GSH in glia and neurons. 43
These convergent findings, taken together at the cellular level, may specifically point to a possible role of myelogenic glial cells of the developing zebrafish embryo with respect to the developmental toxicity of PMAs. Implicating a possible involvement of myelinogenic glia in the observed PMA toxicity, cholesterol, and phospholipids (containing either PC, e.g., sphingomyelin or inositol), which are, as discussed above, elevated in the PMA-exposed embryos are essential components of myelin. In addition, glial cells—and particularly astrocytes—have an established role in both energy supply (glycolysis) to neurons and recycling of glycine, glutamate, and GABA (as discussed above) to neurons.32–35,41,42 Moreover, glial cells are specifically derived from neural crest cells within developing embryos and represent a particularly compelling target for PMAs. In parallel to observed effects on CNS biomarkers, Wnt pathways (as a possible target of PMAs, see above) are well known to control neural crest cell fate (i.e., differentiation and migration), 44 including subsequent role in the development of eye and heart, which are observably affected in the acute teratogenicity of PMAs. 14 Similarly, one of the additional hallmarks of the developmental toxicity of PMAs is lack of development of melanophores which are, likewise, derived from the neural crest through Wnt. 45
Taken together, therefore, the metabolic profile of PMA-exposed embryos would be consistent with a general unifying model, through which cell-specific effects of the PMAs, and particularly, impairment of neural crest (i.e., myelogenic glial) cells through interference with Wnt pathways, could explain observed changes, including (1) elevated free PC, m-Ins, and FA, following proposed disruption of cell membranes and subsequent hydrolysis of phospholipids, along with concomitant compensatory increases in Chol synthesis and/or uptake; (2) increased m-Ins specifically consistent with interference of phosphatidylinositol signaling in Wnt-based pathways, including in particular, determination of neural crest cell fate; (3) decreased Lac/increased Glc indicative of reduced glycolysis in glia which, otherwise, support both myelogenesis and energy demands of neurons; and (4) consequently decreased GABA, Gly, Glu, and Gln suggestive, in general, of a loss of glial cell viability and/or function, for example, neurotransmitter recycling, in relation to CNS development (Fig. 7).
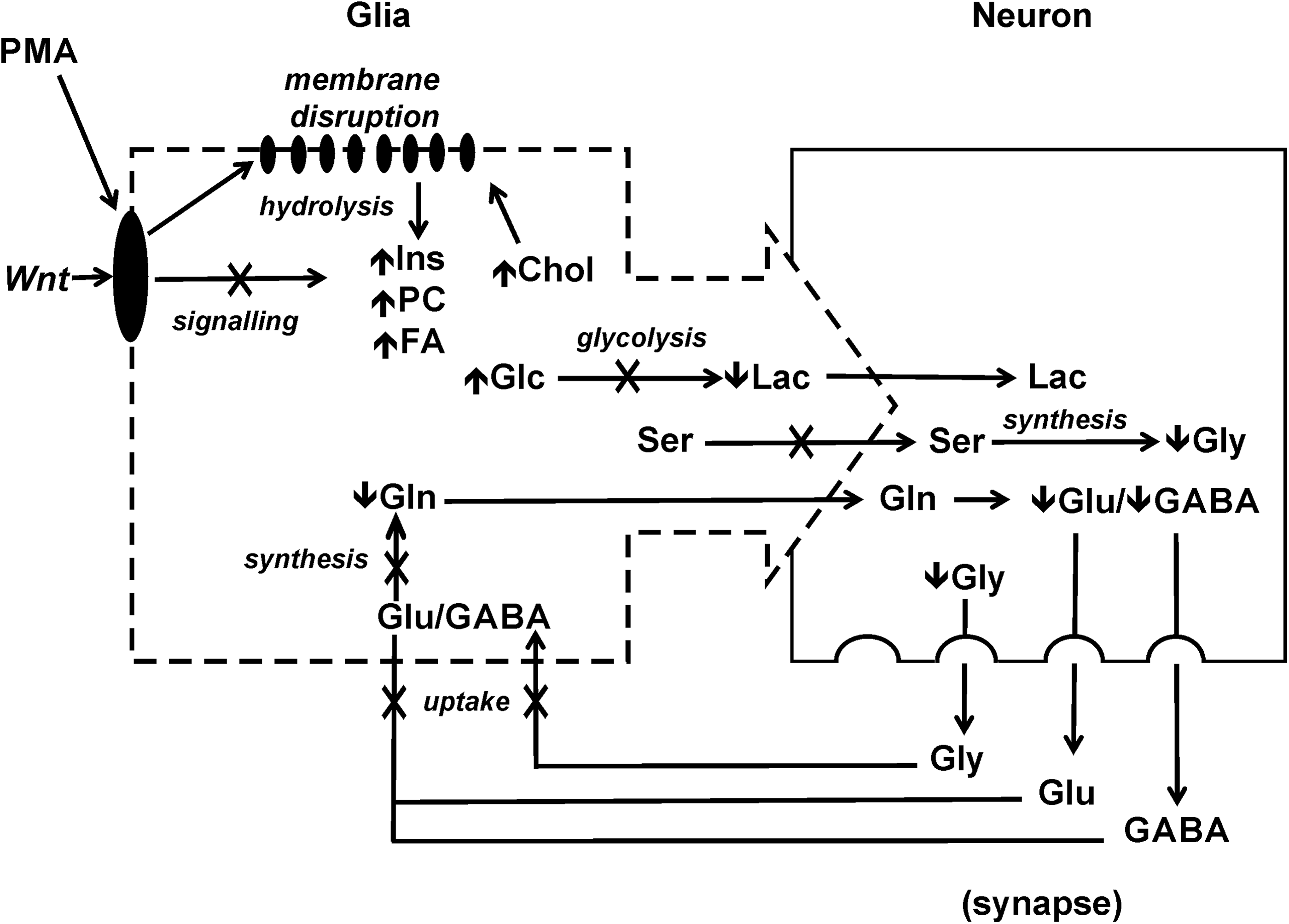
Proposed model of the teratogenicity of PMA in relation to the observed changes in metabolic profile. According to this working model, PMA interacts with membrane lipids, leading to hydrolysis of phospholipids, and consequent release (and, thus, elevated levels) of PC, Ins, and FA, as well as a compensatory increase in Chol. Disruption of membrane lipids would, in turn, interfere with signaling involved in Wnt developmental pathways. The specific inhibition of neural crest-derived glial cells would, thereby, lead to reduced glycolysis (i.e., decreased Lac, increased Glc) as key energy source for neurons, as well as reduced uptake of Glu, and subsequent synthesis of Gln, leading to impaired recycling of Glu and GABA through glutamate/glutamine cycle.
In conclusion, the novel application of HR-MAS NMR to intact zebrafish embryos was found to be, in general, an effective technique for metabolic profiling and specifically a promising approach to identifying potential targets/mechanisms and biomarkers related to toxic, or otherwise bioactive, metabolites (as illustrated, in the current study, by the algal-derived PMAs). Although future experimental studies are clearly required to confirm the model/hypotheses generated by the current HR-MAS analysis, these studies do, indeed, demonstrate the considerable potential of this technique—as a metabolomics tool—for toxicological studies with diverse applicability, in particular, to the zebrafish embryo model.
Footnotes
Acknowledgments
Support for this research was provided by NSF Catalyzing New International Collaboration grant (IIA-1427797). The authors sincerely thank the laboratory of Dr. Hermann Spaink at Leiden University and Dr. Stefan Scholz at Helmholtz Centre for Environmental Research (UFZ) Leipzig for providing zebrafish embryos and associated research support used in exposure studies.
Disclosure Statement
No competing financial interests exist.