
Abstract
Abstract
Dimethyl sulfoxide (DMSO), a widely used carrier solvent, can be toxic to test organisms and has species-specific sensitivity. In this study, the developmental toxicity and stress protein responses of DMSO to rare minnow (Gobiocypris rarus) and zebrafish (Danio rerio) with two tests were compared in the early life stage. In the first test, fertilized eggs were exposed to 0%, 0.0001%, 0.001%, 0.01%, 0.1%, 1.0%, 1.5%, and 2.0% v/v of DMSO until 3 days post hatching. In the second test, larvae from 0 to 8 d were exposed to 2% DMSO until 4 days. Our results showed that DMSO was toxic to rare minnow and zebrafish on multiple indexes, and the no-observed-effect concentrations of DMSO in both species were 1.0% and 0.001% for developmental toxicity analysis and stress protein analysis, respectively. Furthermore, rare minnow larvae were more sensitive than zebrafish to DMSO for spinal malformation. The sensitive period for induction of spinal malformation by DMSO was 0–7 d after hatch (dah) for rare minnow and 0–4 dah for zebrafish. Together, these results will provide support to the use of DMSO in ecotoxicological studies using rare minnow and will contribute to a better understanding of the toxicity of DMSO.
Introduction
D
Rare minnow (Gobiocypris rarus) is a freshwater cyprinid, with lots of attractive features, which make it a suitable model in aquatic toxicity tests and environmental monitoring.7–9 Zebrafish (Danio rerio), a small fish of the family cyprinidae, is also a popular model for aquatic toxicology studies and chemical hazard assessment.10–13 Since 2005, when China REACH (Registration, Evaluation, Authorization and Restriction of Chemicals) came into force, both rare minnow and zebrafish were recommended as organisms for chemical testing. In zebrafish, Hallare et al. (2006) reported that DMSO concentrations <1.5% and 0.01% were appropriate for developmental toxicity and proteotoxicity analyses, respectively. 14 Kais et al. (2013) believed that DMSO may be used without complications as a solvent at a maximum concentration of 0.01%. 15 Less than 0.1% DMSO was often used to make stock solutions of the test substance with no effect on the rate of malformations or survival for zebrafish.16–19 In rare minnow, even though a MAC of 0.01% solvent was recommended for toxicity tests (GB/T 29763–2013), the toxicity of DMSO to rare minnow has not been evaluated to date.
The embryo–larvae toxicity assay was developed to meet the requirements of the 3R principles and has been widely used for chemical testing worldwide.20–23 The common monitoring indicators for embryo–larvae toxicity assay include hatching success, larval abnormalities, and so on.24,25 Aside from those developmental endpoints, the developing embryos may be evaluated for other potential biomarkers to further assess damage caused by the toxicant, such as biochemical (e.g., metallothionein and stress protein) and molecular (DNA damage) responses. Organisms respond to proteotoxicity with the expression of stress proteins, which are able to repair partly denatured proteins. Because the accumulation of these heat shock proteins has been linked to the intensity of stress, these proteins have been regarded as a suitable biomarker in assessing reactions of biota to environmental and physiological stressors.26–29 Heat shock proteins are categorized based on molecular weights. HSP70 is the most responsible for intracellular chaperone and has been well studied within the stress protein family.
In this study, the developmental toxicity and stress protein responses of DMSO to early life stages of rare minnow and zebrafish were compared in two tests. The objectives of the present study are to (1) provide a scientific basis for the use of DMSO in ecotoxicological studies using rare minnow; (2) compare the developmental toxicity and stress protein responses of DMSO in rare minnow and zebrafish; and (3) contribute to a better understanding of the toxicity of DMSO.
Materials and Methods
Chemicals
The solvent tested in this study was DMSO (purity >99.9%; CAS NO.67-68-5; AMRESCO). Before the exposure experiment, test solutions were prepared by dissolving DMSO in aerated reconstituted water (ISO1996: 5.5 mg·L−1 KCl; 294.0 mg·L−1 CaCl2·2H2O; 123.3 mg·L−1 MgSO4·7H2O; and 63.0 mg·L−1 NaHCO3). Tricaine methane sulfonate (MS-222) was obtained from Sigma. All other chemicals used in this study were of analytical grade.
Animals
Adult zebrafish (wild type, AB strain) and rare minnow (Ihb: IHB) were obtained from the Institute of Hydrobiology, Chinese Academy of Science. They were maintained in flow-through feeding aquariums at 26°C ± 1°C with a photoperiod of 12/12 h (light/dark) and fed to satiety twice daily with Chironomus larvae. The preparation and collection of those embryos were performed according to published protocols.30,31 All of the experimental procedures were approved by the Animal Care and Use Committee at Institute of Hydrobiology, Chinese Academy of Sciences (Approval Protocol No. Y21201-1-101) and were in accordance with the National Guiding Principles for the Care and Use of Laboratory Animals.
Embryo and larval toxicity test
At around 2-h postfertilization, eggs were examined under a stereo microscope (SMZ 168; Motic), and only the fertilized eggs were selected for subsequent experiments. Thirty embryos (4–128 cell stage) were randomly distributed into cylindrical dishes (volume 600 ml) containing 400 ml of exposure solution (0.0%, 0.0001%, 0.001%, 0.01%, 0.1%, 1.0%, 1.5%, and 2.0% v/v DMSO) with three replicates. Exposure concentrations of DMSO were based on our previously ascertained range-finding studies, which identified a concentration to induce clear phenotypic effects, and a previous study. 14 Half of exposure solution was replaced daily to keep appropriate concentrations of test compounds and water quality. All experiments were carried out at a temperature of 26°C ± 1°C with dissolved oxygen over 70% and photoperiod of 12/12 h (light/dark). No food was provided until 168 h-postfertilization (hpf). Daily observations for abnormal development and mortality were performed through the lethal and sublethal toxicological endpoints advanced by OECD guidelines 32 and previously published methods.33,34 Cumulative hatching was evaluated at 48, 72, and 96 hpf. Heart rate of embryos was counted microscopically at 48 hpf for 10 randomly selected larvae from each replicate in a room with constant temperature (26°C ± 1°C) and humidity. At 168 hpf, the survival rate was recorded, body length was measured with a Motic SMZ–168 stereomicroscope after anesthetizing by buffered MS-222 with sodium bicarbonate (NaHCO3), then put those larvae (10 larvae per replicate) in one weighing bottle, and measured dry weight after drying to constant weight under 105°C using Mettler Toledo XS 105DU. At the end of the experiment, larvae (10 from each cylindrical dish) were anesthetized and sampled randomly, immediately frozen in liquid nitrogen, and stored at −80°C for subsequent assays.
In experiment one, we found that 2.0% DMSO induced spinal malformation post hatching and the deformity increased with prolongation of exposure time; in addition, the levels of deformity were different between rare minnow and zebrafish. Therefore, we conducted the second experiment to find the sensitive period of the spinal deformities to investigate whether or not there was a species difference in the development toxicity. In the study, 2.0% DMSO solution was prepared and the reconstituted water was used as control with three replicates for each age group. Rare minnow and zebrafish larvae from 0 to 8 d after hatch (dah) were collected for 96-h exposure test. Exposure conditions were consistent with the former test. The larvae were observed daily under the dissecting microscope to detect morphological changes, and the rates of surviving and spinal malformations were recorded after 96-h exposure. At the end of the test, the larvae were moved to the reconstituted water observation for additional 96-h to observe the surviving rate.
HSP70 measurement
The larvae of zebrafish and rare minnow were ultrasonically homogenized for 30 s in 100 μL of Lysis Buffer (Sangon) and then centrifuged at 14,000 g for 15 min at 4°C. The total protein concentration was determined by a BCA Protein Assay Kit (Applygen) and the supernatant was frozen at −80°C for subsequent protein assay. Protein samples (30 μg of total protein per lane) were loaded on SDS/PAGE (12% acrylamide: 0.12% bisacrylamide (w/v), 30 min at 70 V, and 120 min at 120 V), transferred to a PVDF membrane by western blotting (80 V for 60 min at 4°C), and then the membrane was rinsed thrice for 5 min in Tris Buffered Saline (TBS) (NaCl 138 mM, Tris-base 50 mM, KCl 2.7 mM, pH 8.0). Nonspecific binding sites were blocked by immersing the membrane in TBS containing 5% skim milk for 1.5 h at room temperature. The membrane was rinsed thrice every 5 min in Tris Buffered Saline with Tween (TBST)(NaCl 138 mM, Tris-base 50 mM, KCl 2.7 mM, 0.05% Tween-20, pH 8.0) and then incubated at room temperature for 4 h with mouse anti-HSP70 (1:1000 dilution, ab2787; Abcam, Britain) and rabbit anti-beta-actin (1:1000 dilution, bs-0061R; Bioss Beijing Biosynthesis Biotechnology) monoclonal antibodies. After being washed thrice every 5 min, the membrane was incubated with the secondary antibody goat anti-mouse IgG and goat anti-rabbit IgG coupled to peroxidase (1:5000 dilution; Boster) for 1 h at room temperature. After additional washing with TBST for five times, the protein bands in chemiluminescence chromogenic reagents (WBKLS0050; Millipore) were visualized with the Phototope Detection system following the manufacturer's instructions (Gel Doc XR; Bio-Rad). The gray value intensity of HSP70 band in the immunoblots was quantified by densitometric image analysis (ImageJ). Each gray scale was repeated at least thrice.
Statistical analyses
All the experimental data are shown as the mean ± standard deviation (SD). Normality and homogeneity of data were evaluated using the Kolmogorov–Smirnov and Levene's tests, respectively. Statistically significant differences between groups were determined by one-way analysis of variance (ANOVA) and Duncan's multiple range tests using SPSS 19.0. A p-value <0.05 was considered as significant. The no-observed-effect concentrations (NOECs) were determined based on the statistical results. 35 Figures were created using Microsoft Excel 2007 (Microsoft, Inc.) and Adobe Photoshop CS 7.0 (Adobe Systems).
Results
Developmental toxicity of DMSO to rare minnow and zebrafish embryos/larvae
The study showed that only the high concentrations (≥1.5%) of DMSO have toxicity effects on the growth and development of rare minnow and zebrafish embryos/larvae, including decreased survival rate, decreased body length and body weight, increased yolk sac edema, and deflated swim bladder.
The 7-d survival rate was high (100%) in rare minnow exposed to control and lower concentrations of DMSO (≤ 1.0%), but decreased significantly in the group exposed to 2.0% DMSO (Fig. 1A). There was a dose-dependent decrease in the 7-d survival rate of zebrafish exposed to higher concentrations of DMSO (≥0.1%). The decrease was significant in the 1.5% and 2.0% DMSO treatment groups (Fig. 1A). The average heart rate at 48 hpf increased after exposing to lower concentrations of DMSO, but decreased significantly at higher concentrations of DMSO (≥ 1.5%). The NOECs of DMSO to rare minnow and zebrafish in terms of heart rate were 1.0% (Fig. 1B). There was a significant reduction in the body length and body weight of both species exposed to 2.0% DMSO (Fig. 1C, D). Development was unaffected by DMSO exposure up to 1.0% in both species. However, weak to severe developmental abnormal symptoms were observed with increasing DMSO concentration, although the pattern of abnormal symptoms was different between the two species (Table 1). The types of abnormal symptoms primarily included yolk sac edema and weak pigmentation in the rare minnow (Fig. 2), while decreased heart rates and deflated swim bladder in zebrafish (Fig. 3). Cumulative hatching rate at 72 h is rather high at higher concentration of DMSO in both rare minnow and zebrafish. The NOECs of DMSO on the cumulative hatching rate were 1.0% in rare minnow. The cumulative hatching rate at 72 h for zebrafish initially declined as DMSO concentrations increased and then increased as DMSO continued to increase beyond 1.5%. However, no significant effects were observed when exposed to different treatment groups in zebrafish (p > 0.05) (Fig. 4).
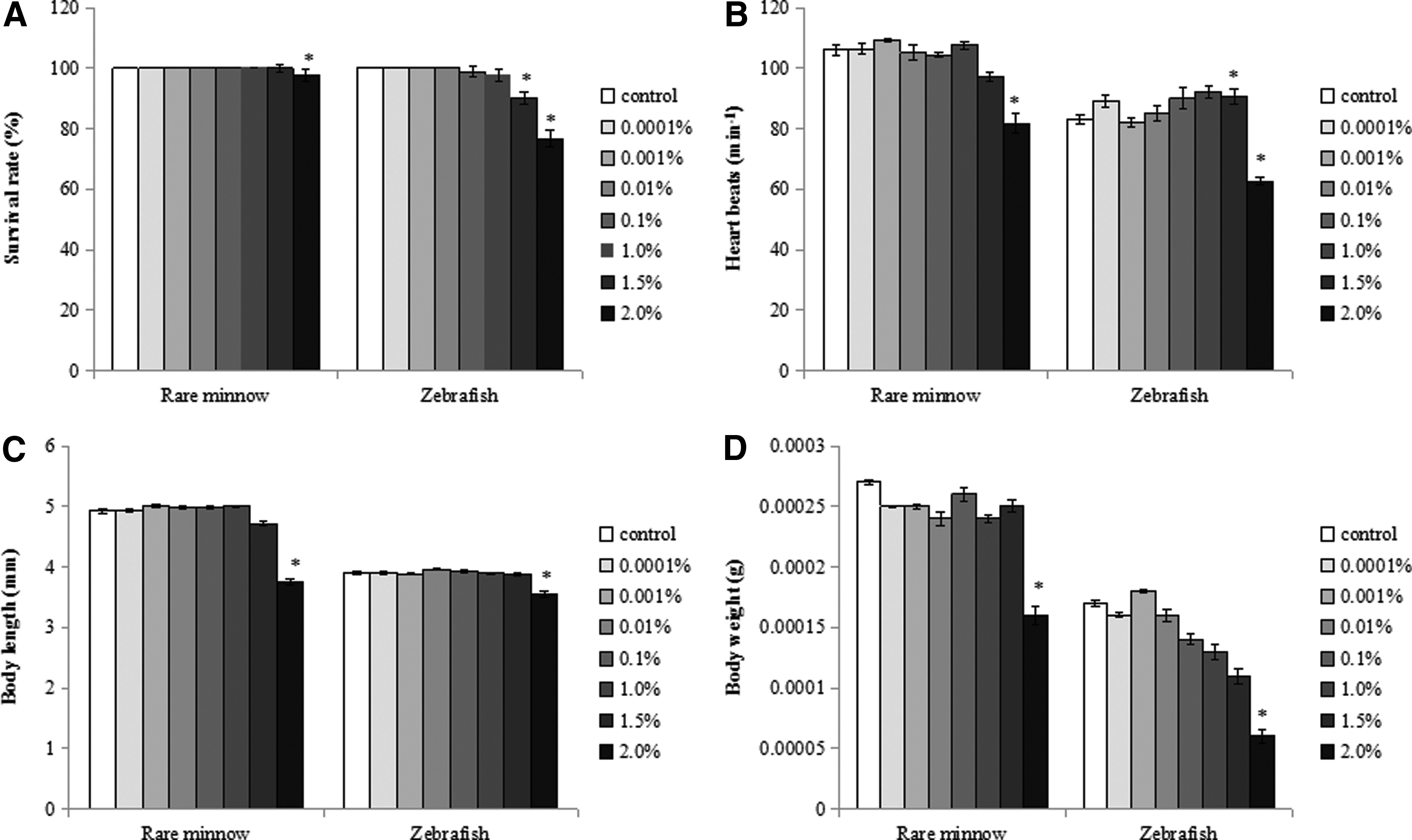
Developmental toxicity of DMSO to rare minnow and zebrafish following semistatic exposure from 2 to 168 hpf.

Microscope images of rare minnow embryos

Microscope images of zebrafish embryos
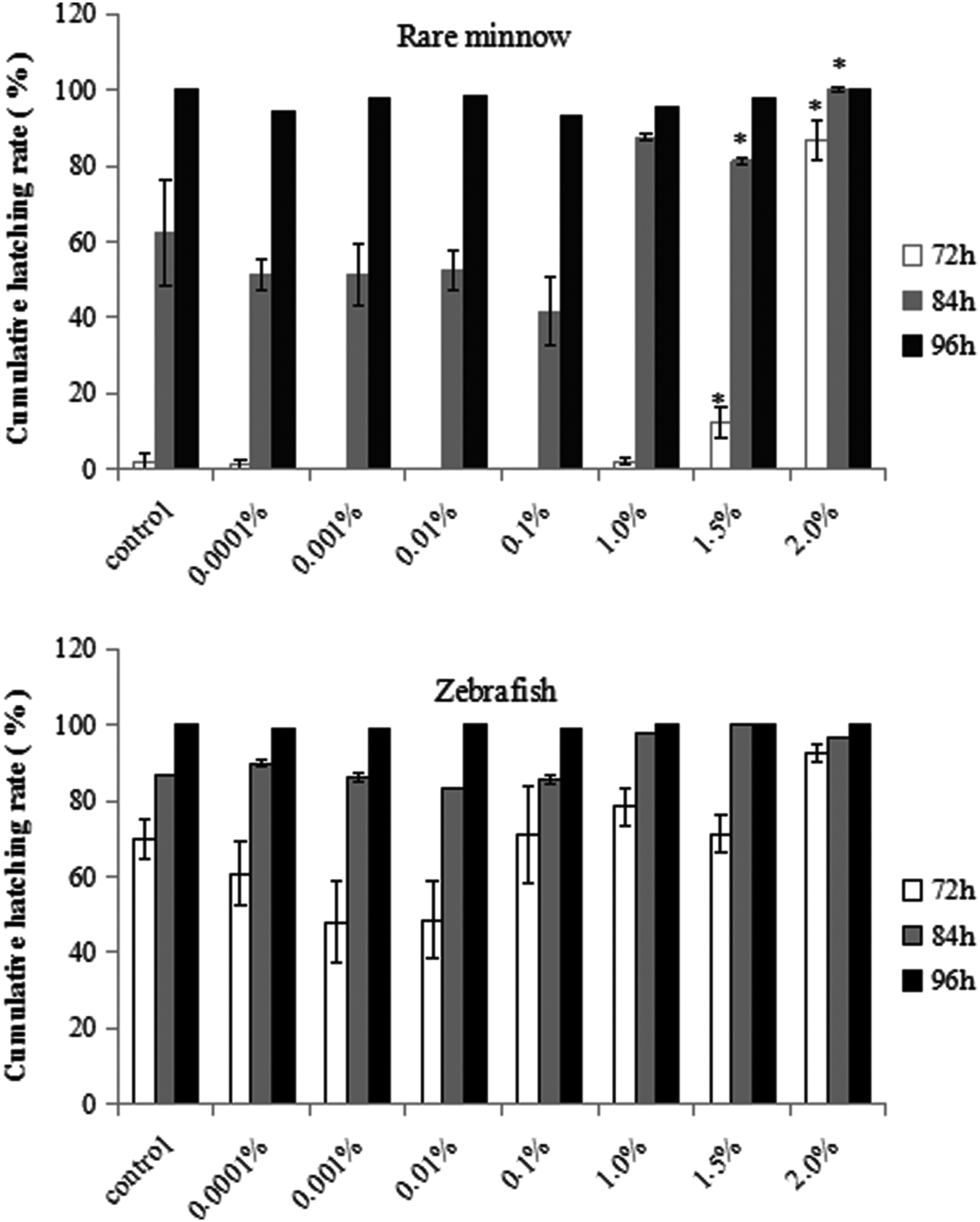
Cumulative mortality rate of rare minnow and zebrafish larvae exposed to different levels of DMSO. Asterisks indicate a significant difference from the control (*p < 0.05).
Asterisks represent a significant difference from the control (*p < 0.05).
Effect of DMSO on the incidence of spinal malformation and the duration of the sensitive period in rare minnow and zebrafish larvae
Spinal malformations were observed in rare minnow and zebrafish, accompanied with abnormal swimming behaviors characterized by whirling in the water (Fig. 5). The frequency of this whirling motion increased with an increasing degree of spinal malformation. Many swirling fish lost the ability to swim and eventually died. All teratogenic larvae exhibited muscle tremors. After transferring these larvae to standard dilution water, these embryos were similarly unresponsive to feed, responded abnormally to touch, and exhibited decreased motor activity compared with the control (data not shown).
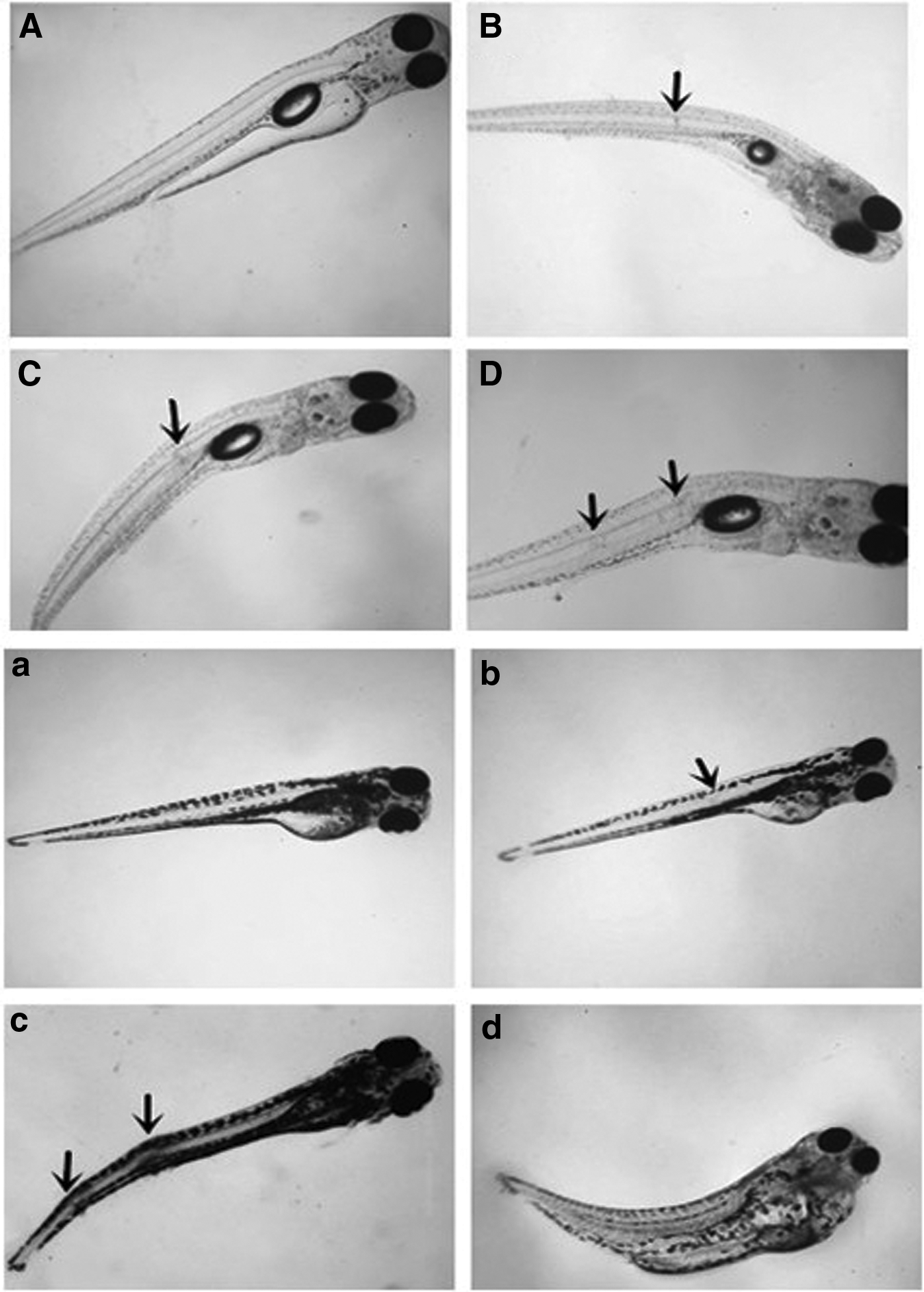
Representative images of spinal deformities in rare minnow and zebrafish exposed to 2.0% DMSO. Rare minnow
The rates of spinal malformation of rare minnow and zebrafish larvae varied widely among the different age groups when incubated in 2.0% DMSO (Table 2). The rates of spinal malformation were characterized by a rapid decrease as age increase for both species. This was particularly true for the rare minnow, as the incidence changed from 65.56% to 37.78% when incubated at 0 dah and 1 dah, respectively. The data also suggested that the malformation rate peaks were at 65.56% and 10.89% at 0 dah for rare minnow and zebrafish larvae, respectively. The sensitive period for induction of spinal malformations was 0–7 dah and 0–4 dah in rare minnow and zebrafish, respectively.
Stress protein response
Western blotting showed that heat shock proteins (HSP70) were detected in all larvae samples taken from control and DMSO treatment groups of rare minnow and zebrafish (Fig. 6A, B). Similar pattern of the content of HSP70 was observed in rare minnow and zebrafish embryos exposed to different concentrations of DMSO (Fig. 6C). For both species, 0.01% DMSO exposure resulted in higher protein content of HSP70 and that decreased as DMSO concentration increased from 0.1% to 2.0%; the NOECs of DMSO in both species were 0.001% for stress protein analysis.

Western blotting for HSP70 expression in rare minnow and zebrafish
Discussion
Developmental toxicity of DMSO to rare minnow and zebrafish
A number of studies have examined the toxicity of DMSO in laboratory animals, including rodents, fish, and amphibians.14,36–39 We assessed the toxicity of DMSO to two fish species recommended for use in testing in China (rare minnow and zebrafish). Exposure to 2.0% DMSO caused decreased heart rates and growth (weight and length), increased larval deformity, and death, which are consistent with previous findings in zebrafish. 14 Zebrafish embryo serves as a widely adopted model for studies of developmental toxicity;40–42 in the study, rare minnow was more sensitive than zebrafish to DMSO based on measurement of cumulative hatching rates and, particularly, spinal malformations. Previous studies have demonstrated that rare minnows are more sensitive to ethynylestradiol (EE2) than zebrafish based on Vtg induction, 43 suggesting that the rare minnow is an ideal model organism for aquatic toxicity tests.
One interesting finding in the present study was that DMSO induced spinal malformations in rare minnow and zebrafish, including spinal flexures, fused vertebrae, raehisehisi and seolios. Similar malformations have also been noted in other species of fish, including Gambusia affinis, Pimephales promelas, and Syngnathus abaster,44–46 after exposing to pollutants such as heavy metals,47,48 pesticides, and industrial wastes.49,50 We often observed disability and distress in affected individuals, indicating that their motor function was severely affected. Furthermore, these fish exhibited poor growth and survival, which suggest that spinal malformations can affect feeding, respiration, swimming, normal morphology, and growth.51–54 Previous studies have shown that DMSO may elevate intracellular Ca2+ levels 55 and other studies have shown that DMSO has estrogenic effects. 56 However, the mechanism by which DMSO induces spinal deformations in rare minnow and zebrafish is still unknown.
Stress protein responses of DMSO in rare minnow and zebrafish
HSP70 is a highly conserved, ubiquitous cellular protein that functions as a molecular chaperone to suppress the aggregation of non-native proteins and promote their refolding and transport to cellular organelles. 57 Induction of HSP70 in response to solvents has been documented in fish.14,58 Consistent with this, we found evidence for induction of HSP70 after exposing to DMSO in the embryo–larval stage of rare minnow and zebrafish (Fig. 6C). However, the concentration range at which induction occurred varied between species. It is interesting that the HSP70 levels initially increased as DMSO concentrations increased and then declined as DMSO continued to increase beyond a threshold for both zebrafish and rare minnow. This attenuation of the stress response following a bell-shaped curve can be explained by the response kinetics of the stress proteins, which have been described previously in other animal groups.14,26,27 Some researchers have speculated that the decrease in HSP70 is offset by the increased incidence of pathological responses among larvae. This was clearly demonstrated in the current results in which at high concentrations of DMSO (≥1.5%), pathologic reactions such as reduction in heart rate, hatching rate, and increased rate of morphological abnormalities in the embryos coincided with a sharp attenuation of the HSP70 induction.
Acceptable levels of DMSO
The NOECs of DMSO were 1.0% v/v in rare minnow and zebrafish for developmental toxicity analysis, which was greater than the MAC values of 0.05% and 0.01% DMSO for aquatic toxicity tests. The NOECs of DMSO in both species for stress protein analysis were 0.001%, which was smaller than the MAC values. The values reported for other species varied widely. For example, in Platynereis dumerilii larvae (38,100 mg·L−1), 59 Platynereis dumerilii embryos (10,630 mg·L−1), 60 and marine microalgae (420–11,000 mg·L−1). 6 Interestingly, we found no difference between rare minnow and zebrafish in terms of the acceptable levels.
Footnotes
Acknowledgments
The study was granted by the National High-Tech Research and Development Projects (2012AA06A302–4). The authors are most grateful to Jiangyan He and Jianghuan Hua for improving the English language.
Disclosure Statement
No competing financial interests exist.