
Abstract
Abstract
Accessing the taxonomy of fish is a basic condition for the conservation of species, and the understanding of their evolutionary relationships. Many Neotropical species of fishes of the order Characiformes are considered incertae sedis because of a lack of consistent evidence of monophyly, with the genus Astyanax among them. With a cytotaxonomic and evolutionary purpose, we compared the karyotype of two species of Astyanax: Astyanax paranae and Astyanax elachylepis. Conventional cytogenetic analyses revealed 2n = 50 chromosomes for the two species, with interspecific and interpopulation karyotype variations. Heterochromatin was primarily localized in the pericentromeric regions, and it was associated with organized regions of the nucleolus (NORs) and/or telomeric regions in some acrocentric chromosomes. The in situ localization of 18S ribosomal DNA (rDNA) and 5S probes varied among A. paranae populations. In A. elachylepis, only one chromosome pair was stained by the 18S probe and another by the 5S probe, confirming a simple NORs pattern. These results support the utility of chromosome markers in Astyanax taxonomy and reveal the conserved characteristics of the karyotype of A. elachylepis, consisting exclusively of two chromosome arms, and the occurrence of simple homologous and non-syntenic sites for 18S and 5S rDNA in this species.
Introduction
A
The Astyanax, the most common and diversified genus in the Characidae family, is widely distributed from the Mexican border with the United States to south of Argentina. It includes ∼150 valid species. 3 These fishes are popularly known as “lambaris” or “piabas,” and they inhabit diverse environments, including headwaters of rivers and streams. 4 It is characterized, along with other genera, as incertae sedis of the family Characidae. 5 Currently, it stands out as a model for various laboratory studies. 6
The genus presents similar morphological forms; however, there is high variability in the number and morphology of the chromosomes, resulting in species complexes.4,7 At least three species complexes are recognized in the genus Astyanax. It includes Astyanax scabripinnis, 7 Astyanax fasciatus,8,9 and Astyanax altiparanae. 10
Astyanax displays wide cytogenetic variation, primarily in the number of chromosomes, karyotype macrostructure, presence of supernumerary chromosomes, heterochromatic block polymorphisms, nucleolus organizer regions (Ag-NORs), variations in the location of ribosomal genes, and triploidy.9,11–20
In addition, studies have shown the occurrence of distinct sympatric cytotypes,8,21 which indicates the diversity of this genus and reinforces the occurrence of cryptic species. However, the amplitude of karyotypic plasticity and specific diversity in Astyanax, as well as its monophyletism are still open questions.
Although previous studies have characterized the karyotype of different species and populations of Astyanax, complementary studies are still necessary to elucidate the karyotype and cytotaxonomic positioning of these fishes. Thus, the goal of this study was to explore the potential of intra and interpopulation markers, using classic methodologies associated with molecular cytogenetics to locate 18S and 5S ribosomal genes. Hence, new data concerning the karyotypic structure and organization of Astyanax elachylepis 22 and Astyanax paranae, removed from the A. scabripinnis complex 23 is presented.
Materials and Methods
The information concerning the Astyanax populations studied are summarized in Table 1 and Figure 1. The fishes were identified by an expert and deposited under LBP 19027 and LBP 19031 vouchers numbers at the Museum of Zoology of the Institute of Biosciences of the State University of São Paulo (UNESP, Botucatu, Sao Paulo, Brazil).
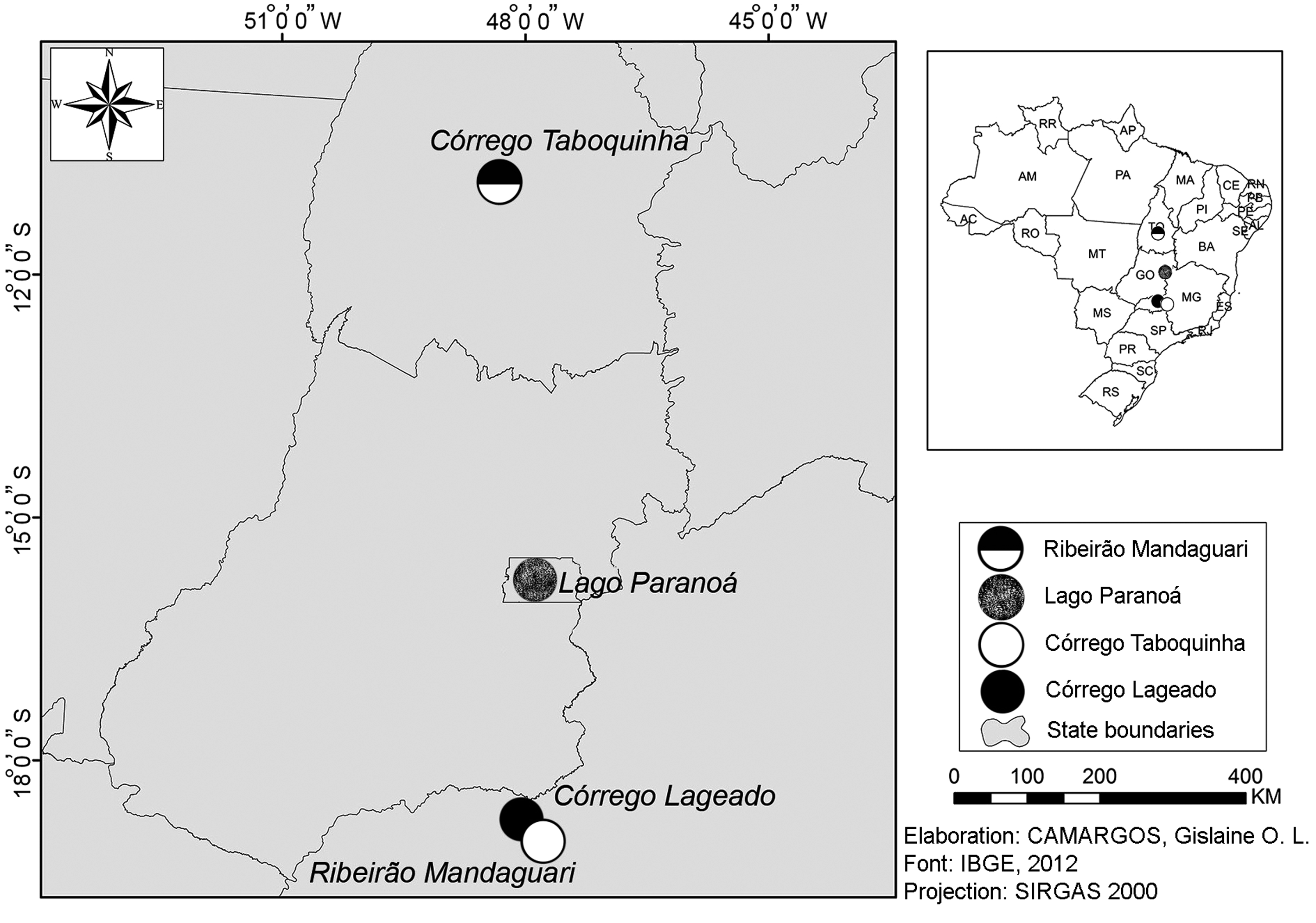
Map of Brazil with indication of collection sites of Astyanax paranae (shaded circles, black and white) in the Paranaíba river basin, and Astyanax elachylepis (circles black and white) in the Tocantins river basin.
DF, Federal District; f, female; m, male; MG, Minas Gerais; N, number of sampled species; TO, Tocantins.
Chromosome preparations of the 50 captured samples were obtained from cells of the anterior kidney, according to Bertollo et al. 24 and Foresti et al. 25 The constitutive heterochromatin was stained using the Summer technique, adapted according to Lui et al. 26 The nucleolus organizer regions (Ag-NORs) were detected according to the methods of Howell and Black. 27 The morphology of each chromosome was established according to the arm ratio (AR) proposed by Levan et al., 28 with the chromosomes arranged in the karyotype in decreasing order by size.
Identification of 18S and 5S ribosomal DNA (rDNA) regions was performed by fluorescent in situ hybridization (FISH), using a 18S labeled probe, obtained by polymerase chain reaction with primers NS1 5′-GTAGTCATATGCTTGTCTC-3′ and NS8 5′-TCCGCAGGTTCACCTACGGA-3′ 29 and a 5S labeled probe with primers A 5′-TACGCCCGATCTCGTCCGATC-3′ and B 5′-GCTGGTATGGCCGTAGC-3′, 30 amplified in the genomic DNA of A. scabripinnis. The labeling of the 18S probe was conducted with the Biotin Nick Translation Mix (Roche), and that of the 5S probe with the Dig Nick Translation Mix (Roche) kit, according to the manufacturer's instructions.
FISH was performed under high stringency conditions (2.5 ng/μL probe, 50% formamide, 2 × saline sodium citrate, 10% dextran sulfate), following the general procedure described by Pinkel et al. 31 The metaphases were analyzed under an epifluorescent microscope (Olympus BX41) coupled with the image capture system DP 71 (Olympus), using the CCD Olympus DP71® system and the DP-Controller software, version 3.2.1.276.
Results
A. paranae and A. elachylepis displayed a diploid number of 2n = 50 chromosomes, with differences in the karyotype formula (Table 2 and Fig. 2). In general, the populations of the two species exhibited marked heterochromatic blocks in centromeric and interstitial regions, and at the end of these chromosomes (Fig. 3a, d, g, j). A. elachylepis had a high number of chromosomes with pericentromeric and interstitial C-positive bands (Fig. 3j). It is noteworthy that the heterochromatin associated with subtelocentric pair 23, carried Ag-NORs in the short arm. The Ag-NORs were simple, as confirmed by FISH with 18S rDNA probe and only occurred in one population of A. paranae (Córrego Lageado), with staining in submetacentric pair 7, and A. elachylepis (Figs. 2, highlighted and 3e, f, k, l), whereas the other two populations of A. paranae (Ribeirão Mandaguari and Lago Paranoá) showed multiple polymorphic sites for Ag-RONs/18S (Figs. 2, highlighted and 3b, c, h, i).

Somatic karyotypes of A. paranae [

Somatic metaphases of A. paranae [
2n, diploid number; a, acrocentric; m, metcentric; NF, fundamental number; sm, submetacentric; st, subtelocentric.
The 5S rDNA sequences were shown by FISH in the two species of Astyanax studied. One submetacentric chromosome pair, with a pericentromeric location of the 5S probe, was conserved in all populations (Fig. 3c, f, i, l). In the A. paranae from the Mandaguari River, this chromosome pair still had 18S rDNA syntenic sites in the short arm (Fig. 3c). The population of A. paranae from Paranoá Lake and A. elachylepis only displayed one submetacentric chromosome pair carrying 5S rDNA (Fig. 3i, l), whereas the populations of A. paranae from the Mandaguari River and the Córrego Lageado had an additional acrocentric chromosome pair carrying the 5S sequences in the short arm (Fig. 3c, f).
Discussion
Our results confirmed a diploid number of 2n = 50 chromosomes for all of species and populations of Astyanax analyzed herein. These data support a conserved condition found in the majority of species, and suggested basal for all Astyanax. 9 In addition, A. elachylepis showed a highly differentiated karyotype, composed exclusively of biarmed chromosomes (14 m + 30 sm + 6 st), which is uncommon in these fishes and far from the karyotype formula described by Tenorio et al. 32 for a population of A. elachylepis from the Araguaia River basin (TO, Brazil), with 6 m + 22 sm +10 st + 12 a. Robertsonian events of centric fission/fusion are required to explain this population difference. On the other hand, these two populations of A. elachylepis share the occurrence of only one pair of chromosomes carrying active Ag-NORs marked by FISH with the 18S rDNA probe. We should highlight that staining occurred in the short arm of the acrocentric chromosome pair in the population from the Araguaia River, whereas in the population from the Tocantins River (present study) the type of chromosomes carrying the NORs was submetacentric. A pericentric inversion event could have diversified this marker in these populations. These differences in karyotype formula (macro and microstructure) are strong evidence of an allopatric speciation process that requires deeper taxonomic study of A. elachylepis.
The three populations of A. paranae studied did not share the same karyotype formula (macrostructure). These populations inhabited the same hydrographic basin of the Paranaiba River and, although all sampling points were in distinct locations along the basin, the karyotype macrostructure showed that the number of metacentric chromosomes (m = 8) was conserved in these populations, and that the inter-population variation in the number of chromosome arms (fundamental number [NF]) was caused by structural alterations in other chromosome classes. Furthermore, interpopulational differences related to the karyotype microstructure referring to the activity of Ag-RONs were shown, along with patterns of constitutive heterochromatin, and location and diversification of 18S and 5S rDNA sequences, reflecting a trend of structuring of these populations. A. paranae could have gene flow restriction due to headwater capture, forming isolated populations. 33
Interspecific differences in karyotype microstructure, specifically in relation to the localization of ribosomal genes are common in Astyanax (reviewed in Piscor et al. 20 ), and among allopatric populations of the same species; for example, A. scabripinnis 34 and Astyanax bockmanni. 19 On the other hand, the fixation of chromosome polymorphic variants, which was observed herein, has a more restrictive occurrence, which suggests that A. paranae formed evolutionarily different population units along the same hydrographic basin. These data are important as cytotaxonomic markers and indicative of incipient speciation processes. They could also aid in decision making in the location of biological conservation areas. This species exhibits an intriguing scenario to be investigated from the perspective of population genetics; however, this was not the goal of this study.
Besides structural chromosome rearrangements, other mechanisms are required to explain the diversification in the karyotype microstructure of the populations analyzed herein, especially in relation to 18S and 5S rDNA. According to Mantovani et al., 35 there are generally multiple locations of 18S sequences in Astyanax, and they can vary in different populations of the same species. In this study, we found multiple 18S locations in two of three populations of A. paranae, accompanying a polymorphism in the activity of these same regions when compared to staining Ag-NORs. The occurrence of bitelomeric staining in one chromosome of the population of Lago Paranoá is also noteworthy. Staining of bitelomeric 18S rDNA was also observed in a single subtelocentric chromosome in other populations of A. paranae18,36 and other species of the genus, such as in Astyanax hastatus, 37 highlighting that events of distribution of the sequences should occur in this species, probably because of translocations between non-homologous chromosomes and/or transposition events. Another peculiar condition was found for the submetacentric chromosome pair, which stains the pericentrometric location of the 5S rDNA, in synteny with the 18S rDNA in the short arm.
Regarding the syntenic 18S/5S, it has been proposed that it is a conserved condition between Astyanax. 38 Our data for the A. paranae population of the Ribeirão Mandaguari support this hypothesis, even though this was not observed in other populations of A. paranae and A. elachylepis. The colocalization of 18S rDNA with transposable elements might be involved with genome evolution and karyotype diversification in A. scabripinnis, 39 and could occur in other species of the genus. This evidence shows that 18S/5S synteny does not prevent the action of evolutionary forces and karyotype diversification of this linkage group in Astyanax.
In conclusion, our results reinforce the utility of cytotaxonomic markers in Astyanax and reveal a conserved condition for the A. elachylepis karyotype, formed preferentially by the biarmed chromosomes, and the occurrence of simple homologous and non-syntenic sites for 18S and 5S rDNA in this species. In addition, populations of A. paranae of the same hydrographic basin show a greater diversification of karyotype formula (macro and microstructure), suggesting that this is a new cryptic species complex.
Footnotes
Acknowledgments
The authors would like to thank Dr. Ricardo Cardoso Benine for taxonomic identification of the species. L.P.S. received a scholarship from CAPES (Coordination for the Improvement of Higher Education Personnel). The work also received the financial support of CNPq (National Council for Scientific and Technological Development).
Disclosure Statement
No competing financial interests exist.