
Abstract
Abstract
Repetitive sequences and their chromosomal locations have been widely studied in species of the Astyanax genus. However, the chromosomal organization of U2 snDNA remains largely unknown. The aims of this study were to examine the chromosomal contexts of U2 snRNA and 5S rRNA genes in Astyanax species and determine the degree of chromosome morphological similarity between species with different diploid numbers. Clusters of U2 snDNA and 5S rDNA were determined in nine species of Astyanax, including two karyomorphs of Astyanax fasciatus Cuvier, 1819. All species exhibited U2 snDNA clusters on two chromosome pairs, except Astyanax mexicanus De Filippi, 1853 (one pair). The 5S rDNA clusters were located on one chromosome pair in Astyanax altiparanae Garutti and Britski, 2000, and Astyanax marionae Eigenmann, 1911, two pairs in Astyanax abramis Jenyns, 1842, Astyanax asuncionensis Géry, 1972, Astyanax bockmanni Vari and Castro, 2007, Astyanax eigenmanniorum Cope, 1894, A. fasciatus (karyomorphs I and II), and Astyanax schubarti Britski, 1964, and four pairs in A. mexicanus. The relationships between the repetitive sequences in different species suggest that A. schubarti and A. mexicanus exhibit an unusual U2 snDNA chromosomal format as a result of events occurring in the evolutionary history of the Astyanax group.
Introduction
T
The chromosomal locations of many repetitive sequences are well characterized in Astyanax, nevertheless some repetitives show variations related to cluster numbers, as the locations of 18S ribosomal DNA (18S rDNA). For instance, Fernandes and Martins-Santos 8 described four and seven sites of 18S rDNA in different A. altiparanae populations from the Paraná River Basin (Paraná state—PR, Brazil), whereas Peres et al. 9 described only a single 18S rDNA site in a population of A. altiparanae from the upper Paraná River Basin (São Paulo state—SP, Brazil). Conversely, fluorescent labeling showed that 5S rDNA location was conserved to the same chromosome pair in these A. altiparanae populations.
The chromosomal locations of histone and U small nuclear RNA (U snRNA) sequences are conserved.10–13 More recently, Piscor and Parise-Maltempi 14 studied eight species of Astyanax and demonstrated that the chromosomal locations of H3 histone gene clusters were highly conserved in A. abramis, A. asuncionensis, A. altiparanae, A. bockmanni, A. eigenmanniorum, and A. fasciatus. Silva et al. 13 observed that the U1 and U2 snDNA clusters were located at different chromosomal sites in different Astyanax species but exhibited strong conservation in the number of sites per genome.
Considering Astyanax a group with wide distribution and distinct cytogenetic features, the aims of this article were to compare the chromosomal organization of U2 snDNA and 5S rDNA in nine species, including those with different diploid numbers, and determine parameters underlying the chromosome evolution of U2 snRNA genes in the Astyanax genus.
Materials and Methods
Sampling and classical cytogenetics
Astyanax specimens were obtained from locations in Brazil as follows: three A. abramis and four A. asuncionensis specimens from the Bento Gomes River in Mato Grosso state (MT), six A. marionae specimens from the Rio Claro stream in Mato Grosso state (MT), five A. altiparanae specimens and one A. schubarti specimen from the Piracicaba River in São Paulo state (SP), three A. bockmanni specimens from the Iguatemi River in Mato Grosso do Sul state (MS), two A. aff. fasciatus specimens (karyomorph I) from the Corumbataí River tributary (SP), five A. fasciatus specimens (karyomorph II) from the Ribeirão Claro River (SP), and three A. mexicanus and three A. eigenmanniorum specimens from aquariophiles in Brazil. Chromosomes were obtained as described by Foresti et al., 15 and chromosome morphologies were determined according to the arm ratios (the most frequently used classification system for fish chromosomes in Brazil), as cited by Piscor et al. 16
Isolation of repetitive DNA probes and fluorescence in situ hybridization
Genomic DNA was extracted from fin samples as described by Sambrook and Russell. 17 The 5S rDNA probe was prepared using polymerase chain reaction (PCR) with primers described by Pendás et al. 18 and Martins and Galetti 19 (A, 5′-TAC GCC CGA TCT CGT CCG ATC-3′; and B, 5′-CAG GCT GGT ATG GCC GTA AGC-3′). The U2 snDNA probe was prepared using PCR with primers described by Bueno et al. 20 (U2F, 5′-ATC GCT TCT CGG CCT TAT G-3′; and U2R, 5′-TCC CGG CGG TAC TGC AAT A-3′). The 5S rDNA probe was labeled by PCR with biotin-14-dATP (Invitrogen, San Diego, CA), and the U2 snDNA probe was labeled by PCR with digoxigenin-11-dUTP (Roche, Mannheim, Germany). Probes labeled with digoxigenin-11-dUTP were detected using antidigoxigenin–rhodamine (Roche), and probes labeled with biotin-14-dATP were detected using Alexa Fluor 488-conjugated streptavidin (Invitrogen). Single- and two-color fluorescence in situ hybridization (FISH) was performed using mitotic metaphasic chromosomes, according to Pinkel et al. 21 and with modifications as described by Cabral-de-Mello et al. 22 Chromosomes were counterstained with VECTASHIELD Mounting Medium (Vector, Burlingame, CA) containing DAPI (4′,6-diamidino-2-phenylindole). Chromosomes and fluorescent signals were visualized with an Olympus BX51 microscope coupled to a digital camera (Olympus model D71). Images were captured using DP Controller software.
Results
Species with 2n = 50 chromosomes were A. abramis, A. asuncionensis, A. altiparanae, A. bockmanni, A. eigenmanniorum, A. mexicanus (Figs. 1A–F and Table 1), and A. aff. fasciatus (karyomorph I; the first described karyomorph for this population) (Fig. 2A and Table 1). All the examined A. mexicanus cells contained one acrocentric B chromosome (Fig. 1F, box). Species with smaller diploid numbers were A. marionae (2n = 48 chromosomes), A. fasciatus (karyomorph II; 2n = 46), and A. schubarti (2n = 36) (Figs. 2B–D, respectively; Table 1).

Locations of U2 snDNA and 5S rDNA clusters on chromosomes of Astyanax species with 2n = 50 chromosomes.
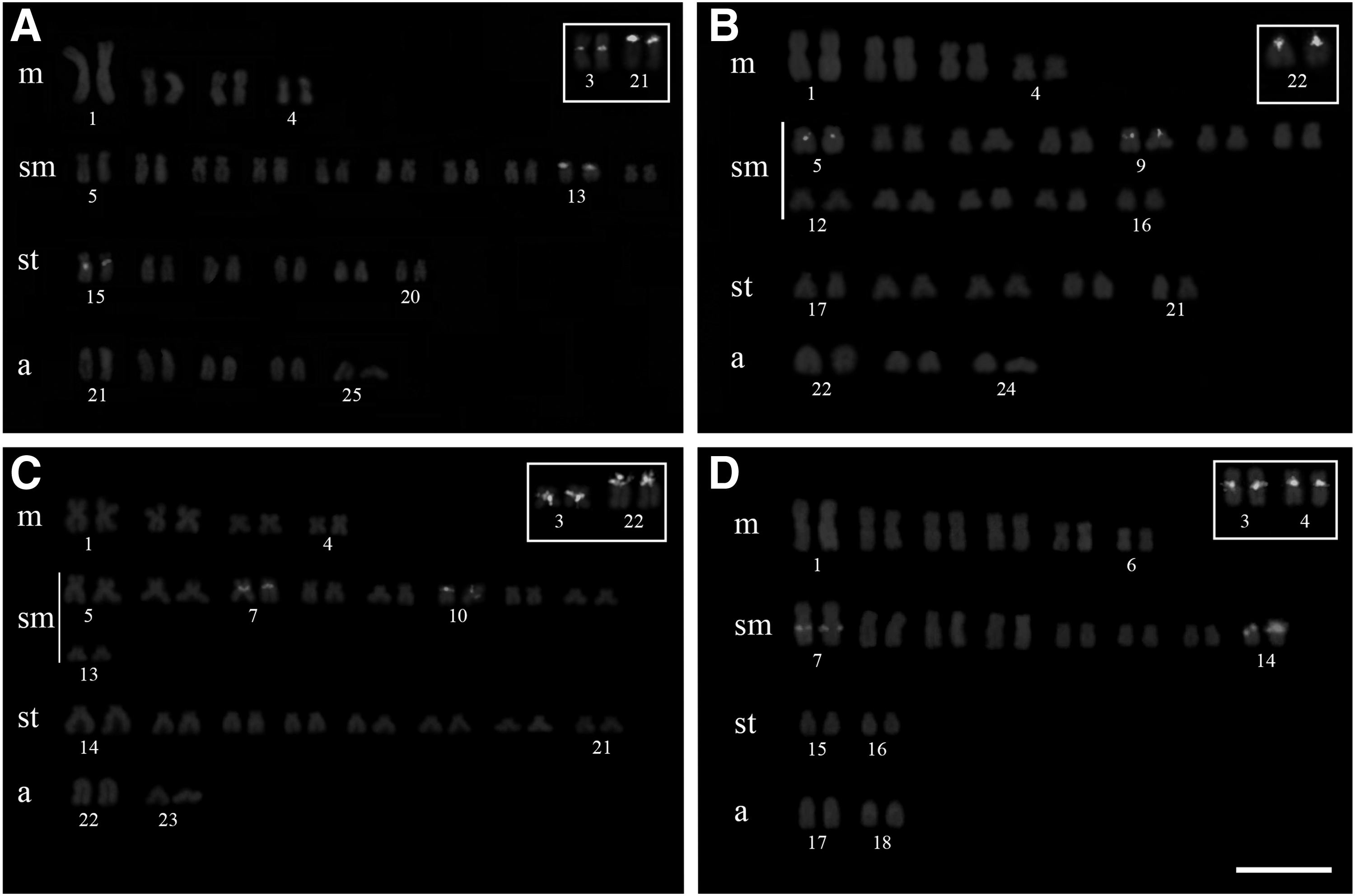
Locations of U2 snDNA and 5S rDNA clusters on chromosomes of Astyanax marionae (2n = 48), Astyanax schubarti (2n = 36), and two Astyanax fasciatus populations (karyomorph I, 2n = 50, and karyomorph II, 2n = 46).
Fundamental numbers.
Numbers of clusters (5S rDNA).
Numbers of clusters (U2 snDNA).
B chromosomes.
FN, fundamental number.
The U2 snDNA clusters were observed on two chromosome pairs in eight of the nine Astyanax species (including the two A. fasciatus karyomorphs) (Figs. 1 and 2; Table 1). In A. mexicanus, U2 snDNA was observed on only one chromosome pair (Fig. 1F and Table 1). The U2 snDNA clusters were located on chromosome pairs 17 and 20 (sm) in A. abramis (Fig. 1A), pairs 11 and 13 (sm) in A. asuncionensis (Fig. 1B), pairs 10 and 13 (sm) in A. altiparanae (Fig. 1C), pairs 5 and 11 (sm) in A. bockmanni (Fig. 1D), pairs 15 (sm) and 16 (st) in A. eigenmanniorum (Fig. 1E), pair 8 (sm) in A. mexicanus (Fig. 1F), pairs 13 (sm) and 15 (st) in A. aff. fasciatus (karyomorph I) (Fig. 2A), pairs 5 and 9 (sm) in A. marionae (Fig. 2B), pairs 7 and 10 (sm) in A. fasciatus (karyomorph II) (Fig. 2C), and pairs 7 and 14 (sm) in A. schubarti (Fig. 2D). All fluorescent signals were located in the pericentromeric regions, with the exception of an interstitial signal observed on pair 7 in A. schubarti (Fig. 2D).
In five of the seven species with 2n = 50 chromosomes (A. abramis, A. asuncionensis, A. bockmanni, A. eigenmanniorum, and A. mexicanus), the 5S rDNA was located on pair 2 (m) (Figs. 1A, B, D–F, respectively). In A. altiparanae, the 5S rDNA was located on pair 5 (sm) (Fig. 1C), and in A. aff. fasciatus (karyomorph I), the 5S rDNA was observed on pairs 3 (m) and 21 (a) (Fig. 2A). Species with 5S rDNA on pair 2 (m) also exhibited signals in pericentromeric regions on other chromosomes. Pericentromeric fluorescent signals were noted on pair 23 (st) in A. abramis (Fig. 1A), pair 25 (a) in A. asuncionensis (Fig. 1B), pair 19 (a) in A. bockmanni (Fig. 1D), pair 22 (a) in A. eigenmanniorum (Fig. 1E), and pairs 15 (sm), 22 (a), and 25 (a) in A. mexicanus (Fig. 1F). The three species with smaller diploid numbers, A. marionae (2n = 48), A. fasciatus karyomorph II (2n = 46), and A. schubarti (2n = 36), harbored 5S rDNA clusters at pericentromeric regions on pair 22 (a) (A. marionae; Fig. 2B), pairs 3 (m) and 22 (a) (A. fasciatus karyomorph II; Fig. 2C), and pairs 3 and 4 (m) (A. schubarti; Fig. 2D).
A summary diagram indicating the chromosomal locations of the U2 snDNA clusters in the nine Astyanax species is shown in Figure 3. Note that three groups were formed: the first group with two chromosome pairs bearing U2 snDNA clusters shared by several species (Fig. 3A), the second group with only one pair (Fig. 3B), and the third group with two pairs but the first pair with interstitial clusters (Fig. 3C).
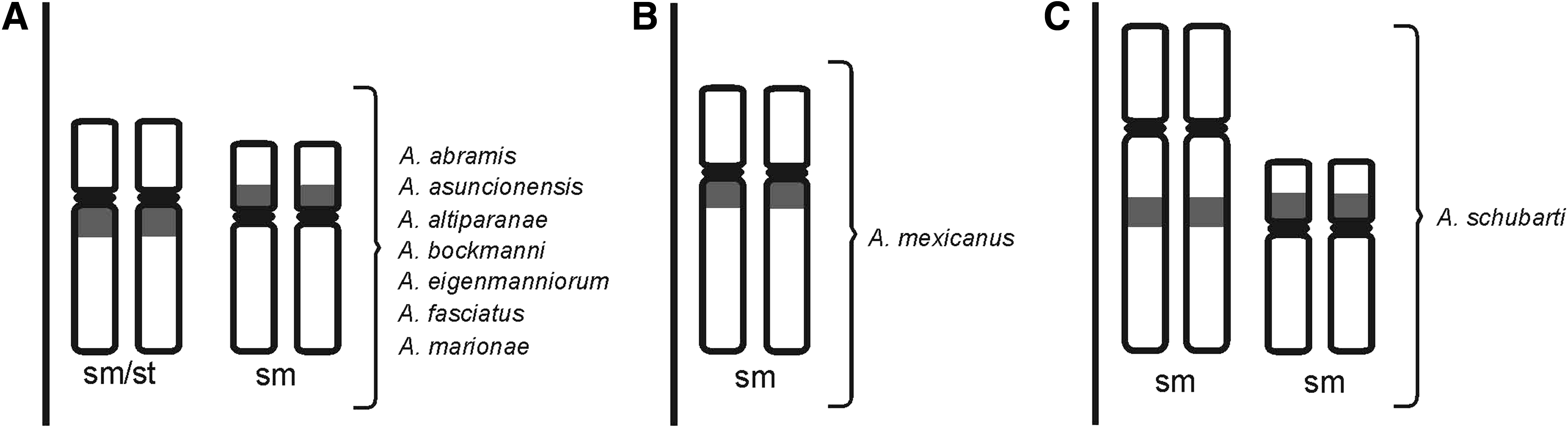
Diagram indicating the chromosome pairs bearing U2 snDNA clusters in the nine Astyanax species.
Discussion
As demonstrated by previous cytogenetic observations, the most common diploid number in the Astyanax genus is 2n = 50 chromosomes. This is consistent with the majority of species in the family Characidae. 2 However, other diploid chromosome numbers are observed in the Astyanax genus, such as the species with 2n = 36, 46, and 48 examined in this study. Furthermore, species complexes with variable diploid numbers are found in Astyanax, such as the “scabripinnis complex” and the “fasciatus complex” (see, e.g.,23,24). Here, A. fasciatus karyomorphs from the same river system were examined that had two different diploid numbers (2n = 46 and 50). Astyanax fasciatus is known to have several karyomorphs, mostly with 2n = 46, 48, and 50 chromosomes. 7
The Astyanax genus is distinguished cytogenetically by diploid number variability from 2n = 36 chromosomes for A. schubarti 2 and Astyanax correntinus Holmberg, 1891, 25 to 2n = 50 chromosomes for most species, for example, A. altiparanae and A. bockmanni.3,4 Variations in karyotype formula and fundamental number (FN) are also widely observed even in populations of the same species. For example, Fernandes and Martins-Santos 3 reported two different karyotype formulae and FN in two A. altiparanae populations from the Índios and Paraná rivers (PR, Brazil), and these values differ from those of the Piracicaba River (SP, Brazil) studied here. In contrast, FISH examination has identified that repetitive sequences showed similar chromosomal locations in Astyanax species.
Chromosomal locations of 5S rDNA are conserved in some Astyanax and exhibit three forms. 26 The form found in most species, including the A. fasciatus karyomorphs analyzed here, exhibits one metacentric pair and one acrocentric or subtelocentric pair with 5S rDNA sites located on the long arm, both near the centromere. 26 According to Vicari et al., 27 Astyanax species with two chromosome pairs bearing 5S rDNA sites exhibit probable synapomorphic features.
In fish, 5S rDNA and other repetitive DNA clusters may be located on the same chromosome pair. For example, while 5S rRNA and histone genes can occur on the same chromosome in Astyanax species,10,14 the 5S rDNA is close to 18S rDNA clusters in Bryconamericus aff. iheringii Boulenger, 1887, 16 another characid fish. The chromosomal locations of the 5S rDNA and U2 snDNA clusters were not consistently linked in the Astyanax species examined here because that these represent two distinct classes of repetitive DNA with completely different functions. Supiwong et al. 28 found that U2 snDNA and 5S rDNA sequences were also carried on different chromosome pairs in the naked catfish Mystus bocourti Bleeker, 1864 (Siluriformes). This spatial separation of 5S rDNA and U2 snDNA appears to be a common feature in fish (see, e.g.,29,30).
All the species examined here had clusters of U2 snDNA on two chromosome pairs, with the exception of A. mexicanus (one pair; Fig. 3). Therefore, our results suggest that the two pairs with U2 snDNA clusters may represent a similar form shared by species of the first group (Fig. 3A), and the only one chromosome pair of A. mexicanus (Fig. 3B) and two pairs of A. schubarti (Fig. 3C) represent different forms of genomic organization of U2 snRNA genes. These two different forms may be explained due to probable reduction in the diploid number (2n = 36) of A. schubarti and ancient separation of A. mexicanus from the South America as previously proposed by Piscor and Parise-Maltempi. 14
Recently, Silva et al. 13 found U2 snDNA clusters on two chromosome pairs in different Astyanax species, except in Astyanax jordani Hubbs and Innes, 1936 (one pair). Silva et al. 13 also showed that, while U1 and U2 snRNA genes were located on different chromosome pairs in different species, the numbers of U1 and U2 sites per genome were strongly conserved. Martins and Galetti 19 proposed that 5S rDNA on a single pair of chromosomes probably represented a more ancient genomic condition in Leporinus Spix, 1829 (Anostomidae).
The eyed epigean form (surface fish) of A. mexicanus is widely distributed in northeastern Mexico and southern Texas, and the eyeless hypogean forms (cavefish) live in some caves inside this extension. 31 Therefore, an ancestral link is possible between the single pair of chromosomes carrying U2 snDNA in A. mexicanus from Mexico and single U2 snDNA pair in other species of Astyanax from North and Central America, as for example, A. jordani studied by Silva et al. 13 that also showed one chromosome pair bearing U2 snDNA clusters.
In summary, the variability in diploid chromosome number in the Astyanax genus is not reflected in the chromosomal organization of the U2 snRNA genes. However, U2 snDNA sites appear to be located on two chromosome pairs with medium size and similar morphologies in almost all Astyanax species. The U2 snDNA cluster stability could be the result of an evolutionary advantage or association with specific DNA segments or particular regions of the genome, which may have facilitated the maintenance of U2 snDNA on two chromosome pairs in South American Astyanax species.
Footnotes
Acknowledgments
The authors are grateful to Dr. Anderson Luis Alves (EMBRAPA) for provision of aquarium species and Dr. Carlos Alexandre Fernandes (UEMS) for provision of laboratory facilities for cytogenetic preparations. This study was supported by CAPES (Coordenadoria de Aperfeiçoamento de Pessoal de Nível Superior).
Disclosure Statement
No competing financial interests exist.