
Abstract
Abstract
To optimize diet particle size for ingestion by zebrafish larvae, Danio rerio, a series of diet selection experiments were carried out using two different length classes: 5-day-old, first feeding larvae with a mean standard length (MLs) of 3.8 mm and 15-day-old larvae with MLs of 5.2 mm. For this purpose, crosslinked protein-walled capsules with five different size classes (≤20, 21–45, 46–75, 76–106, and 107–212 μm) were used in selection experiments. A particle size selection model was then developed after accounting for both rate of loss of capsules in the water column and their ingestion by larvae. Results indicated that concentration of larger particles in the water column decreased rapidly and less than 20% of capsules larger than 75 μm were available for the larvae after 30 min. Zebrafish larvae accepted a wide range of particle sizes, but larvae preferred particles much smaller than the maximum size they could ingest. While first feeding larvae preferred 21–45 μm capsules, 15-day-old larvae preferred capsules in the 47–75 μm size range. A better understanding of the behavior of small-sized food particles in the water column and their acquisition by fish larvae is important to optimize feeding protocols in successful larvae culture.
Introduction
I
Fish larvae have been shown to exhibit preference among food items even when they were offered different size groups of live feed organisms such as rotifers and Artemia nauplii. 3 In larval fish culture, use of optimal diet size is important to increase feeding success, which in turn affects growth and survival. However, despite extensive information on the size preference of cultured and wild fish larvae feeding on live food organisms,4–9 the particle size preference of artificial diets is less studied. Characteristics of artificial diets that affect acceptability by fish larvae such as organoleptic properties, movement, buoyancy, and detectability are often ignored. 2 Dietary size preference is of particular importance to altricial fish larvae due to their specific requirements for small-sized particles during early feeding.
Relative to live feed organisms, the acceptability of inert particles is inherently lower. This is a major bottleneck for the extensive use of microparticulate diets during early stages of altricial larval fish. In larval fish culture, several problems related to the size of artificial diets have been reported. 4 For example, particles with high sinking rates will sink to the bottom of larvae tanks, which in turn will create fouling problems. 10 Small particles, on the contrary, can hardly be detected by fish larvae. 11 Large particles can also cause blockage of the digestive tract. 11 The use of one size class results in inferior growth rates; it was reported that larvae reared on only one size class of prey eventually grow to a size where the metabolic cost of search and capture exceeds the calorific value of the small prey item. 12 The increase in size of prey preferred by fish larvae as they grow has been documented for many species,12–14 and therefore, adjustment of optimal size of dietary particles is needed as fish larvae develop and grow. In addition to particle size, the concentration of particles in the water column is critical and should be adjusted so that encounter rates are maximized. It is suggested that a favorable ratio is maintained by fish larvae between the energy gained by prey ingestion and that spent in capturing the prey to maximize intake for growth.2,15,16
It has been stated that an inert particle is more likely to be ingested by fish larvae if it has the correct size, low sinking rate, and attractive smell and taste. 10 Considering the importance of feeding success for larval fish, the objective of this study was to determine which size range of particles was preferred by zebrafish larvae when they were offered a range of different sizes at similar concentrations. Differential loss of differently sized particles is usually underestimated in studies with inert diets and should be considered when examining particle size preference of fish larvae. This is critical as larger particles have higher sinking rates than those of smaller particles, and thus, larval encounter possibilities in the water column are limited to smaller size particles that stay suspended for longer periods of time. Therefore, in the present study, to determine diet size preference of zebrafish larvae, we first determined the rate of loss of particles in the rearing units. In feeding trials, taking into account of differential sinking rates of different size dietary particles, the larvae were offered different size classes of particles simultaneously at similar concentrations. A model was then developed to determine relationship between concentrations of particles in the water column and inside the larval gut.
The experiments were carried out using zebrafish larvae, Danio rerio, as a model. Zebrafish larvae posses a number of experimental advantages for use in feeding studies. Zebrafish larvae are altricial, transparent, and the gut content can easily be observed under a microscope until 20–25 days old, making them very suitable for feeding experiments. In addition, zebrafish larvae readily accept a wide range of microparticulate diets 17 and can be cultured under laboratory conditions.
Materials and Methods
In the present study, a yeast-based commercial larval food (Microfeast R Plus L-10; Salt Creek, Inc.) was delivered to zebrafish larvae by encapsulation within crosslinked protein-walled capsules (CLPWC) using a method as described. 18 Preliminary feeding experiments with zebrafish larvae fed on Microfeast showed that this diet can be successfully used for first feeding and promotes growth and survival during 10 days after the start of exogenous feeding. Experiments on diet size selection were carried out with CLPWC because these capsules had a rigid, spherical shape that could easily be seen and measured in the stomach. In all experiments, five different size classes (≤20, 21–45, 46–75, 76–106, and 107–212 μm) of CLPWC were used.
In this study, the experiments with CLPWC were carried out using Imhoff cones with a total volume of 1 L. The cones were modified so that total volume was 1.2 L and culture water entered the containers from the bottom and left from the side, near the top, creating an upwelling current. The flow rate was 80 mL/min so that in each container, four turnovers of culture volume per hour were maintained throughout the duration of the experiments. During the experiments, a 10-μm Nitex screen attached to the outlet of each container prevented the loss of capsules larger than 10 μm due to water exchange. No aeration was used in the culture vessels to generate water currents or dispense food.
Rate of loss of CLPWC
A series of experiments were carried out to determine the average rate of loss of five different size classes of CLPWC in Imhoff cones with an upwelling flow rate of 80 mL/min. For each size class, the experiment was carried out in triplicate. At the beginning of experiment for each size, a given weight of CLPWC was added to cones to give initial concentrations of either 50 or 100 capsules/mL for each size. One milliliter of samples of culture water at 15-min intervals was taken and changes in the number of capsules/mL were determined over a period of 60 min for each size class. Samples were taken 15 cm below the surface where zebrafish larvae typically concentrate in greatest numbers.
The general equation used to describe the exponential decrease in the concentration of CLPWC during a period of 60 min and with a flow rate of 80 mL/min was as follows:
where xi(t) = number of capsules of a given size class at time t, in a volume of 1.2 L, Ci = initial concentration of CLPWC/mL, e = 2.7818, α = rate at which particles were lost from suspension, t = times in minutes.
Equation (1) was rearranged so that the sinking rate of CLPWC during 60 min could be calculated from the standardized initial concentration of 10 capsules/mL for each size class:
α, the rate at which particles are lost from suspension, was determined for each size class, by rearrangement of Equation (1) as follows:
where Co = initial concentration, Ct = concentration at time t.
A fitted mean “α” value was calculated for each size class by using experimental data for samples taken at 15-min intervals, over a period of 60 min with a flow rate of 80 mL/min. These fitted mean “α” values for each size class were then used in the analysis of particle size preferences of zebrafish larvae.
Diet size preference of zebrafish larvae
A final set of experiments was carried out to determine the size of capsules zebrafish larvae preferred to ingest when they were either 3.8 mm (5-day-old first feeding larvae) or 5.2 mm in length (15-day-old larvae). For each size class, a total of 300 fish were placed in three cones (100 larvae/cone) and the flow rate was adjusted to 80 mL/min (4 turnovers/h). The experiments were designed so that an average of 2 capsules/mL of each size class was maintained in the water column by adding a predetermined weight of capsules from each size class every 15 min over a period of 1 h to compensate for the rate of loss of capsules. Due to differences in the rate of loss of each size class, different amounts of capsules of each size class were added to each cone. The mean numbers of CLPWC/mg are given in Table 1. Throughout the duration of the experiment, the laboratory was kept under controlled temperature of 28°C. Six, 15 W fluorescent lights provided a mean light intensity of 10 μmol photons s−1 m− (Li-Cor light meter; Model No. Li-185B).
SD, standard deviation.
After a period of 1 h, all the larvae in the cones were sampled and preserved in 70% ethanol after anesthetizing. Preliminary experiments with CLPWC showed that after 12 h in 70% ethanol, there was no shrinkage in the mean standard length of larvae. The larvae were placed on a slide, their guts were opened with a needle, and the gut contents were forced out. The stomach samples of a total of 90 fish (30 fish/cone) were then observed under a microscope and photographed. The sizes of the ingested capsules were then measured by an image analysis software (NIH image) and the mean feeding rate (the mean number of particles ingested by one larva) for each particle size throughout the experiment was determined.
Preliminary experiments with CLPWC showed that after 12 h of suspension in distilled water and 70% ethanol, the increase in diameter of capsules was 5% compared with the initial diameter. Final diameters of CLPWC were corrected for this effect. The particle size preference of zebrafish larvae was determined by comparing the average size distribution of suspended capsules in the water column during 1 h after correcting for different rates of loss of capsules of different size, with the size composition of ingested capsules at the end of 1 h. A mathematical model to determine the particle size preference of zebrafish larvae was developed based on the following assumptions:
(a) the reduction in concentration of particles was due to ingestion by the larvae and to the rate of loss (i.e., gravitational settling, capsules sticking to screened outlets and sides of cones), (b) the larvae's ingestion rate was constant throughout the experiment, (c) the larvae were sampled before CLPWC were digested or egested, and (d) experimental conditions were the same throughout the experiment.
The following equation was used to determine the particle size preference of zebrafish larvae:
where dCi(t)/dt is the rate of decrease in concentration Ci at time t per 1.2 L; −αi is the rate at which particles were lost from suspension for a given size class; Ci is the concentration of a given size of capsule in the container per 1.2 L; and Zi is the constant obtained by the mean feeding rate/hour.
Solving the above equation gives:
where Ci is the concentration of a given size class at time t per 1.2 L and βi is the initial concentration for a given size class per 1.2 L.
For t = 0 to t = 15
−
(time before the first addition of capsules), the equation becomes as follows:
For t = 15+ (time after the first addition of capsules) to t = 30− (time before the second addition of capsules),
where Ci (15) = ci (15 − ) + δi and δi is the amount added every 15 min for a given capsule size class.
As shown in Equations (6) and (7), the average concentration of capsules in each size class in the water column was calculated for a period of 60 min by accounting for the rate of loss of capsules, ingestion by the larvae, and addition of capsules from each size class to compensate for sinking at 15-min intervals. To maintain an average concentration of 2 capsules/mL for each size class throughout the duration of the experiment, a predetermined weight of capsules for each size class was added to containers at 15-min intervals to compensate for losses.
The selection coefficients for zebrafish larvae fed on the five different size classes were then determined by the following equation:
The coefficients obtained for each size class of CLPWC from Equation (8) were then normalized using the following equation:
The % relative selection coefficients for the various size classes of capsules were then compared to each other to determine which size class was preferred by zebrafish larvae either 3.8 or 5.2 mm in length.
Results
Rate of loss of CLPWC
The changes in the mean concentration of different size classes of CLPWC during 60 min of suspension in Imhoff cones under an upwelling flow of 80 mL/min are summarized below in Table 2. There was a dramatic decrease in the concentration of CLPWC under an upwelling flow rate of 80 mL/min at the end of 1 h. Observed data showed that the concentrations of ≤20 and 21–45 μm size capsules after 1 h were 3.0 and 2.8/mL, respectively, indicating that 70%–72% of these particles were not available for fish larvae after 1 h. This decrease in concentration was particularly significant for capsules larger than 75 μm; only 16% of CLPWC were available after 1 h. Rate of capsule loss was considerably higher for particles >75 μm. The loss of CLPWC for both 76–106 and 107–212 μm size classes in the water column was similar with only 14% and 8% being available after 30 min and 1 h, respectively.
Data in Table 2 were used to calculate a fitted mean “α” value to describe the rate of loss of CLPWC for each size class using Equation (2). This value was then used to formulate an equation to describe the mean sinking rate of CLPWC for each size class using Equation (4). The predicted concentrations of CLPWC were similar to those of measured levels. Figure 1A–E gives comparison of predicted values with measured data (Fig. 1A–E). The equation predicted that the concentrations of ≤20 and 21–45 μm size capsules after 1 h should be 4.0 and 3.6/ mL showing a 60% and 64% decrease, respectively, compared with initial concentrations. Final concentration of 46–75 μm size capsules should be 1.2/mL and final concentrations of 76–106 and 107–212 μm size capsules should be both 0.3/ mL, showing a decrease of 97% from initial concentration.
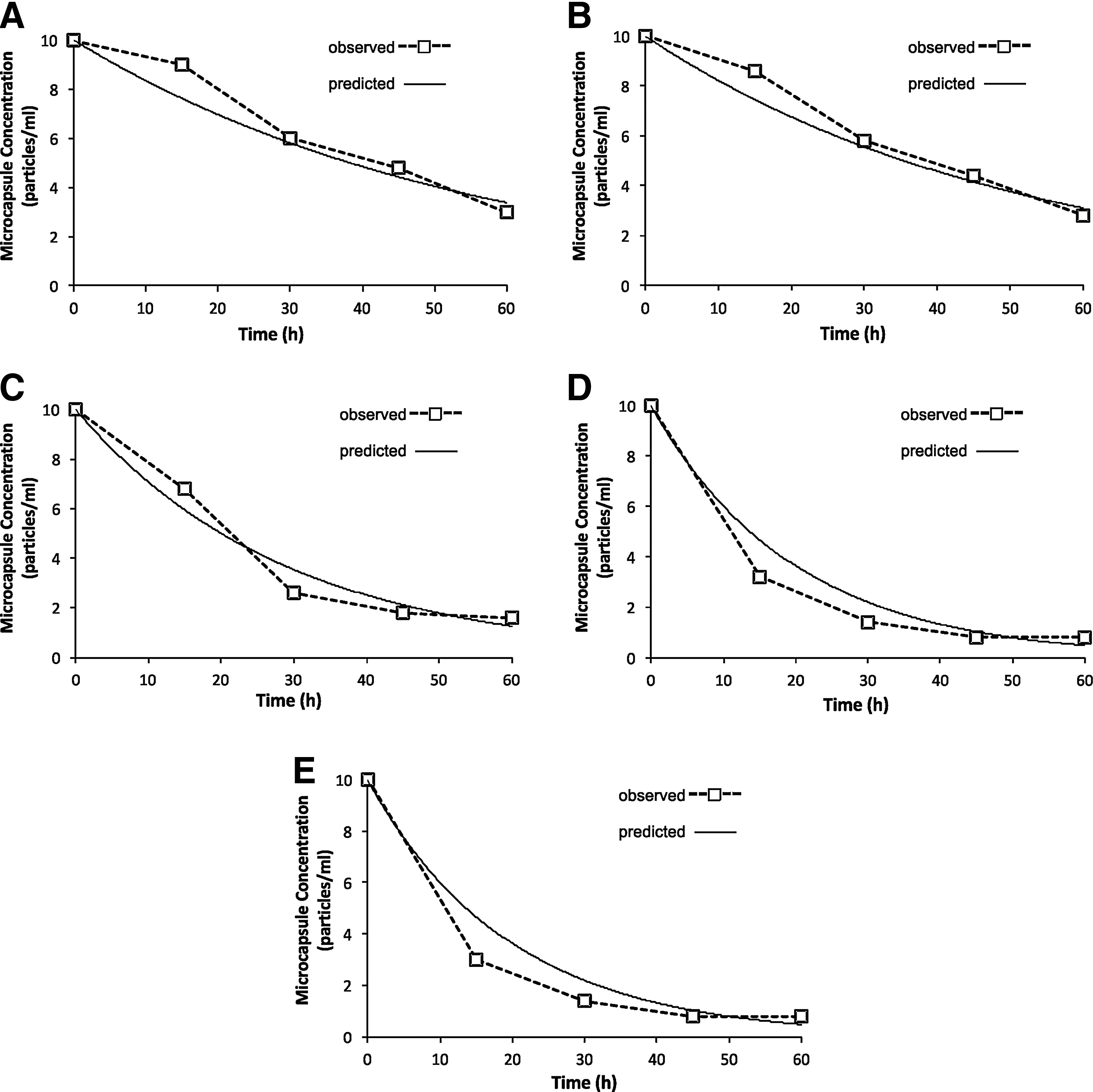
Mean measured (□) and predicted rates (—) of loss of crosslinked protein-walled capsules from the water column.
Diet size preference of zebrafish larvae
In this study, diet size selection of zebrafish larvae was determined by considering the concentrations of different size capsules both in foreguts of larvae and in the water column [Eq. (4)]. Diet size selections are summarized for larvae of 3.8 (5 days old) and 5.2 mm (15 days old) length in Tables 3 and 4, respectively. Preliminary observations showed that while 5-day-old first feeding larvae have a mouth width of 180–200 μm, 5.2 mm larvae have a mouth width of 290–320 μm and are capable of feeding on newly hatched Artemia nauplii >450 μm.
Analysis of the percent relative particle size selection for 3.8 mm zebrafish larvae showed that 21–45 μm capsules were ingested by larvae with a relative selection value of 63.5%, compared with 3.1% and 32% for ≤20 and 46–75 μm capsules, respectively. Only three capsules in the size range of 76–106 μm were observed in the guts of 3.8 mm zebrafish larvae corresponding to a relative selection value of 0.5%. The largest capsule size ingested by 3.8 mm zebrafish larvae had a diameter of 82.3 μm. Larvae were not capable of ingesting 107–212 μm size capsules at first feeding (Table 3).
Zebrafish larvae 5.2 mm in length preferred larger sized particles compared with 3.8 mm larvae. Relative diet size selection for 5.2 mm zebrafish larvae indicated lowest preference (0.7%) for capsules ≤20 μm. On the contrary, 46–75 μm capsules were preferred (61% relative selection) compared with 21–45 μm capsules (27.2%). Although 5.2 mm larvae ingested 107–212 μm capsules, these large capsules were preferred with a relative selection of only 0.9% (Table 4). The largest CLPWC ingested was 160.6 μm in diameter.
Discussion
Determination of rate of diet particle loss from the water column is particularly important when attempting to provide a constant food supply to fish larvae in culture conditions and minimizing loss of particles due to sinking. In this study, an upwelling flow rate of 80 mL/min in the cones partially compensated for the sinking of microparticles in the water column and zebrafish larvae were able to catch and ingest various size particles without difficulty. Inside the conical-shaped containers, the flow rate decreased from the bottom to the top as a factor of increasing cross-sectional area. The differential flow rate also provided a habitable zone 15 cm below the surface of the cones where the larvae were observed to aggregate and started feeding and capture behavior. Flow rates higher than 80 mL/min were not tested due to observed detrimental effects such as too much turbulence and agitation in the water column. Despite a flow rate of 80 mL/min, concentration of larger particles in the water column decreased rapidly and less than 20% of capsules larger than 75 μm were available for the larvae after 30 min indicating their lower availability as food particles for fish larvae.
In addition to increased suspension time of dietary particles, upwelling flow within the containers created turbulence that may have positively affected encounter rates of microparticles and thus acceptance by first feeding larvae. There are numerous studies describing the increase in encounter rates between planktonic predators and prey under microscale turbulence conditions although this theory has not been proven empirically. 19 With younger larvae, for example, it was suggested that slower swimming speeds and shorter perceptive radii could reduce pursuit success under high turbulence conditions compared with older larvae, in which faster swimming speeds and larger perceptive radii act to diminish the detrimental effects of high turbulence. 19 Therefore, optimization of flow rate is particularly important for altricial larvae to help them capture food particles easily when they are fed with inert diets.
The observed feeding activity of CLPWC by 5- and 15-day-old larvae indicated that at first feeding and onward, zebrafish are equipped with the necessary anatomical and physiological features required for feed acquisition. CLPWC were readily accepted by 5-day-old zebrafish larvae, suggesting that the taste of microparticles was attractive. This finding is in accordance with the timing of taste bud development in zebrafish larvae that coincides with the onset of feeding. 20 In this study, from the start of exogenous feeding, zebrafish larvae accepted a wide range of particle sizes, but larvae preferred particles much smaller than the maximum size they could ingest. It was suggested that the lower limit of acceptable prey size appears to be partly determined by the metabolic requirements of the larvae, while the upper limit is determined by the mouth width, which increases in proportion to larval length. 21
This study indicated that 3.8 mm zebrafish larvae preferred 21–45 μm capsules. The particle size/mouth width ratio of 3.8 mm zebrafish larvae corresponds to about 15.7%–18.3% considering a mean particle size of 33 μm for 21–45 μm capsules. On the contrary, although 3.8 mm zebrafish larvae were capable of ingesting capsules with a particle size/mouth width ratio of up to 43.3%–50.5%, the preference of capsules >76 μm by zebrafish larvae was only 0.5%. Similarly, Fernández-Díaz et al. 4 reported that Sparus aurata larvae with a 4 mm total length preferred 25–50 μm particles. Our data indicated that the size of capsules ingested by zebrafish larvae increased; 5.2 mm larvae selected 46–75 μm size capsules. Furthermore, it was determined that the preference for 76–106 μm capsules by 5.2 mm larvae was 20 times greater than that for 3.8 mm larvae, suggesting that diet size preference of larvae is directly proportional to larval length, as reported in Refs.,4,22–24 for other larval fish species using either live or inert foods. This finding is consistent with those reported earlier and indicates improved feeding performance as the larvae grow.
In the present study, while the upper size limit of capsules that could be ingested by first feeding zebrafish larvae was determined to be around 76–106 μm, 21–45 μm particles had higher preference. Similarly, although 15-day-old larvae could ingest capsules in 107–212 μm size class, larvae preferred 46–75 μm particles. In feeding trials, since the encounter possibilities of different size particles were similar, these results suggest that zebrafish larvae can ingest particles much larger than the preferred size when they encounter a wide size range of particles. In contrast, although larger size capsules represented only a small proportion of the total ingested capsules, in terms of weight, they accounted for a considerable proportion of the total ingested biomass in the gut. For example, while 76–106 and 106–212 μm capsules represented 11.55% and 1.03% of the total ingested capsules, respectively, these two size classes represented 44.73% of the total ingested biomass combined (26.61% and 18.12%, respectively). Nevertheless, considering higher rates of loss of dietary particles >106 μm from the water column, a relatively narrow size range of dietary particles between 21 and 75 μm should be used for first feeding zebrafish larvae to increase acceptability and minimize waste of dietary particles.
The chance of any organism to be a potential prey depends on the physical capacity of predatory larvae to catch and eat it. 4 It has been reported that the biomechanics of larval feeding is similar to that of adult fish; while attacking/swimming toward their prey, larval fish create a suction by rapidly opening their mouth that drives the prey with a volume of water into the mouth.25,26 It has been suggested that since smaller mouths produce weaker suction flows, fish larvae are more successful in capturing inert particles during early stages and less successful in capturing live feed organisms. 26 This may also explain higher preference of smaller size capsules that would require weaker suction flows for capturing and the increase in size of capsules preferred by larger size zebrafish larvae. Similarly, first feeding gilthead seabream, Sparus aurata, larvae could ingest Artemia salina nauplii, but larvae started positively selecting these organisms only after the mouth size increased to allow ingestion even though the ratio of prey size/mouth width was not optimal. 4 For fish larvae feeding on live feed organisms, the prey size/mouth width ratio has been reported as 25%–50%. 27
In the present study, although zebrafish larvae of 5.2 mm ingested a wide range of particles, the maximum size range of the capsules ingested was smaller than the size of Artemia nauplii, the usual food of cultured larvae of this size and the ratios of the largest capsule sizes ingested/mouth width were found to range between 40% and 55%. In addition to mouth gape, esophagus diameter is considered as a critical factor in particle size preference.2,7,28 In this study, the prey size/mouth width ratio and the largest capsule sizes recovered from the stomach may indicate the physiological upper limit of particle sizes that can go through the esophagus of 3.8 and 5.2 mm zebrafish larvae and point out the importance of compressibility or deformability of inert particles. Fernández-Díaz et al., 4 using particles of different hardness, showed that different sizes of gilthead sea bream, Sparus aurata, ingested larger particle sizes when the particles were softer. Although CLPWC used in the present study are not deformable following hydration, deformability of diet particles may help larvae ingest larger size particles and indicates the importance of structural characteristics of the food item.
In the present study, the particle size preference of zebra fish larvae was determined in the presence of a wide range of spherical CLPWC. An ad hoc procedure was used to determine the mean rate of loss of different sizes of microcapsules from the water column after examining experimental data. The overall goal was to compensate for the rate of loss of capsules in the containers by regular additions to maintain similar concentrations of different size capsules during diet size preference experiments. Sinking of capsules was the major contribution to the rate of loss of capsules from suspension. In commercial setups without the presence of upwelling currents, the high sinking rates of capsules larger than 75 μm can result in decreased ingestion of these capsules by larvae, which may require more frequent feedings and can impair growth. In addition, higher sinking rates of larger size dietary particles will increase feed waste, which, in turn, will exacerbate water quality-related problems. Therefore, a better understanding of the acquisition of food particles by fish larvae is important and this information can be used to optimize feeding protocols for the rearing of commercially important food fish. Consideration of biological limitations as well as physical characteristics of artificial diets is critical. From this point of view, efforts toward increasing buoyancy and deformability of microparticles are important to prolong their suspension and to increase the preference and acceptability of inert particles.
Footnotes
Acknowledgments
This study was supported by Saltonstall-Kennedy Program NOAA Award 66FD 0107. The authors thank Dr. Mehmet Emin Alpay for his contributions to the article.
Disclosure Statement
No competing financial interests exist.