
Abstract
Abstract
There has been a rapid expansion in use of transgenic technologies in zebrafish. We report a novel example of transinteractions of genetic elements, or transvection. This interaction led to a novel expression pattern and illustrates a precautionary example regarding use of transgenes in zebrafish.
R
We generated a Tol2-based transgenic line expressing the human inwardly rectifying potassium channel gene kir2.1 (KCNJ2), Tg(UAS:hkir2.1-mCherry). kir2.1 has been used in prior studies including in zebrafish to hyperpolarize and silence neurons. Interestingly, when we crossed this line to a double transgenic line we have used previously, the triple-transgenic Tg(foxP2.A.2:Gal4), Tg(UAS:GFP), and Tg(UAS:kir2.1-mCherry) embryos had GFP expression throughout the brain and not just in the expected subset of foxP2 neurons (Fig. 1A–D) (“green brain” phenotype). There was no expansion of endogenous foxP2 expression (in situ data, not shown). Intriguingly, UAS:GFP and UAS:kir2.1-mCherry expression did not match (Fig. 1B–D); neither transgene's expression matched the normal expression pattern of foxP2.A.2. 1 This was unexpected, given that both UAS:GFP and UAS:kir2.1-mCherry transgenes share the same UAS enhancer and minimal core promoter (MCP) sequences. Using a GCaMP5 reporter, inhibition of electrical activity matched the altered expression pattern of UAS:hkir2.1-mCherry labeling. For experiments we used the widely used Gal4-VP16413–470 and 10x UAS constructs. Although we did not formally test whether any of the transgenes had more than one insertion, we have used these lines over multiple generations and not noted any variegation of expression, suggesting that they are either single copy; or if multiple copies, are closely linked.
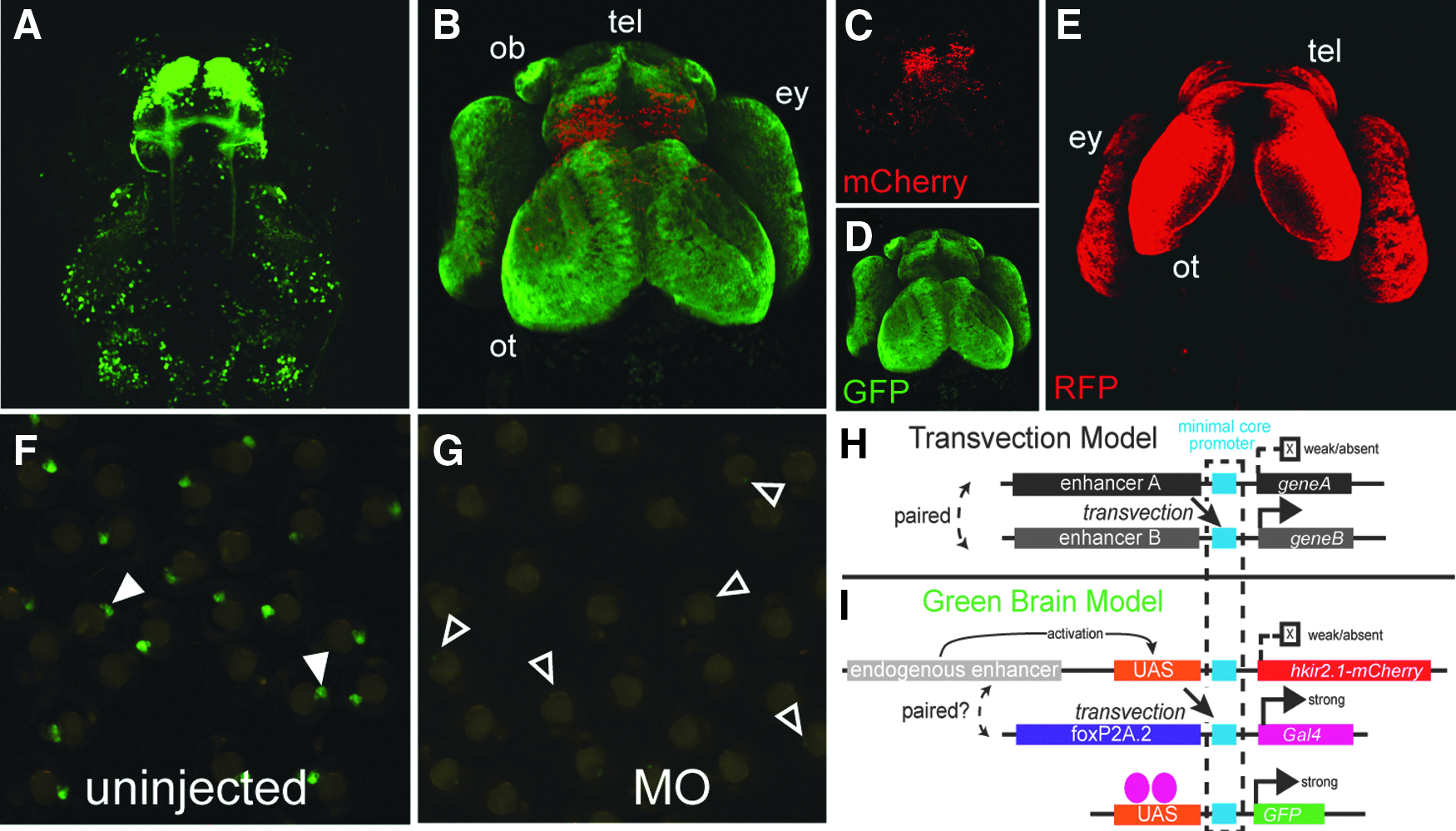
We were unable to recapitulate the green brain using chemical or genetic mechanisms to disrupt neuronal activity, including glutamate receptor antagonists (AP5, CNQX), a sodium channel inhibitor (MS-222), increased extracellular K+, or crossing to tetanus toxin Tg(UAS:TeTx-CFP) fish. The potassium channel blocker chloroquine that specifically inhibits kir2.1 did not suppress the green brain phenotype. We did not observe a similar expansion in GFP labeling using other Gal4 driver lines. However, a Gal4 morpholino prevented the green brain phenotype (Fig. 1F, G). The expansion of GFP segregated with only the joint presence of the foxP2.A.2:Gal4 and UAS:kir2.1-mCherry transgenes, and we could create a “red brain” phenotype in foxP2.A.2:Gal4, UAS:RFP, and UAS:kir2.1-mCherry embryos (Fig. 1E). Outcrosses demonstrated that the transgenes were not at the same or a nearby genomic locus. Thus, the green brain is Gal4/UAS dependent, is dependent on two specific transgenes (foxP2.A.2:Gal4 and UAS:hkir2.1:mCherry), but is distinct from the normal expression pattern of foxP2.A.2.
This phenomenon is consistent with transvection, the trans interaction of genetic elements. 2 Transvection can involve cis and/or trans interactions of genetic elements, often between paired (allelic) insertions (Fig. 1H). 3 Transvection can occur in trans when an enhancer drives expression by binding and activating expression at the paired allele. For example, in Drosophila, inserting two transgenes with different enhancers but a shared MCP into the same genomic location, including for UAS-driven transgenes, is sufficient to elicit transvection. 4
Many transgenes in the zebrafish community use the same MCP containing the viral E1b TATA box fused to the carp β-actin 5′ UTR. We hypothesize that the green brain phenotype is because of transvection between the UAS:hKir2.1:mCherry and foxP2.A2:Gal4 transgenes, containing the same MCP, which by chance have inserted into interacting positions in the genome. The UAS:hKir2.1:mCherry may have inserted into a genomic region where an endogenous cis-regulatory element constrains its UAS expression, but leads to trans-driven expression of foxP2.A.2:Gal4, and the subsequent expansion of GFP (or any other UAS-dependent gene) (Fig. 1I). Our findings are an important reminder to include controls for expected transgene expression patterns because of the potential for transvection interactions when using multiple transgenes in zebrafish.
Footnotes
Disclosure Statement
No competing financial interests exist.