
Abstract
Abstract
In the Pantanal of Mato Grosso do Sul, electric fish (Gymnotus spp.) are the primary source of live bait, accounting for more than three-quarters of total sales. Based on chromosomal and molecular markers, the present study attempted to identify the Gymnotus species used as bait in the region of Corumbá, Mato Grosso do Sul, Brazil. Three species were detected, based on their distinct karyotypes: G. paraguensis (2n = 54), G. sylvius (2n = 40), and G. pantanal (2n = 39–40, X1X2Y/X1X1X2X2), with no evidence being found of interspecific hybrids. All three species presented a single nucleolar organizer regions (NOR) (heterochromatin CMA3+/DAPI−) and pericentromeric heterochromatin in almost all chromosomes, with a few distal and/or interstitial blocks. G. sylvius and G. pantanal had one and two pairs of chromosomes with 5S rDNA sites, respectively, while G. paraguensis had 17 chromosome pairs with these markers. The three species formed well-defined clusters in the DNA barcoding analysis. The integrated analysis of the cytogenetic and DNA barcoding data confirmed that the diversity of Gymnotus species exploited as live bait in the study region has been underestimated. These findings indicate that the markers analyzed represent valuable tools for the conservation and fishery management of the Gymnotus stocks exploited.
Introduction
B
The fish fauna of the Pantanal is also very diverse, with 269 species being identified up to now. 4 One notable group is the order Gymnotiformes, which presents a number of unique morphological characteristics in comparison with other Neotropical fish. A prominent gymnotiform family is the Gymnotidae, which includes Gymnotus Linnaeus, 1758 and Electrophorus Gill, 1864. 5 These fish are able to emit and detect electrical signals for communication and active electric location, which is the most studied aspect of the biology of this group.6–8
Many fish species are not easily distinguishable on the basis of external morphological characteristics, and in Gymnotus, reliable species identification may require alternative techniques such as cytogenetics, which suggests the existence of cryptic species in this diverse genus. In the Pantanal, however, bait fishermen are unable to use cytogenetic techniques, and rely on the observation of specific external characteristics to differentiate the fish for capture, management, and marketing. The feature most commonly used for identification is the coloration pattern, with two principal groups, black and yellow. During observations in the field, however, it was possible to confirm that more than one species may present the same coloration pattern.
The cytogenetics of Gymnotus have been relatively well studied, 9 and recent studies have contributed to the increase in the number of recognized species from 9 10 to 37. 11 This research has revealed considerable karyotype diversity, with diploid numbers ranging from 2n = 34 in G. capanema 11 to 2n = 54 in G. carapo, G. mamiraua, G. paraguensis, and G. inaequilabiatus.9,12–14 A multiple sex chromosome system X1X1X2X2/X1X2Y was also observed in G. pantanal, in which females have 40 chromosomes and males, 39 (Table 1).13,15
m/sm, metacentric/submetacentric; st/a, subtelocentric/acrocentric.
Given these difficulties, recently developed molecular techniques provide an important way of increasing the reliability of species identification. 16 In particular, DNA barcoding constitutes a rapid and highly effective tool for the delimitation of species. 17 This approach, which has shown to be effective in a wide range of organisms,17–26 uses a small segment of the mitochondrial cytochrome oxidase I (COI) gene to identify known species and reveal new species.
In the Pantanal, electric fish are the principal source of commercial live bait for sport fishing, contributing more than three-quarters of total sales in the region. 27 This live bait is popular for the capture of the most prized local game fish. State resolution 22, published on August 25, 2011 by the Mato Grosso do Sul Department of Environment, Cites, Planning, Science and Technology (SEMAC-MS) registers the marketing of only two species, G. inaequilabiatus and G. paraguensis, as live bait. However, a preliminary analysis of microsatellite markers indicated that at least three Gymnotus species are marketed as live bait in the region of Corumbá. 28 Considering these evidence, the present study characterized chromosomal markers and sequences of the mitochondrial cytochrome C oxidase I gene (the barcode region) in specimens of Gymnotus captured for sale as live bait in the region of Corumbá, Mato Grosso do Sul, on the middle Paraguay River in Brazil.
Materials and Methods
Cytogenetic studies
A total of 78 specimens representing the species G. paraguensis (N = 39), G. pantanal (N = 29) and G. sylvius (N = 10), were obtained from live bait suppliers near Corumbá (Mato Grosso do Sul, Brazil) in the Paraguay River basin in 2011 and 2013. The specimens were obtained from three locations—(1) the São Lourenço River (17°50′53.6″ S, 57°23′30.3″ W), (2) downstream from the confluence of the São Lourenço and Paraguay rivers (17°54′48.83″ S, 57°27′39.13″ W), and (3) the Paraguay River upstream from its confluence with the São Lourenço River (17°51′37.9″ S, 57°31′11.1″ W). The voucher specimens were deposited in the fish collection of the Institute of Biosciences at the Federal University of Mato Grosso, Brazil, under serial numbers CPUFMT3205–3208 and CPUFMT4166.
This study was carried out in strict accordance with the recommendations provided in the Guide for the Care and Use of Laboratory Animals. When necessary, the specimens were first anesthetized and then euthanized using an overdose of clove oil, 29 to avoid any suffering.
The chromosomal preparations were obtained directly from kidney cells. 30 The nucleolar organizer regions were detected with silver nitrate impregnation (AgNOR) 31 and the samples were C-banded to determine the distribution of the heterochromatin. 32 The CG-rich regions were stained with chromomycin A3 (CMA3) 33 and the AT-rich regions were stained with DAPI (4,6-diamino-2-phenylindole) fluorochrome. 34
For the Fluorescence in situ hybridization technique (FISH), 35 the 18S rDNA and 5S rDNA probes were labeled according to the manufacturer's instructions (Biotin and Dig 11 Nick Translation mix, Roche, respectively). Chromosomes were classified according to their arm ratios, 36 with adjustments for fish cytogenetics.37–39
Molecular analysis
Molecular DNA barcoding data were obtained from 28 specimens representing G. paraguensis (N = 18), G. pantanal (N = 5), and G. sylvius (N = 5). For this, the genomic DNA was extracted from muscle or liver tissue. For the specimens with nonspecific tissue samples for molecular studies, we used samples of the cell suspensions prepared for the chromosome studies. This material was suspended in 1.5 mL tubes before removing a small aliquot, of ∼100 μL. This aliquot was centrifuged (10,000 rpm for 10 min) and dried at 37°C. After the material was completely dried, the genomic DNA was extracted using an adapted salt-extraction. 40 After adding the extraction solution and mixing, the samples were incubated at 55°C for no >1 h, to prevent the degradation of the DNA. Immediately after this precipitation step, the samples were washed three or four times in 70% ethanol.
The regions of the COI (∼700 bps), 18S and 5S genes were amplified by polymerase chain reaction (PCR) using the respective primers: COI FishF1 and COI FishR1, 19 18S NS1 and 18S NS2, 41 and 5S A and 5S B. 42 All the amplification reactions were run in a Biocycler (Eppendorf), with a denaturation temperature for COI of 94°C for 5 min followed by 30 cycles of 94°C for 45 s, 52°C for 1 min, 72°C for 1 min, and a final extension of 5 min at 72°C (COI). For 18S, the denaturation temperature was 94°C for 5 min, followed by 35 cycles of 94°C for 45 s, 55°C for 1 min, 72°C for 5 min, and a final extension at 72°C for 5 min, and for the 5S, denaturation at 94°C for 5 min, followed by 35 cycles of 94°C for 45 s, 60°C for 1 min, 72°C for 1 min, and final extension at 72°C for 5 min.
The PCR products of the COI gene were sequenced using the Sanger et al. 43 technique and are available for comparisons in the National Center for Biotechnology Information (GenBank, www.ncbi.nlm.nih.gov), under accession numbers KX389650–KX389669. The 18S and 5S PCR amplified probes were used in FISH (fluorescence in situ hybridization) protocols.
The BioEdit 44 software was used to analyze the quality of the mtDNA sequence, and the Clustal W alignment tool was used to align the multiple sequences. The partial COI sequences were analyzed using the neighbor-joining method under the Kimura 2-parameter model (K2P) 45 in the MEGA 6.0 program, 46 with the support being evaluated by1000 bootstrap replicates, with the mean intra- and inter-specific distances also being determined. Partial sequences of the COI gene of G. inaequilabiatus and Electrophorus electricus were used as outgroups (accession numbers KU289044.1 and AP011978.1).
Results
Overall, 39 of the 78 samples analyzed presented a diploid number of 2n = 54 chromosomes (50m-sm + 4st-a), while 10 specimens were 2n = 40 (36m-sm + 4st-a), and 29 were 2n = 39/40 (15m-sm + 24st for males and 14m-sm + 26st it for females). The karyotypes are consistent with those recorded previously for G. paraguensis (Fig. 1a), 47 G. sylvius (Fig. 1b),13,47 and G. pantanal (Fig. 1c).13,15

Karyotypes of female and male Gymnotus paraguensis
The AgNORs were observed in the interstitial portion of the short arm of metacentric (m) pair 1 in G. paraguensis, on the short arm of subtelocentric (st) pair 20 in G. sylvius, and on the short arm of subtelocentric pair 16 in G. pantanal (Fig. 2a, b, c—box).

C-banded karyotypes of G. paraguensis
In G. paraguensis, the C-banding revealed the occurrence of heterochromatic blocks in the pericentromeric region of all chromosomes, in the interstitial region, more proximal to the centromere on the long arm of pair 7 and at the end of the short arm of chromosome pair 1, that is, the AgNOR sites (Fig. 2a). In G. sylvius, pericentromeric heterochromatin was found on several chromosome pairs (except pairs 3, 6, 15, 16, and 17), in the interstitial region of the long arm of pair 18, and on the short arm of pairs 10, 11, and 20, that is, the AgNOR sites (Fig. 2b). In G. pantanal, the heterochromatin was found near the centromeric region in all chromosomes and on the short arm of pair 16, the AgNOR sites (Fig. 2c).
The FISH with 18S rDNA probes confirmed the results obtained with silver nitrate staining (Fig. 3a, c, e), while hybridization with the 5S rDNA probes revealed a number of sites for this sequence, with ∼17 chromosome pairs in G. paraguensis (Fig. 3b), in the pericentromeric region of pair 4 in G. sylvius (Fig. 3d) and pairs 3 and 5 in G. pantanal (Fig. 3f). No preference for any segment in the case of DAPI staining, with the exception of secondary constrictions, which appear as darker portions, that is, negative DAPI (Fig. 4a, c, e). In addition, the CMA3 staining revealed the brightest heterochromatin blocks (positive CMA3) in the proximal region of the short arm of chromosome pair 1 in G. paraguensis (Fig. 4b), on the short arm of pair 20 in G. sylvius (Fig. 4d), and on the short arm of pair 16 in G. pantanal, corresponding specifically to the AgNOR sites (Fig. 4f).
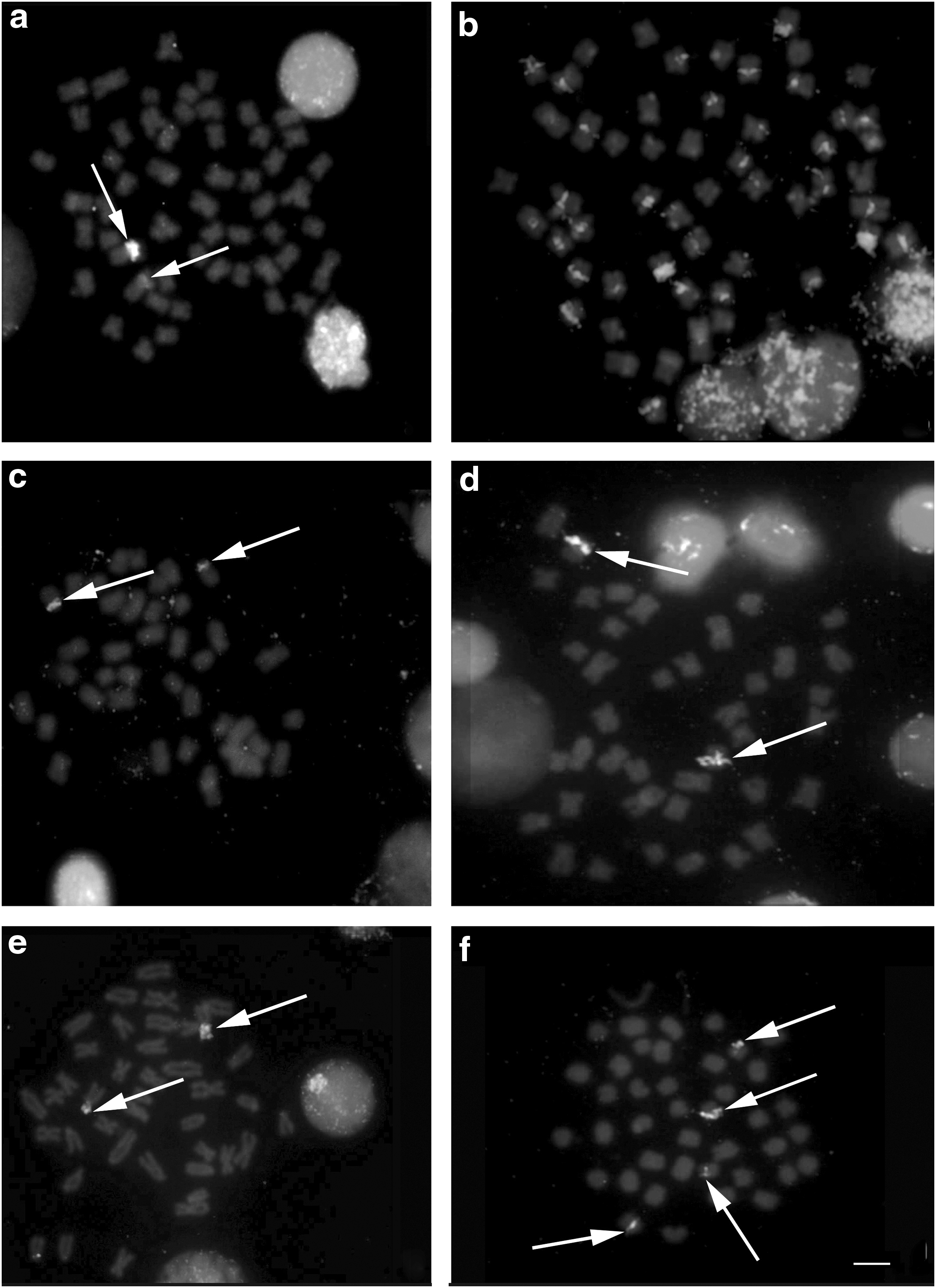
Distribution pattern of the 18S and 5S rDNA sites in G. paraguensis
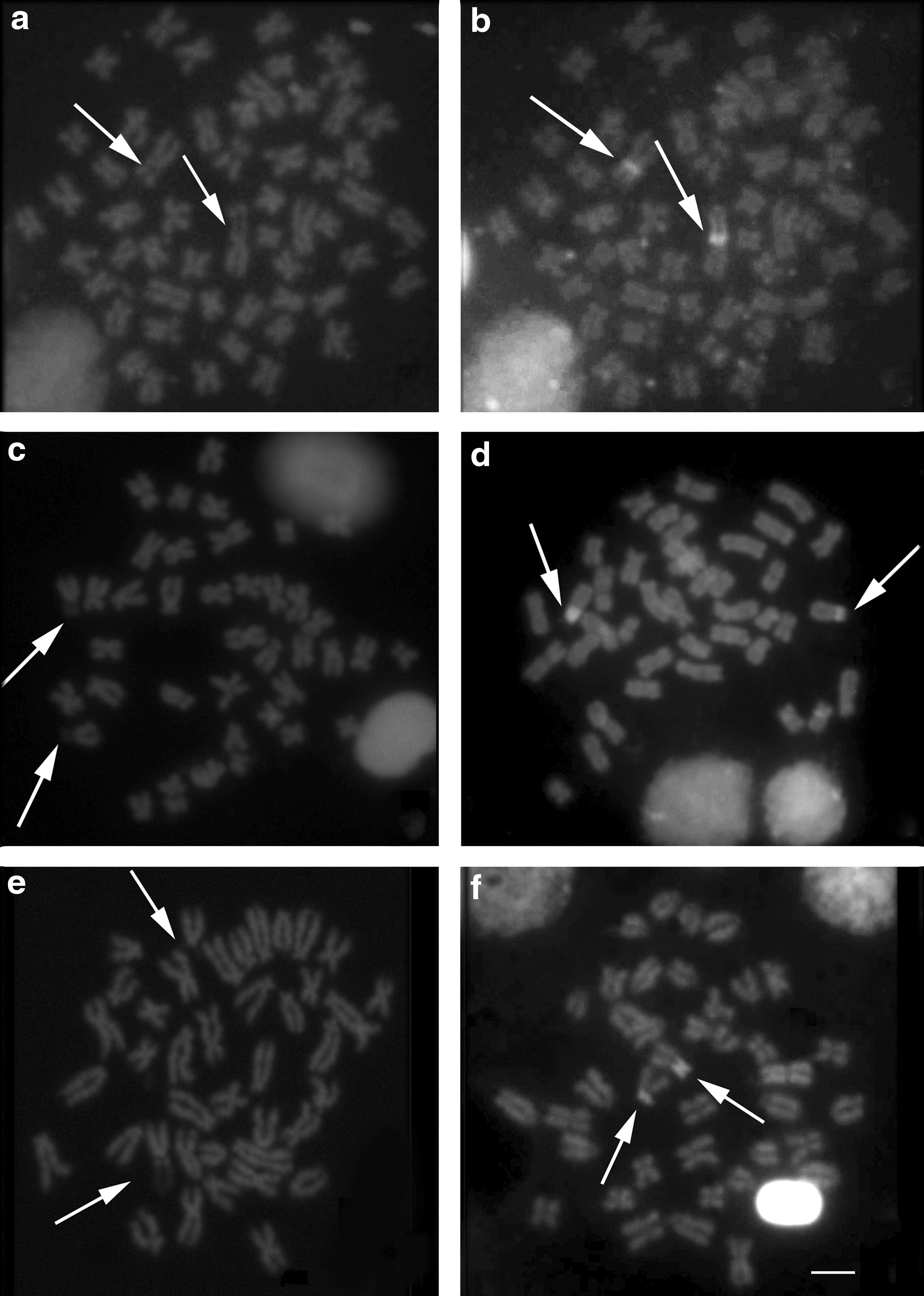
Metaphases obtained by staining with DAPI and CMA3 in G. paraguensis
The DNA barcoding data of the 28 specimens—G. sylvius (n = 5), G. pantanal (n = 5), and G. paraguensis (n = 18)—revealed three distinct groups, as observed in the dendrogram (Fig. 5). The mean intraspecific difference was 0.63% ± 0.22% in G. sylvius and 0.39% ± 0.14% in G. paraguensis, while no intraspecific variation was observed in G. pantanal. The interspecific genetic distance among G. sylvius, G. pantanal, and G. paraguensis were high, ranging from an average of 6.13% to 8.75% (Table 2).
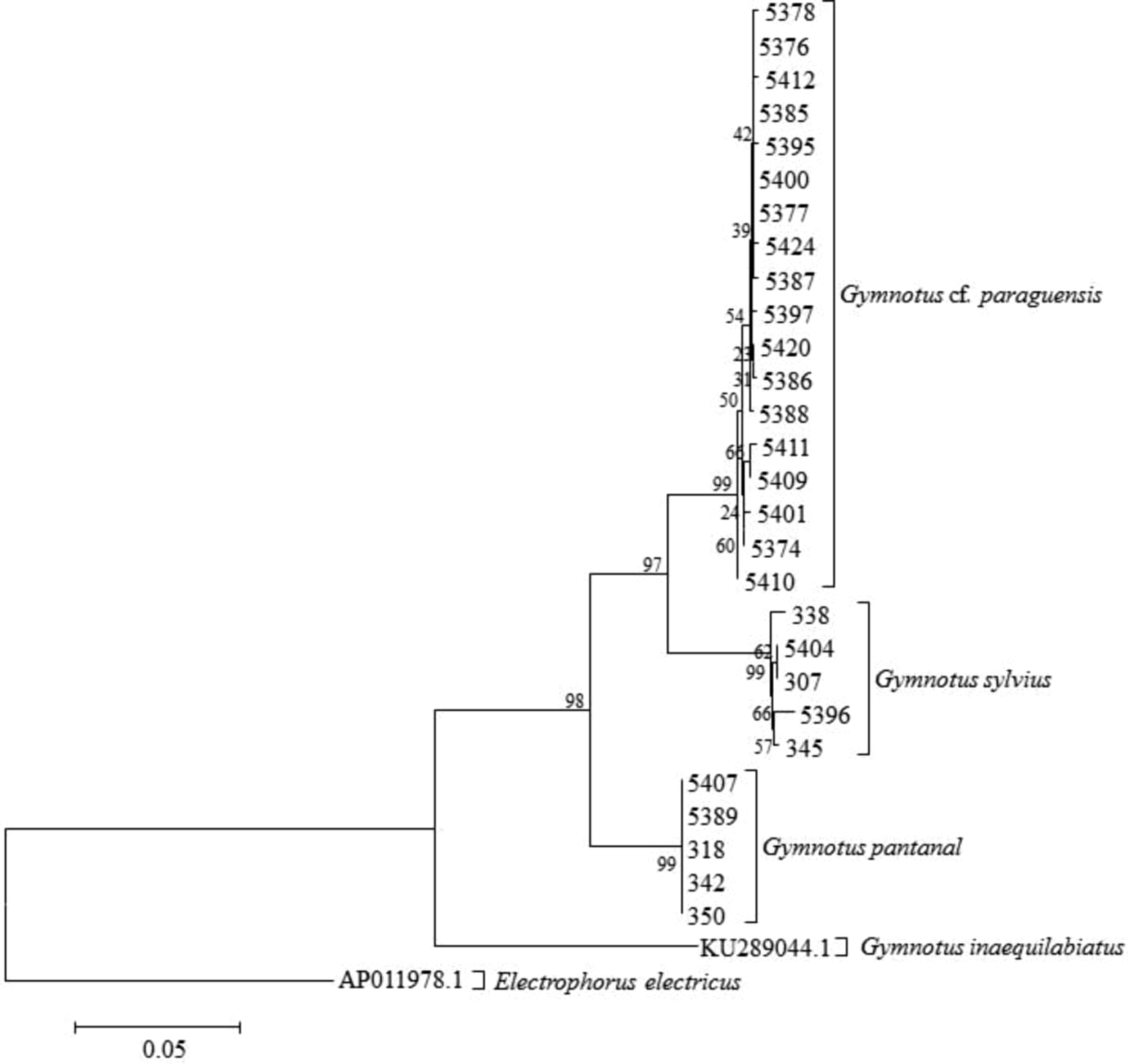
Dendogram obtained by the NJ based on the K2P distances among 28 specimens identified as G. paraguensis, G. sylvius, and G. pantanal collected on the middle Paraguay River. K2P, Kimura 2-parameter model; NJ, neighbor-joining.
The values in the diagonal are the intraspecific distances (a).
K2P, Kimura 2-parameter model.
Discussion
In Gymnotus, the diploid number and karyotype formula vary considerably among species. The results of this study are consistent with the published data,9,11–14,47–51 indicating the existence of species-specific karyotypes. Thus, karyotypic data have been shown to be important diagnostic tools for taxonomic studies. 13 This considerable interspecific karyotype diversity may be related to the limited dispersal of individuals, resulting in the formation of many isolated populations, given the preference of these fish for lentic waters.14,52
The species of this genus typically engage in short migrations,52,53 inhabiting a range of different environments, varying from slow-flowing rivers to floodplain systems. During the rainy season, individuals may disperse passively, favoring gene flow among different stocks, although the existence of these local populations may also provide suitable conditions for chromosomal changes that may eventually become fixed, resulting in the emergence of different Gymnotus species. However, despite these interspecific differences in karyotypes, it is possible to assume that environmental pressures have not played any important role in the morphological differentiation of the species, which are typically indistinguishable under macro-analysis.
The basic karyotypes observed in the different Gymnotus species allow for the reliable differentiation of species, especially given the absence of hybrids, as observed in this study. However, some groups that have similar diploid numbers may be indistinguishable when only conventional karyotypes are analyzed. In this context, more refined cytogenetic analyses, such as the definition of the nucleolus organizer regions, revealed a single AgNOR system in the three study species, confirming the results of previous studies.12–14,48,51,54 Interspecific and intraspecific polymorphisms in the AgNORs are known from a number of fish groups, especially in terms of the size and number of active sites and, at least for the species studied, these variations are not taxonomically informative, and were thus irrelevant to the aims of this study.
The data on the 5S segments in G. pantanal and G. sylvius were similar to those obtained by other authors.50,55 The restriction of the 5S rDNA sites to only one chromosome pair, as observed in G. sylvius, was expected, given that the 5S ribosomal gene generally corresponds to a highly conserved region of the genome. 56 Even with four 5S rDNA markers, the pattern observed in G. pantanal was also conserved, given that its interstitial location can be found in unrelated eukaryote taxa. 57 The distribution of the interstitial 5S rRNA gene may represent an advance in the organization of these genes in the vertebrate genome. 58
Moreover, the distribution pattern of the 5S rDNA sequences in G. paraguensis may be related to structural rearrangements of the karyotype in this species with the involvement of Tc1-like. 50 In the species G. inaequilabiatus, G. cf. carapo and G. pantherinus, the spread of this genetic sequence has been facilitated by breaks in the heterochromatin regions. 14 Transposable elements of retro transposons (Rex3) are associated with the distribution of 5S rDNA in Erythrinus. 59 However, some studies have shown that transposable elements are important structural components of the heterochromatic regions. These sites appear to have played an important role in the evolution of the fish genome. 60 The existence of pseudogenes may also account for the observed variation, 61 given that the heterochromatin may also present inactive sequences, similar to those observed in the 5S rDNA, as observed in Centropyge aurantonotus. 62
The distribution of the constitutive heterochromatin, including the centromeric and pericentromeric regions of the study species, is common in other gymnotiforms. 51 Of the heterochromatic segments observed in this study, only the heterochromatin associated with the AgNORs appears as CMA3+ segments. Fluorescent CMA3+ regions associated with AgNOR sites are common in some species of Neotropical fish, 63 such as G. mamiraua and G. cf. carapo, 9 and this pattern appears to be common in most Neotropical species karyotyped with this fluorochrome, such as Astyanax spp., 64 cichlids, 65 and Hoplias malabaricus. 66 Despite its importance for the characterization of some chromosomal segments, this marker did not provide any useful information for the identification of the species analyzed in this study. Similar situations were observed after DAPI coloration that was ineffective for a better characterization of the chromosomes of these species. Cytogenetic studies identified well-defined karyotypes, but were unable to characterize a possible fourth species in the G. paraguensis group. In this context, the DNA barcode was included to provide a better diagnosis of the number of Gymnotus explored as live baits in the Pantanal region, since this tool has been successfully used in different animal groups, especially fish.23,26,67–70
In this study, the DNA barcode discriminated three units, G. paraguensis, G. sylvius, and G. pantanal, with mean intraspecific divergence 16 times lower than that found between congeneric species. Similarly in marine fish, 19 mean values of 0.39% and 9.93% were obtained for intra- and inter-specific comparisons, respectively, reinforcing it potential for the discrimination of species. DNA barcoding on freshwater Canadian fishes showed mean conspecific divergence of 0.2% and congeneric divergence of 8.37%, permitting the discrimination of 93% of the species. 24 A recent study of freshwater fish from Cuba 71 returned mean values of 0.6% (conspecific) and 9.1% (congeneric), with 96% of the species being discriminated. Other studies recorded mean values of 0.3% (conspecific) and 8.4% (congeneric), discriminating 97.5% of the species. 72 All these studies reinforce the present findings on Gymnotus.
However, a mean interspecific diversity of 10% may be due to the small number of species analyzed in the genus. 26 In this case, the analysis of dominant species, with at least five specimens per species from distinct geographical areas will tend to raise mean intraspecific differences.
The findings of this study, which identified at least three species of Gymnotus being used as live bait, provides important insights for the definition of conservation strategies for the management of the fishery resources used in professional fishing and the regulation of the trade in these fish, especially with regard to overexploitation and the monitoring of congeneric species introduced by humans. A review of the existing legislation will nevertheless be needed, as well as the development of a new management plan for the fish used as live bait in the Pantanal of Mato Grosso do Sul, Brazil, given that the local authorities (SEMAC-MS: Resolution No. 22 of August 25, 2011) recognized the marketing of only two species, G. inaequilabiatus and G. paraguensis. This study has updated this publication by adding at least two more species, G. pantanal and G. sylvius, and questioning the local occurrence of G. inaequilabiatus in the local stocks.
Footnotes
Acknowledgments
We are grateful to Dr. Claudio de Oliveira for his invaluable scientific collaboration, the Brazilian National Research Council (CNPq—case number 477057/2009-8), the Mato Grosso State Research Foundation (FAPEMAT—case number 453511/2009 and 841.147/2009 PRONEX), EMBRAPA Pantanal, and CAPES. The collection of samples was authorized by the competent environmental authorities: SEMAC—MS and Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis (IBAMA—license 12174-2).
Disclosure Statement
No competing financial interests exist.