
Abstract
Abstract
Heart rate (HR) is a periodic activity that is variable over time due to intrinsic cardiac factors and extrinsic neural control, largely by the autonomic nervous system. Heart rate variability (HRV) is analyzed by measuring consecutive beat-to-beat intervals. This variability can contain information about the factors regulating cardiac activity under normal and pathological conditions, but the information obtained from such analyses is not yet fully understood. In this article, HRV in zebrafish larvae was evaluated under normal conditions and under the effect of substances that modify intrinsic cardiac activity and cardiac activity modulated by the nervous system. We found that the factors affecting intrinsic activity have negative chronotropic and arrhythmogenic effects at this stage of development, whereas neural modulatory factors have a lesser impact. The results suggest that cardiac activity largely depends on the intrinsic properties of the heart tissue in the early stages of development and, to a lesser extent, in the maturing nervous system. We also report, for the first time, the influence of the neurotransmitter gamma amino butyric acid on HRV. The results demonstrate the larval zebrafish model as a useful tool in the study of intrinsic cardiac activity and its role in heart diseases.
Introduction
C
In mammals, it has been anatomically observed that asymmetry in cardiac sympathetic innervation translates into functional differences. The SNS innervates the sinus node and the right atrium through the right stellate ganglion, whereas the left stellate ganglion mainly innervates the left ventricle. The effect of this asymmetry is that right stellate ganglion activation increases heart rate (HR, positive chronotropism) by acting on the sinus node and generating an anti-arrhythmic effect, whereas left stellate ganglion activation stimulates the myocardium (positive inotropism) and tends to be pro-arrhythmic and hypertensive. 12 In contrast, the PNS regulates cardiac activity via the vagus nerve, which originates in neurons in the brain stem in the nucleus ambiguus and dorsal motor nucleus of the vagus; their axons have connections with intrinsic cardiac ganglia that are located in the right atrium wall. Postganglionic neurons innervate the sinus node, and the activation of these neurons increases vagal activity, which reduces HR.13–15 In addition, a complex neuronal net with diverse neurons has been described and shown to interact with both the SNS and the PNS.8,16
Because of this organization, HR varies over time according to the balance between sympathetic and parasympathetic activity. Sympathetic and parasympathetic influences on heart activity are revealed by analyses of the time intervals between heartbeats, which appear as R waves on an electrocardiogram (ECG) or peaks in pulse wave records. The intervals between consecutive R waves on an ECG are usually analyzed. This method is termed heart rate variability (HRV) analysis and comprises a linear and nonlinear analysis. In the linear component, analysis of the time domain determines the HR and RR intervals (interval between two consecutive peaks), whereas frequency domain analysis extracts the frequency and power spectrum of the different signal components. Nonlinear analysis is a study that evaluates two consecutive RR intervals and projects them in a plane called the Poincaré graph to assess abnormal RR intervals that are undetectable in linear analysis.17–19
The HRV analysis in the time domain is useful for long-term assessments, whereas the analysis in the frequency domain allows the evaluation of short periods of cardiac activity. 20 The HRV analysis in the frequency domain shows that parasympathetic activity predominates during sleep and rest and is a predominant component of the high-frequency band (HF band), which is interpreted as high HRV. The sympathetic influence determines activity in the low-frequency band (LF band), which determines a high HR, and a low HRV in the time domain. High HRV ensures cardiac adaptability, and low HRV represents limitations in cardiac adaptation and a low capacity to maintain body homeostasis, which is a characteristic of various heart diseases.21,22
Some authors have suggested that diseases such as acute myocardial infarction, heart failure, and hypertrophic cardiomyopathy may occur due to an imbalance in cardiac sympathetic innervation, which would favor episodes of arrhythmia and sudden cardiac death.18,23 It has also been proposed that pharmacological interventions could decrease the likelihood of changes in variability; this hypothesis is supported by results observed in experimentally denervated animals and in patients receiving beta-blocker drugs in whom the risk of sudden death was reduced because the ventricular fibrillation threshold was increased several times. 24
Despite the potential clinical utility of a study on HRV, little is known about its development and the elements that determine it or the instruments used for its evaluation.17,22 The use of low-cost experimental animal models to study the development of HRV, assess methods of analysis, and establish its relationship with cardiovascular diseases could be a fundamental tool to clarify these aspects.19,25 One of these animal models is the zebrafish larva model.
Zebrafish is a small freshwater fish that is easily acquired and has a high reproductive capacity. An important advantage of this organism is that fertilization and embryogenesis occur externally and that development from fertilization to young adult is fast and completed in ∼3 months. In addition, the fish is transparent in the early stages of development, enabling the observation of the development of internal organs with conventional microscopy techniques. 26 The size of adult fish (usually less than 5 cm in length) facilitates the maintenance of many specimens in a small space and reduces the costs associated with equipment installation and maintenance compared with the costs for rodent laboratory animal housing. The need for animal handling is minimal, which also reduces the requirement for technical staff. Zebrafish is used in multiple areas of research, including biochemistry, genetics, pharmacology, toxicology, oncology, neurosciences, and cardiology, among others.27–34
Given the characteristics of zebrafish production, their early visceral development and transparency as larvae, zebrafish is an ideal model for studying cardiovascular function.35–38 From a cardiovascular standpoint, zebrafish has substantial advantages, because its heart develops rapidly and is functional at 2 days postfertilization (dpf) and because many of the genes that determine its cardiac development have been identified.39,40 Also, cardiovascular morphological and physiological changes in embryos and larvae can be directly observed by using conventional microscopes.41,42 In addition, in the first week of development, zebrafish larvae can survive without circulation, because embryo and larval oxygen consumption are independent of cardiac function and their tissues can meet their needs via simple diffusion of oxygen from the environment, which is different from adult fish.43,44
Though the zebrafish heart has a simpler structure than the human heart, it possessed analogs of the mayor components of the mammalian hearts. It has been found to be a cardiac conduction system with pacemaking and conduction tissues comparable with those of higher vertebrates and humans. In this system, the electrical signal initiates in the zebrafish sinoatrial pole, equivalent to SA node in mammals, and travels through the atrium to the atrioventricular region, a specialized tissue at which electrical signaling reduces intensely. In the end, the signal travels through a ventricular conduction system to the apex and to the rest of the ventricle.45–48
The ECG shows a P wave that represents atrial depolarization, a QRS complex (with Q, R, S waves) that represents ventricular depolarization, and a T wave that represents ventricular repolarization.49,50 The shape of its action potentials are very similar to those of mammalian hearts and depend on ions channels, as Nav1.5 sodium channel, Cav1.2 and Cav3.2 calcium channels, and ether-a-go-go related gene (ERG) potassium channel. 50 The HR of zebrafish is closer than humans in comparison with other animal models, including mice and rats. Furthermore, as another fish, cardiac output is modulated by a dual autonomic innervation engaged by internal (blood pressure) and external (oxygen, CO2) changes.51,52
Finally, using immunohistochemical and neurotracing techniques, extrinsic and intrinsic cardiac innervation has been described and neurotransmitters (acetylcholine, adrenaline, noradrenaline) and cotransmitters (vasoactive intestinal polypeptide; nitride oxide) have been detected in the heart of zebrafish, in concordance with the results of studies of the heart of goldfish.53,54 Although GABA (an inhibitory neurotransmitter) was not evaluated in the intracardiac nervous system, GABAergic neurons have been detected in the midbrain, the hindbrain, and the spinal cord in zebrafish larvae. 55 All these aspects make zebrafish at early stages an ideal model with which to study intrinsic cardiac activity regulated by neural activity and its role in HRV. To date, there is not enough information regarding regulatory factors and HRV in zebrafish larvae.
Our aim was to evaluate HRV analyses and to explore the role of intrinsic cardiac activity and extrinsic neural activity in HRV in larval zebrafish, specifically focusing on the potential role of the neurotransmitter GABA. In the present study, HRV in zebrafish larvae was assessed under normal conditions and under the acute effect of excitatory substances such as caffeine, dopamine, and bicuculline or depressant stimulus such as cold water, alcohol, lidocaine, acetylcholine, methacholine, and muscimol.
Materials and Methods
Subjects
Wild adult zebrafish, which were kept in individual static tanks under appropriate temperature (25°C), pH (7.3), and light-dark cycle (14–10) conditions, were used to obtain larvae. The zebrafish were separated by gender, and a pair of adult fish were placed in an aquarium the day before reproduction. Fertilized eggs were collected and placed in Petri dishes under appropriate light and temperature conditions up to the time of the experiments. The zebrafish care protocol was approved by the Research and Ethics Committee of the Faculty of Medicine and the AICUC of the Pontificia Universidad Javeriana. Eighty-four zebrafish larvae, 5 ± 2 dpf, 3.8 ± 2 mm in length, were used in this study. Normal larvae with normal appearance, without morphological alterations, and with normal rhythmic heart activity were selected. The number of subjects is consistent with those reported in similar publications and follows the three Rs principle. 56
Zebrafish larvae were distributed randomly into control and experimental groups. Initially, the larvae were transferred to Petri dishes with aquarium water at 25°C and anesthetized by using a 0.6 mM MS-222 solution (tricaine methanesulfonate 3-ethyl aminobenzoate; Sigma-Aldrich, St Louis, MO), to reduce motion artefacts due to larvae movements. Larvae were positioned on the right side for evaluation and video recording. Direct evaluation of the cardiac activity was performed by using an upright microscope (Accu-Scope® EXC-350; Accu-Scope Inc., Commack, NY) and video recording for 30 s for subsequent analysis. Videos were collected by using an Optikam 2 Mp camera (Optika SRL, Ponteranica, BG, Italy) and ToupView Software (ToupTek Photonics, Zhejiang, P.R. China). This system provides imaging capacity of 30 fps (frames per second) at 640 × 480 pixels and a data rate of 13,500 kbps (kilobytes per second). Later, substances were added to the medium for the baseline registry. After 3–5 min of treatment, the cardiac contractile activity was again recorded for 30 s.
Drugs
The acute effect of the following drugs on the cardiac activity was evaluated: 4% ethanol, 1 mM caffeine, 0.43 mM lidocaine, 69 μM acetylcholine, 1 mM and 10 mM methacholine, 10 mM dopamine, 1 mM bicuculline, and 100 μM muscimol (Sigma-Aldrich). The effect of low temperature (6°C ± 2°C) was also evaluated. Drugs were diluted by using MS-222 solution to make up the final concentration. The drug doses were determined from data obtained in our previous published work and data reported in recent literature. In addition, previous drug toxicity assay was performed for all drug doses. For this, a toxicity drug test was running to detect mortality or changes in behavior, exploring tremor or seizures, which could cause noise during video recording.
Due to this evaluation, it was not possible to assess the HRV at the 2.5 mM caffeine concentrations, because the larvae exhibited involuntary movements, which generated signal noise, and it was not possible to evaluate the signal. Similarly, the HR was evaluated with or without MS-222 in the medium. No differences were found between HR of larvae with and without MS-222. The HR of each larva was recorded and compared with the cardiac activity after applying the drug.
Data analysis
HRV was analyzed from the recorded videos. Image analysis software Ethovision XT® (Noldus Information Technology, Inc., Leesburg, VA) was used to automate the procedure for counting the HR. This software allows the demarcation of a heart area and measures the number of pixels in the area with changes in brightness over time during ventricular filling. The variation in the number of pixels in the area was represented in a graph as a percentage of the activity and reflected the cyclical waves with peaks and valleys, which made it easy to count the heart beats (Fig. 1A). Previous studies have shown a constant short delay (30 ms) between ventricular depolarization and contraction. 57 This represents a good correspondence between ECG signal and ventricular filling detected with optic methods and could be a good estimator of HR. 58

HRV analysis method in zebrafish larvae.
Data from each record were exported and archived in text format for further HRV analysis. Kubios HRV software (Fig. 1B) was used for the HRV analysis. The software was developed by the University of Eastern Finland and is available at http://kubios.uef.fi, free of charge. In this program, the video cardiogram data were imported and graphed; the intervals between the peaks (RR) were measured to generate a tachocardiogram (Fig. 1C). Then, the HRV analysis was performed at each signal and included a linear and nonlinear analysis (Table 1). 59
bpm, beats per minute; HF, high frequency; HR, heart rate; LF, low-frequency; RMSSD, root mean square differences of succesive RR intervals; RR, interval between two consecutive peaks; VLF, very low frequency.
The linear analysis included the following two components: (1) time domain analysis and (2) frequency domain analysis. In the first case, the HR, its standard deviation (SDHR), the RR interval average, and its standard deviation (RR, SDNN) were analyzed. The following three frequency bands were established for the analysis in the frequency domain: (1) very low frequency (VLF, 0.0–0.04 Hz), (2) LF (0.04–0.15 Hz), and (3) HF (0.15–0.4 Hz); the frequency and power spectrum components of the signal, as well as the LF/HF ratio, were determined in each band.
Nonlinear analysis provided two consecutive RR intervals (RRn and RRn +1), which were projected on the Poincaré graph and used to adjust to an elliptical function. The two parameters are the measurement SD1, which describes the short-term variability in relation to breathing or respiratory movements, and SD2, which describes the long-term variability (Fig. 1C; Table 1). These parameters were evaluated both before and after the application of drugs or a change in the temperature, which ranged from 23°C to 25°C to 6°C ± 2°C. Researchers who are interested can find more detailed information about HRV parameters in previous HRV standards report and reviews.58–60
A paired Student t-test was used to assess the effects of each drug. A significance level of p < 0.05 was used.
Results
In this article, the acute effect of interventions that can modify intrinsic cardiac activity or cardiac activity modulated by the autonomic nervous system was assessed by using HRV analysis. The intrinsic cardiac activity can be altered by factors that modify heart tissue properties, which depend on both the membrane and the cellular microenvironment and include hypothermia, caffeine, lidocaine, and alcohol.
Substances that can modulate neural activity include acetylcholine, methacholine, dopamine, bicuculline (GABAA antagonist), and muscimol (GABAA agonist). These agents interfere with adrenergic and cholinergic transmission, as reported by other authors, and modify the action of the neurotransmitter GABA, which has not been previously reported.
Intrinsic cardiac activity and HRV in zebrafish larvae
Low temperature: HRV analysis in the time domain showed that exposure of the larvae at 6°C ± 2°C caused a significant decrease in the HR and an increase in the duration of the RR interval average (n = 9; 102 ± 10 vs. 63 ± 7 bpm; 599.38 ± 54.43 ms vs. 1013.08 ± 226.01 ms; p < 0.05) compared with cardiac activity at 23°C–25°C. No significant differences were observed in the frequency domain, though in several cases, the LF/HF ratio increased by decreasing the medium temperature. This finding indicates an increase in the LF component and, therefore, sympathetic activity. In the nonlinear analysis, significant differences were observed in SD2 (Fig. 2).

HRV and intrinsic activity. Increased RR interval duration and significantly decreased HR were observed on exposure of zebrafish larvae to low-temperature medium and 4% alcohol. The RMSSD parameter increased with exposure to lidocaine, suggesting an increase in vagal tone. Bar graphs show the values (means ± SD) of parameters. Asterisks represent significant differences. bpm, beats per minute; Caff, caffeine; Ct, control; Lid, lidocaine; OH, alcohol; RMSSD, root mean square differences of succesive RR intervals; RR, interval between two consecutive peaks; Temp, temperature.
Lidocaine: Exposure of larvae to 0.43 mM lidocaine causes an increase in the RMSSD compared with basal root mean square differences of succesive RR intervals (RMSSD) records (n = 12; 67.33 ± 59.65 vs. 110.68 ± 108.69; p < 0.05). Some authors suggest that this variable is associated with an increase in vagal activity. 61 Significant differences in the NN50, pNN50, and TINN were also observed. Definitions and details of these parameters have been described in numerous papers (Table 1).58–60 No significant differences were observed in the frequency domain. Significant differences in SD1 were observed in the nonlinear analysis (Figs. 2, 4).
Alcohol: Exposure of the larvae to 4% alcohol causes a significant increase in the average RR and a decrease in the HR compared with baseline records (n = 12; 625.47 ± 59.43 ms vs. 726.78 ± 77.94 ms; 99 ± 11 bpm vs. 85 ± 8 bpm; p < 0.05). Arrhythmic heart beat occurred in some cases. No significant differences in the analysis of the frequency domain or nonlinear analysis were observed. However, in some larvae, periods of asystole, which changes the appearance of the Poincare graph, were evident (Figs. 2–4).
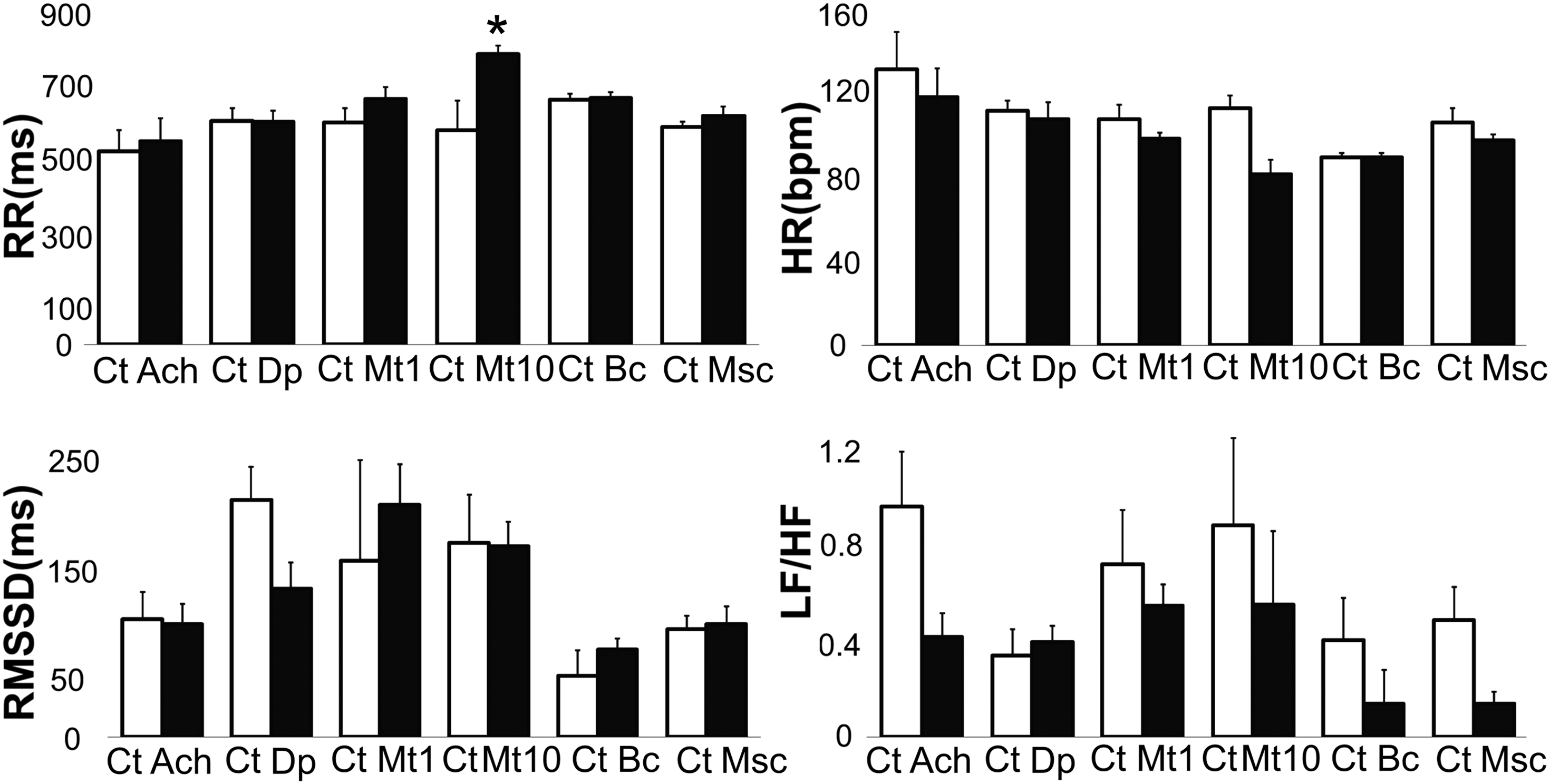
HRV and neural regulation. Increased RR interval duration and significantly decreased HR were observed on exposure of zebrafish larvae to 10 mM methacholine, an acetylcholine agonist. The RMSSD parameter decreased with exposure to dopamine, suggesting a decrease in vagal tone or an increase in sympathetic tone. The LF/HF ratio decreased with exposure to muscimol, a gamma amino butyric acid agonist, suggesting an increase in vagal tone. Bar graphs show the values (means ± SD) of parameters. Asterisks represent significant differences. Ach, acetylcholine; Bc, bicuculline; Ct, control; Dp, dopamine; Msc, muscimol; Mt 1, 1 mM methacholine; Mt 10, 10 mM methacholine.

Nonlinear analysis and intrinsic cardiac properties. Examples of Poincaré plots are shown. The graphs show the variability of the RR intervals. An increase in the duration of the RR intervals was observed. RR intervals greater than 2000 ms were observed on exposure to low temperature and alcohol, reflecting the occurrence of arrhythmia episodes. RRn and RRn +1 are two consecutive intervals.
Caffeine: Exposure to 1 mM caffeine generated a significant difference in the SDHR in comparison to the control (n = 9; 34.98 ± 22.38 vs. 23.91 ± 21.33; p < 0.05). A decrease in HR was observed in some cases, though it was not statistically significant. Arrhythmic heart beat episodes were also observed in some cases (Figs. 2–4).
Extrinsic cardiac activity and HRV in larval zebrafish
Acetylcholine and methacholine: Exposure to acetylcholine resulted in a decrease in HR and LF/HF ratio in some cases, indicating an increase in the HF component, though these changes were not statistically significant. Nonlinear analysis showed significant differences in SD2 (n = 5; 121.08 ± 29.35 vs. 81.20 ± 46.57; p < 0.05). Exposure to 1 mM methacholine (n = 5), an acetylcholine analog, more stable and less sensitive to acetylcholinesterase (AChE) activity than acetylcholine, decreased the HR, but the decrease was not statistically significant. Exposure to 10 mM methacholine showed significant differences in the RR average interval, HR, and SDHR (n = 7; 582.03 ± 59.67 vs. 789.19 ± 211.49; 114 ± 16 vs. 83 ± 17; p < 0.05). This result indicates that the effect of acetylcholine is inconsistent, and probably this is due to its fast inactivation by AChE. 53 The effect of methacholine was dose dependent and consistent with cholinergic activity. No significant differences in the analysis of the frequency domain or in the nonlinear analysis were observed (Figs. 3, 5).

Nonlinear analysis and neural regulation. Examples of Poincaré plots are shown. The graphs show the variability of the RR intervals during different interventions. The greater dispersion with methacholine and muscimol reflects greater variability, whereas less scatter with dopamine reflects lower variability. RRn and RRn +1 are two consecutive intervals.
Dopamine: The exposure of larvae to dopamine showed no significant changes in the HR or RR interval average. Significant changes in the RMSSD and SD1 were observed (n = 9; 214.46 ± 72.51 vs. 167.57 ± 90.75; 154.54 ± 52.42 vs. 119.79 ± 64.94; p < 0.05). This result may reflect a decrease in vagal activity.
Bicuculline and muscimol: Exposure of larvae to bicuculline, a GABAA receptor antagonist, 62 showed significant differences in the TINN (n = 7; 130.71 ± 56.01 vs. 173.57 ± 97.63; p < 0.05). Exposure to muscimol, a GABAA receptor agonist, 63 showed significant differences in the LF/HF ratio, which decreased (n = 9; 0.49 ± 0.43 vs. 0.14 ± 0.15; p < 0.05). This finding may indicate an increase in the HF component and, therefore, increased parasympathetic activity or a decrease in LF and thus sympathetic activity (Figs. 3–5). These data, although they are not conclusive, may indicate a possible modulatory role for GABA in the sympathetic-parasympathetic balance, which could be more evident in later stages of development.
Analysis and Discussion
The aim of the present study was to evaluate HRV under normal conditions and to compare it with activity influenced by different modulating elements, some of which are related to neural control, such as dopamine, cholinergic agents, and GABA agents, and other compounds related to intrinsic cardiac activity, such as lidocaine, alcohol, caffeine, and low temperature.
The drugs evaluated are highly soluble, which ensures that they penetrate different compartments, and affect myocardial tissue and the central and peripheral nervous systems via a simple media dilution without the use of special injection techniques. In this sense, the aim of the project was not to determine the effective dosage level of the drugs but, rather, to explore their ability to induce or prevent changes in cardiovascular activity. However, because there is no definite or stable integration between cardiac and nervous tissue in the early stages of development, it is possible to identify and characterize intrinsic cardiac activity at this stage of development and identify early influences on the nervous system.
Intrinsic cardiac activity and HRV in larval zebrafish
A negative chronotropic and arrhythmogenic effect was observed on exposure of zebrafish larvae to low temperature, caffeine, alcohol, and lidocaine, indicating a possible alteration in cardiac tissue properties, which depend on sodium channels, potassium channels, and secondary messengers such as calcium in myocardial cells.
Lin et al. reported that there is a direct relationship between a decrease in temperature (4°C) and a decrease in HR and proposed that the temperature could affect ion channels, thereby prolonging the duration of a cardiac tissue action potential, with a greater effect on atrial tissue. 64
Rana et al. reported a decrease in HR with exposure to caffeine that was dose dependent and caused cardiac arrest in some cases. The effects were not reversed on addition of ruthenium, adenosine, or calcium to the medium. These trials suggest that the ryanodine receptor and calcium-induced calcium release are also involved in cellular plasticity processes. The authors suggested that the negative chronotropic effects could be due to the early presence of adenosine receptors, which modulate the activity of potassium channels, maintain cardiomyocyte membrane hyperpolarization, and, therefore, reduce HR. 65
Although most studies on the effects of alcohol on the heart have examined chronic exposure and its effects on development,66–68 it has been shown that acute alcohol administration decreases myocardial contractility and ejection fraction in animal models and humans, even at concentrations as low as 0.05%.69–71 The depressor mechanism of alcohol on cardiac activity is related to its ability to interact with cell membrane calcium channels, which modify the intracellular calcium signals that are key during myocardial contraction.72,73 Another possible effect of alcohol has to do with its role as a positive allosteric modulator of GABAA receptors, which increase GABA currents. 74 However, during the early stages of zebrafish development, a greater influence would be on intrinsic cardiac activity and, to a lesser extent, on neural regulatory activity during the maturation process. This mechanism may be more important in later developmental stages, where alcohol may have an additional effect similar to that of the GABAA agonist muscimol.
Studies have shown a depressor effect for lidocaine on the heart. Lidocaine reduces cardiac activity in isolated, perfused rat hearts because of the inhibition of both AV conduction and atrial activity in the rat heart. Lidocaine decreased the diastolic resting potential, the maximum rate of depolarization, and the action potential amplitude, which was dose dependent, in an isolated rabbit Purkinje fiber-ventricular muscle model.75,76
Neural control or extrinsic cardiac activity and HRV in larval zebrafish
Drugs that modulate neural activity had less of an impact on HRV, and arrhythmogenic effects were not observed.
Methacholine, a cholinergic agent, showed a dose-dependent effect that decreased HR, increased RR, and decreased the LF/HF ratio. The dose of dopamine that affected beta-adrenergic receptors showed no significant changes in the HR, though it decreased the RMSSD, which may indicate a decrease in parasympathetic activity. Schwerte et al. characterized the sympathovagal balance in zebrafish larvae and reported that the response to adrenergic stimuli appears at 4 dpf and that the cholinergic response appears at 5 dpf, though this result was not always consistent or expected in the first week. This effect would reflect immaturity in neuromyocardial structures and suggests that the sympathovagal balance appears after 12 dpf. 77
The data reported here are consistent with and complement those results; our study assessed drugs such as methacholine and dopamine and add new information regarding muscimol and bicuculline. We found that muscimol, a GABA agonist, showed a positive effect on parasympathetic activity, which was reflected by a decrease in the LF/HF ratio. It has been reported that GABA and its agonists reduce the HR in isolated rat hearts. However, the application of GABA to vagal neurons increases the HR by inhibiting the effect of GABA on these neurons. Unexpectedly, bicuculline reduced the LF/HF ratio, though these data were not statistically significant. This effect may indicate an excitatory effect on vagal neurons, which may increase parasympathetic activity.
Briefly, we can conclude that HRV analysis, especially time domain and nonlinear components, can be used to evaluate heart activity in zebrafish larvae. HRV analysis in the time domain showed chronotropic effects for the tested agents, which was reflected in the HR and interval RR averages. An arrhythmogenic effect was most evident in the nonlinear analysis and Poincare charts, especially in the case of alcohol, where 2000 ms RR intervals were observed, reflecting a moment of asystole that was not visible in the linear analysis (Fig. 4). In contrast, HRV analysis in the frequency domain was not conclusive; no significant data in the HF or LF components were observed in the frequency domain, though the VLF component was more noticeable in some cases, which may indicate that this component reflects the intrinsic cardiac activity-dependent metabolic demands of the heart tissue raised by some authors. 78 Further studies are necessary to standardize HRV analysis in the frequency domain in zebrafish.
Cardiovascular diseases are the top cause of morbidity and mortality in human beings worldwide, and sudden cardiac death is a common outcome in many of them. It has been observed there are changes in the innervation of the heart in cases of sudden cardiac death associated with arrhythmias, acute myocardial infarction, heart failure, and some neurological diseases, though it is not clear what mechanisms are involved in these disorders. 79
It has been proposed that an autonomic imbalance between the cholinergic and adrenergic systems causes sudden cardiac death. There is also evidence of the relationship between alterations in cardiac innervation, particularly cholinergic-adrenergic changes not only in sudden cardiac death but also in arrhythmias, acute MI, and heart failure, though it is unclear what mechanisms are involved in these disorders.12,24 However, it is necessary to consider the role of other agents that have been identified at the central nervous system level, including the peripheral level (growth factors, neuropeptides, and GABA), that may participate in these processes.
A large number of studies show that GABA is present in both the central and peripheral nervous systems and that it participates in cardiovascular regulation mechanisms by modulating circuits in the brain stem, spinal cord, and sympathetic chain.10,11,80,81 However, in contrast to the extensive research on the role of GABA in the CNS, little is known about the role of GABA in the peripheral nervous system.82,83 Some studies have shown that preganglionic fibers are immunoreactive to GAD67, a glutamate decarboxylase enzyme subtype responsible for GABA synthesis, in the superior sympathetic cervical ganglion.7,80 Similarly, connections between supraspinal preganglionic GABAergic neurons and sympathetic preganglionic neurons have been identified. 9
Other authors have reported that GABA modulates synaptic plasticity in the SCG and affects synaptogenesis,7,9,80 though the specific role of GABA at this level is unclear. It is also unclear whether there is a direct relationship between sudden cardiac death and GABAergic neural control. Recent studies have shown that GABA and some neurotrophic factors may have a role in HRV and that they can modify their expression to conform to demands under normal and pathological conditions. It has also been reported that benzodiazepines, GABA agonists with anxiolytic and anticonvulsant effects, reduce vagal activity, increase HR, and reduce HRV. 84
In our experiments, muscimol decreased HR, which agrees with insulated heart experiments and may indicate an effect on intrinsic cardiac activity. 85 Bicuculline, a GABA antagonist, displayed a negative chronotropic effect that can be explained by the excitatory actions of vagal neurons. It has been reported that GABA has an excitatory effect in early development due to high chlorine intracellular concentrations, which are secondary to the predominance of the chlorine input cotransporter NKCC1. This changes to an inhibitory GABA phenotype in adult life due to the increased expression of the cotransporter KCC2 chlorine output. 82 The observed effect of GABA agonists and antagonist agents on HRV reinforces the hypothesis, suggesting an immature regulation system in which neural connections are not well established. 85 This conception implies that the effect described here due to GABA may change and become more evident at later stages of development, though additional studies are required to confirm this hypothesis.
In summary, the responses of zebrafish larvae heart to agents that modify cardiac activity, including caffeine, lidocaine, alcohol, hypothermia, dopamine, acetylcholine, methacholine, bicuculline, and muscimol, were explored in this study. We found that heart activity does not depend significantly on the neural regulation during the early stages of development in zebrafish, which has already been reported in other studies investigating the formation and maturation periods. In contrast, it has also been shown that intrinsic cardiac activity determines HR activity in the development stage, as it can easily be altered by factors that modify the electrochemical properties of cardiac tissue. This level of development should take into account the metabolic demand of the heart tissue, cell membrane components, and their properties and tissue microenvironment homeostasis as determinants of cardiac activity.
These aspects can be altered with stimuli such as caffeine, lidocaine, alcohol, and hypothermia. With advanced cardiovascular development, systemic blood flow, cardiac output, and an increased metabolic demand from tissues and interactions with the nervous system establish the role of neural and endocrine-controlling agents in cardiac activity.
We show for the first time the possible role of the neurotransmitter GABA in the regulation of cardiac activity. Finally, we show that zebrafish larva is an ideal model for the study of the intrinsic cardiac activity, which can be one of the most important causes for fatal outcomes that are common to many cardiovascular diseases, especially those in which cardiac tissue properties are altered, such as in ischemic disease, hypoxic states, cardiomyopathies, and metabolic heart disease, among other conditions. These results and the analytical instruments employed in this study, with other molecular and genetic tools, can be used for future heart physiology and pathophysiology studies, in which zebrafish are becoming more widely used.
Footnotes
Acknowledgment
This work was supported by a research grant from the Pontificia Universidad Javeriana, Bogotá, Colombia. Project: 5605; budget code 120112Z0401200.
Disclosure Statement
No competing financial interests exist.