
Abstract

Dear Editor:
I
One possibility is that bradycardia represents a primary response to heart failure. Since cardiac output = heart rate × stroke volume, an adaptive increase in heart rate serves as an important physiological response to maintain cardiac output in settings of contractile dysfunction. A true bradycardia would therefore accelerate a decline in overall cardiac performance in low cardiac output states in which myocardial contractile function is already compromised. The possibility that zebrafish are incapable of developing a compensatory increase in heart rate seems unlikely, however, given that tachycardia has been documented in a heart failure model associated with a sarcomere protein mutation. 3 Moreover, in our hands, we have noted a dose-dependent tachycardia during epinephrine administration in wild-type TE fish aged 6 months (Fig. 1A, B), demonstrating that zebrafish are able to mount an appropriate chronotropic response to increased sympathetic stimulation.
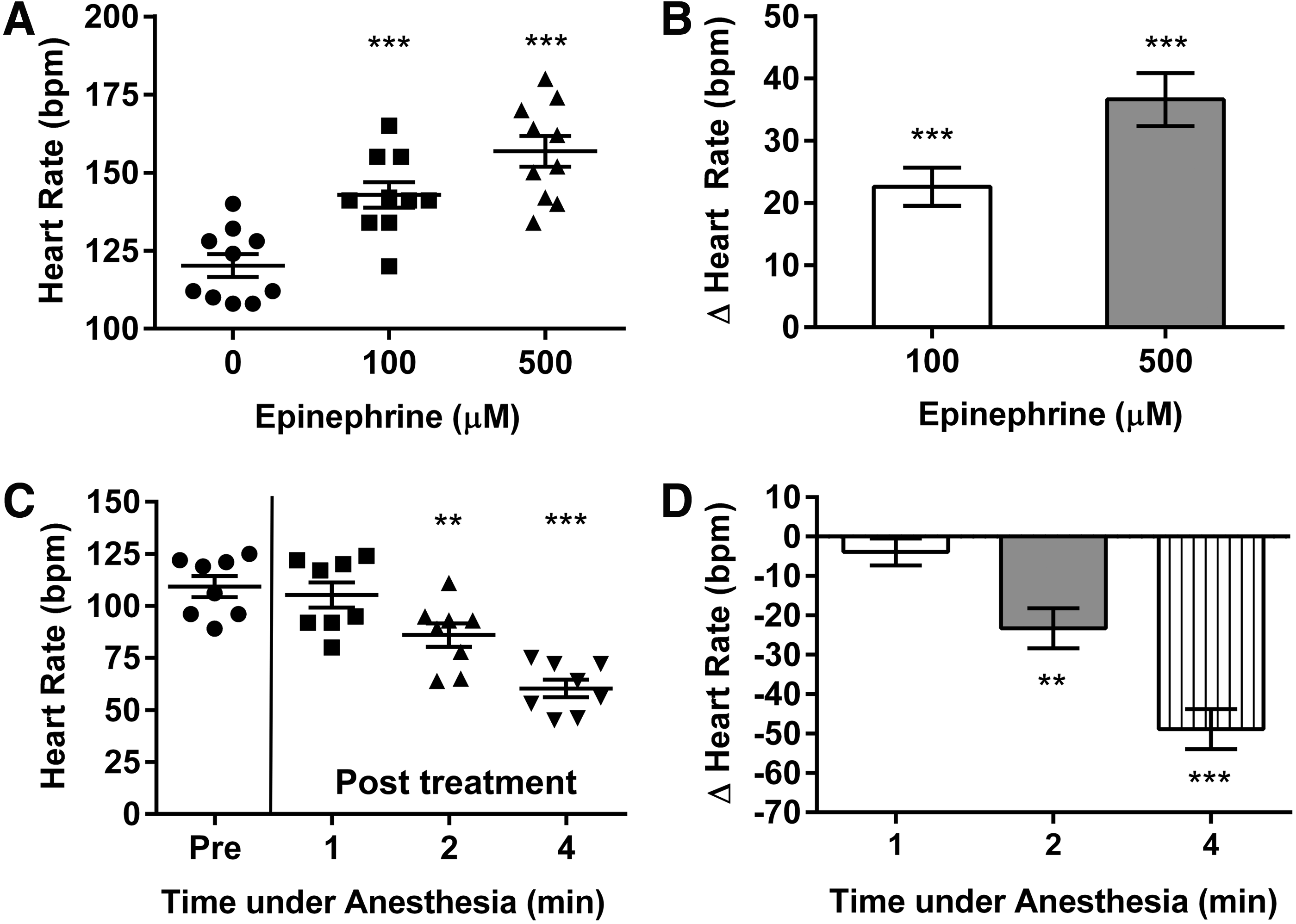
Heart rate responses to epinephrine and anesthesia. Heart rate was measured by observing cardiac contraction over a 15 s period on B-mode echocardiography performed at room temperature (25°C) in male wild-type TE fish anesthetized using tricaine methanesulfonate 200 mg/L (Sigma-Aldrich, St. Louis, MO).
Another possibility is that bradycardia might be indicative of advanced disease and a decompensated state. Phenylhydrazine (2.5 μg/mL) results in profound hemolytic anemia (relative hemoglobin <10% that of untreated animals).2,4 It is notable that Sun et al., who first reported the phenylhydrazine protocol used by Ernens et al., observed an initial tachycardia in unanesthetized tr265/tr265 transgenic fish with severe anemia at 5 and 10 days of age, but bradycardia from 3 weeks. 4 Interestingly, this time point also coincided with the need for anesthesia to facilitate measurements.
Anesthetic agents and temperature are known to influence heart rate during echocardiography,5,6 and Ernens et al. suggested that the dose of anesthetic agent used (tricaine methanesulfonate, 160 mg/L), and/or the performance of their studies at room temperature, may have contributed to their heart rate results. 2 These factors would apply equally to wild-type and anemic fish, however, and thus do not explain the observed differences between groups. 2 A parsimonious explanation is that anemic zebrafish have a substantially greater susceptibility than healthy zebrafish to cardiodepressant effects of anesthesia. We also found that a similar dose of tricaine caused bradycardia in anemic zebrafish, with progressive reductions of heart rate occurring with increasing duration of anesthetic agent exposure (Fig. 1C, D).
Optimizing anesthetic regimens to gain adequate sedation and image acquisition at the same time as minimizing side effects is challenging, and the use of anesthetic agent during echocardiography may confound interpretation of the underlying disease phenotype. It should be noted that in addition to negative chronotropic effects, anesthetic drugs can also directly depress myocardial contraction. Collectively, these considerations argue for a critical need to use minimum doses of anesthetic drugs when performing in vivo cardiac phenotype analysis, particularly in zebrafish heart failure models.
Footnotes
Acknowledgments
The authors are supported by the National Health and Medical Research Council of Australia, Heart Foundation of Australia, Royal Australasian College of Physicians, Estate of the Late RT Hall, St Vincent's Clinic Foundation, and the Ian Potter Foundation.
Disclosure Statement
No competing financial interests exist.