
Abstract
Abstract
Most species of the genus Harttia inhabits the headwaters of small tributaries, but some species are restricted to the main channel of some rivers. This feature, combined with limited dispersal ability, leads to the formation of small isolated populations with reduced gene flow. Currently, there are 23 taxonomically defined and recognized species, and 17 of these are found in Brazil, distributed in several hydrographic basins. Despite this diversity, few chromosomal data for the species belonging to this genus are found in the literature. Thus, this study analyzed, by classical and molecular cytogenetics methodologies, the chromosomal diversity of this genus, to discuss the processes that are involved in the evolution and karyotype differentiation of the species of the group. Seven species of Harttia were analyzed: H. kronei, H. longipinna, H. gracilis, H. punctata, H. loricariformis, H. torrenticola, and H. carvalhoi. The chromosomal diversity found in these species includes different diploid and fundamental numbers, distinct distribution of several repetitive sequences, the presence of supernumerary chromosomes in H. longipinna and multiple sex chromosome systems of the type XX/XY1Y2 in H. carvalhoi and X1X1X2X2/X1X2Y in H. punctata. Lastly, our data highlight the genus Harttia as an excellent model for evolutionary studies.
Introduction
T
There are 23 known species for the genus Harttia. 3 Of these, 17 are found in Brazil: H. absaberi Oyakawa, Fichberg, and Langeani, 2013; H. carvalhoi Miranda Ribeiro, 1939; H. depressa Rapp Py-Daniel and Oliveira, 2001; H. dissidens Rapp Py-Daniel and Oliveira, 2001; H. duriventris Rapp Py-Daniel and Oliveira, 2001; H. garavelloi Oyakawa, 1993; H. gracilis Oyakawa, 1993; H. kronei Miranda Ribeiro, 1908; H. leiopleura Oyakawa, 1993; H. longipinna Langeani, Oyakawa, and Montoya-Burgos, 2001; H. loricariformis Steindachner, 1877; H. novalimensis Oyakawa, 1993; H. punctata Rapp Py-Daniel and Oliveira, 2001; H. rhombocephala Miranda Ribeiro, 1939; H. torrenticola Oyakawa, 1993; H. trombetensis Rapp Py-Daniel and Oliveira, 2001; and H. uatumensis Rapp Py-Daniel and Oliveira, 2001.
In this genus, only seven species present cytogenetic data: H. kronei,4,5 H. loricariformis,6,7 H. carvalhoi,5,8 H. absaberi cit. Harttia sp. n., 9 H. longipinna, 10 H. torrenticola, 5 and H. punctata. 11 The karyotype macrostructure of the group is quite diverse. The diploid number varies from 52 chromosomes, found in females of H. carvalhoi from the Grande stream in Paraiba do Sul river basin,5,8 to 62 chromosomes in H. absaberi cit. Harttia sp. n from Passa Cinco river basin. 9
Cytogenetic analysis also demonstrated the occurrence of multiple sex chromosome systems in Harttia.5,8,11 Populations of H. carvalhoi have a diploid number of 52 chromosomes for females and 53 chromosomes for males, which characterizes a system of multiple sex chromosomes of the type XX/XY1Y2.5,8 In H. punctata, species with 2n = 57/58 chromosomes, was observed a multiple sex chromosomes system X1X1X2X2/X1X2Y. 11 In a population of H. longipinna, with 2n = 58 chromosomes, was observed an inter and intraindividual variation of up to two B chromosomes. 10
Given this context, this study aimed at understanding the origin of the chromosomal diversity in the genus Harttia, correlating chromosomal rearrangements and evolutionary mechanisms.
Materials and Methods
Specimens of seven species of the genus Harttia were collected from different Brazilian river basins (Table 1) and deposited in the zoological collection of the Museu de Zoologia da Universidade de São Paulo (MZUSP) (Table 1). All procedures were performed in compliance with the Ethics Committee on Animal Experimentation (Process No. 13/2014) of the Universidade Estadual de Ponta Grossa (UEPG).
GO, Goiás; GPS, global positioning system; MG, Minas Gerais; MZUSP, Museu de Zoologia da Universidade de São Paulo; PR, Paraná; SP, São Paulo; ♂, male; ♀, female.
The fishes were anesthetized with clove oil 12 and then sacrificed. Metaphasic chromosomes were obtained from cephalic kidneys.13–15 The chromosomes were organized in metacentric (m), submetacentric (sm), subtelocentric (st), and acrocentric (a). 16 The fundamental number (NF) was calculated considering metacentric chromosomes, submetacentric, and subtelocentric as bearing two arms, and acrocentric chromosomes as bearing only one. Heterochromatic regions were detected by the C-banding procedure 17 and the slides were stained with propidium iodide. 18 The nucleolar organizing regions (Ag-NORs) were obtained by silver nitrate staining. 19 Both methods were applied sequentially, after conventional chromosomal staining with Giemsa (sequential analysis).
The fluorescence in situ hybridization (FISH) 20 was performed using 18S rDNA, 21 5S rDNA, 22 and telomeric sequence [TTAGGG]n 23 probes. The 18S rDNA probe was labeled by nick translation with biotin-16-dUTP (Biotin Nick Translation mix; Roche Applied Science, Mannheim, Germany) and the 5S rDNA probe was labeled with digoxigenin-11-dUTP (Dig Nick Translation mix; Roche Applied Science). The telomeric sequence probe [TTAGGG]n was labeled by polymerase chain reaction, using digoxigenin-11-dUTP (Roche Applied Science). All hybridizations were performed with 77% stringency (200 ng from each probe, 50% deionized formamide, 10% dextran sulfate, 2 × saline sodium citrate (SSC); pH 7.0–7.2). After hybridization, the slides were washed for 20 min in a solution of 15% formamide/0.2 × SSC at 42°C, 0.1 × SSC at 60°C for 15 min and 4 × SSC/0.05% Tween for 10 min at room temperature, the latter consisting of two washes of 5 min. The signal detection was performed using avidin-FITC (Sigma-Aldrich, St. Louis, MO) for 18S rDNA probe; and anti-digoxigenin-rhodamine (Roche Applied Science) for 5S rDNA probes and telomeric sequence [TTAGGG]n. The chromosomes were analyzed using an Olympus BX51 epifluorescence microscope (Olympus, Tokyo, Japan). The software DP2-BSW (Olympus) was used for image capture. Approximately 30 metaphases were analyzed to determine the diploid number, karyotype formula, and the presence or absence of interstitial telomeric signal (ITS). The sites that showed 100% of repeatability were considered as a sign of true hybridization for all probes.
Results
Harttia kronei
The 18 analyzed specimens (11♂ and 7♀) of H. kronei showed 58 chromosomes (16m+16sm +16st +10a) and NF = 106, without a system of differentiated sex chromosomes (Fig. 1and Table 2). The C-banding showed a small amount of heterochromatin preferentially allocated in pericentromeric position of few chromosomes of the complement (Fig. 2). A prominent band in the interstitial position of the short arm of the second metacentric pair was observed, which is coincident with the NOR (Fig. 1) and the 18S rDNA site (Fig. 3). The FISH with 5S rDNA probe revealed a single site of this marker, which is allocated in pericentromeric position of the long arm of the first chromosome pair submetacentric (Fig. 3). No ITS was detected in the karyotype with the use of telomeric probe (Fig. 4).
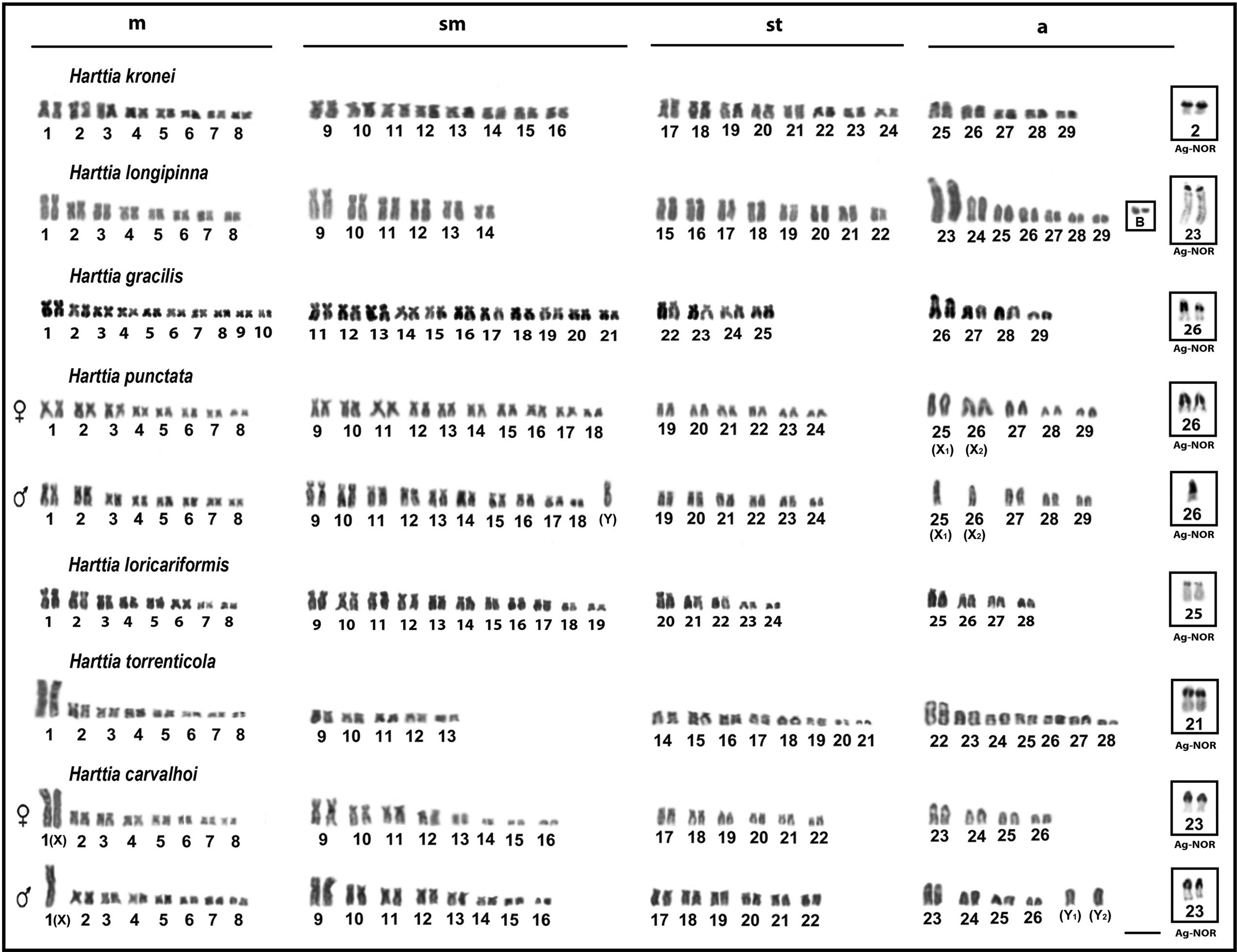
Karyotype of the Harttia species submitted to conventional staining with Giemsa. Ag-NORs are highlighted. Bar = 10 μm. Ag-NORs, Ag-nucleolar organizing regions.

Karyotype of the Harttia species submitted to C-banding. Bar = 10 μm.
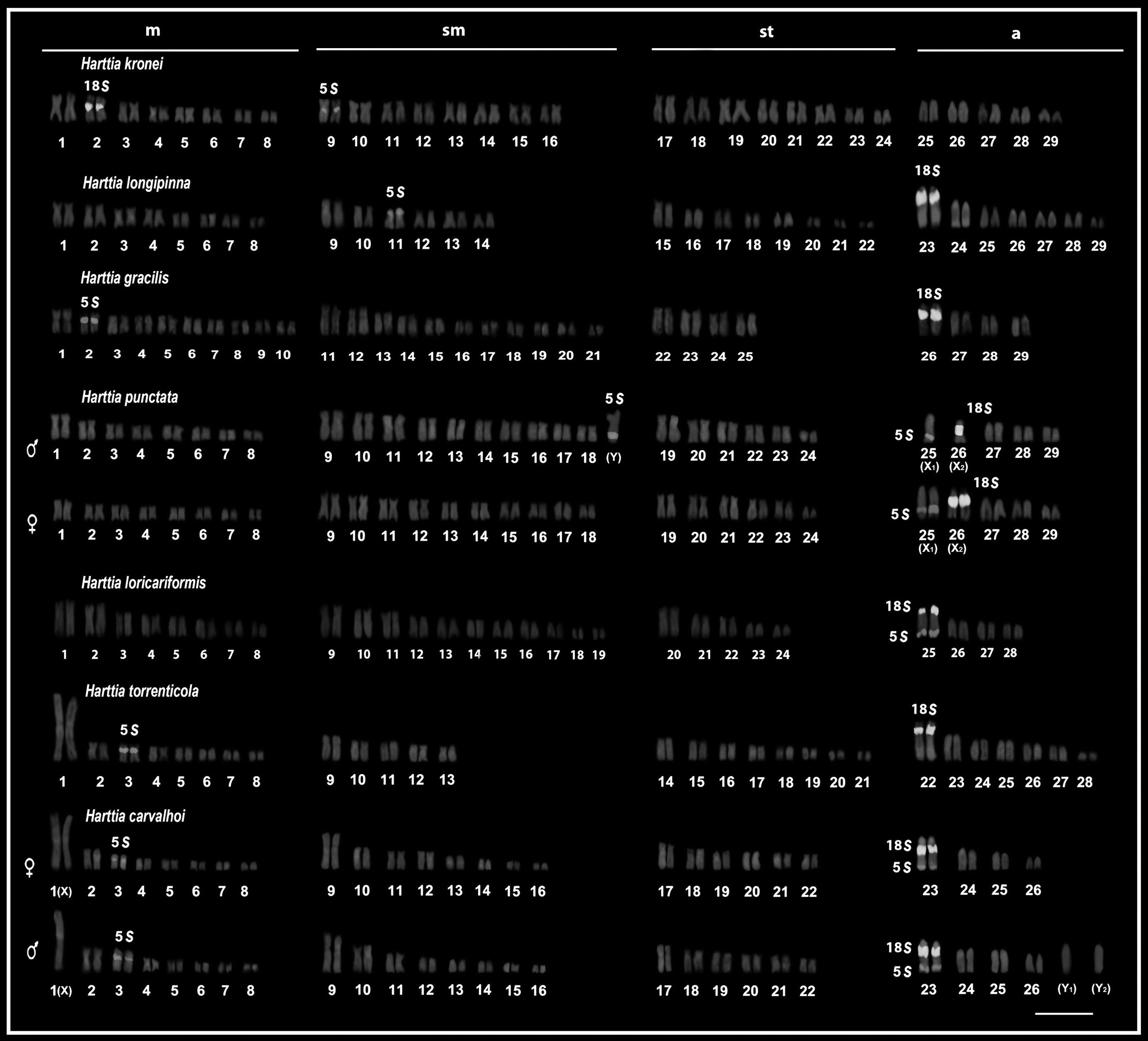
Karyotype of the Harttia species submitted to Double-FISH with probes of rDNA 5S and rDNA 18S. Bar = 10 μm. FISH, fluorescence in situ hybridization.
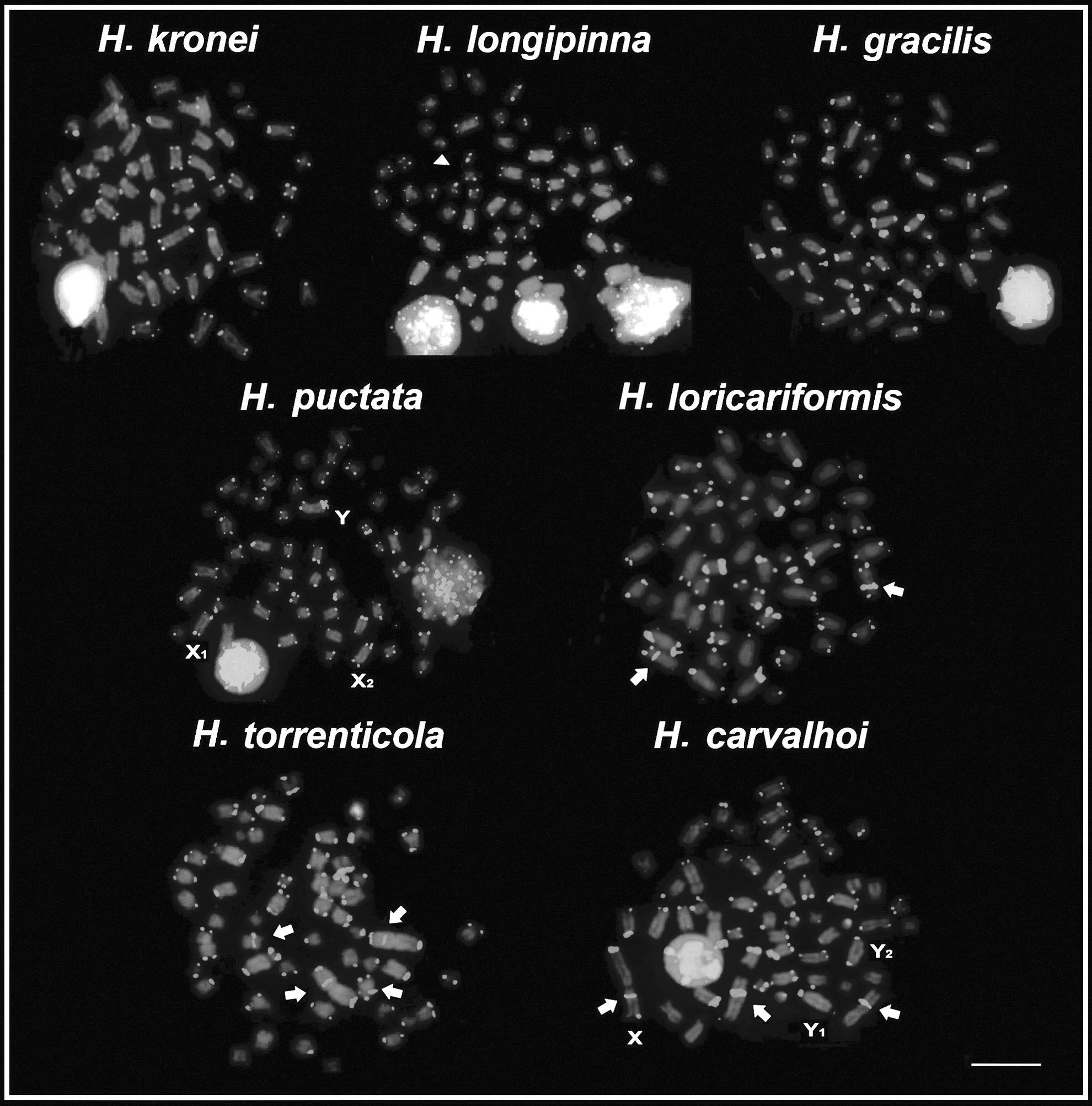
Metaphases of Harttia species submitted to FISH with probe of telomeric sequence (TTAGGG)n. The arrows indicate ITS and the arrowhead indicates B chromosome. Bar = 10 μm. ITS, interstitial telomeric signal.
a, acrocentric; m, metacentric; NF, fundamental number; Ref, reference; sm, submetacentric; st, subtelocentric; 2n, diploid number; ♂, male; ♀, female.
Harttia longipinna
The 50 analyzed specimens (30♂ and 20♀) of H. longipinna presented 2n = 58 chromosomes (16m+12sm +16st +14a) and NF = 102, in both males and females without a system of differentiated sex chromosomes (Fig. 1 and Table 2). In 23 specimens (16♂ and 7♀) a variation of up to two B chromosomes was detected. A small amount of heterochromatin was located in pericentromeric and terminal positions of some chromosomes of the complement A, especially the pair 23 that features two prominent C+ blocks, being a proximal and an interstitial on the long arm (Fig. 2). The B chromosomes showed totally heterochromatic (Fig. 2, in box). The impregnation by silver nitrate showed simple NORs allocated in the proximal region of the long arm of the pair 23, coinciding with the block of heterochromatin (Fig. 1). The location of the rDNA 18S probe by FISH (Fig. 3) confirmed the results on the number and location of the NORs. Hybridization with rDNA 5S probe revealed a single pair of this marker, present in pericentromeric position of the short arm of the submetacentric pair 11 (Fig. 3). FISH with telomeric probe showed no ITS in this karyotype (Fig. 4).
Harttia gracilis
The 23 analyzed specimens of H. gracilis (8♂ and 15♀) showed diploid number equal to 58 chromosomes (20m+22sm +8st +8a) and NF = 108, in both males and females without a system of differentiated sex chromosomes (Fig. 1 and Table 2). The heterochromatin showed pale staining allocated in the pericentromeric region of a few chromosome pairs, especially in the pairs 2 and 26 (Fig. 2). The silver nitrate staining showed a single active NOR in the proximal region of the long arm of the acrocentric pair 26, matching with the block C+ (Fig. 1), also subsequently confirmed by FISH with rDNA 18S probe (Fig. 3). Hybridization with rDNA 5S probe revealed the presence of a single site allocated in the pericentromeric position of the metacentric pair 2 (Fig. 3). No ITS was detected in the karyotype with the use of telomeric probes (Fig. 4).
Harttia punctata
The 32 analyzed specimens of H. punctata (18♂ and 14♀) showed diploid number equal to 58 chromosomes (16m+20sm +12st +10a) for females and 57 (16m+21sm +12st +8a) for males, both with NF = 106 (Fig. 1 and Table 2). Thus, it is noticeable that male heterogamety, which is represented by a large submetacentric chromosome (Y), present exclusively in the male karyotype, and two acrocentric chromosomes without their homologs (X1 and X2), which corresponds to the pairs 25 and 26 in the females (Fig. 1). The heterochromatin was preferentially allocated, in both sexes, in the pericentromeric regions of some chromosomes of the complement (Fig. 2). It is pertinent to emphasize three major heterochromatic blocks allocated in the two largest acrocentric pairs in both sexes: whereas in the chromosome pair 25 (X1) was found a proximal and a terminal block, and in the pair 26 (X2) was evidenced a large block in the interstitial portion, in the terminal portion of the long arm of the Y chromosome a prominent heterochromatic block was observed. The impregnation by silver nitrate showed simple NORs allocated in the interstitial portion of the acrocentric pair 26, such that only one chromosome homolog is present in the male karyotype (Fig. 1). Hybridization with rDNA 18S probe showed the interstitial region of acrocentric pair 26 (X2) labeled (Fig. 3). For the rDNA 5S probes, these sites were located at the terminal region of the acrocentric pair 25 (X1) and in the terminal region of the long arm of the Y chromosome (Fig. 3). Both in males and in females, the sites of 5S rDNA and 18S are coincident with heterochromatic blocks. No ITS was detected in the karyotype (Fig. 4).
Harttia loricariformis
The four specimens of H. loricariformis (2♂ and 2♀) showed diploid number equal to 56 chromosomes (16m+22sm +10st +8a) and NF = 104, in both males and females, without a system of differentiated sex chromosomes (Fig. 1 and Table 2). A small amount of heterochromatin was preferentially allocated in centromeric and pericentromeric regions of some chromosomes, especially in the largest acrocentric pair (pair 25) (Fig. 2). The silver nitrate impregnation showed a single pair of active NORs in the short arm of pair 25 (Fig. 1), coincident with the location of heterochromatin and rDNA 18S probe (Fig. 3). The cistron of rDNA 5S was allocated in the terminal portion of the long arm of the pair 25 in synteny with the rDNA 18S (Fig. 3). FISH with telomeric probe demonstrated the occurrence of ITS in a subtelocentric pair (Fig. 4).
Harttia torrenticola
The 13 specimens of H. torrenticola (5♂ and 8♀) showed diploid number equal to 56 chromosomes (16m+10sm +16st +14a) and NF = 98, in both males and females, without a system of differentiated sex chromosomes (Fig. 1 and Table 2). The C-banding showed a little amount of heterochromatin, which was mainly allocated in the centromeric and terminal regions of some chromosomes. Except for a large block that is located in the proximal region and a minor in terminal position in pair 22 and an evident block in the pericentromeric region of pair 1 (Fig. 2). The silver nitrate impregnation revealed simple NOR located in interstitial position of the long arm of the acrocentric pair 22, which coincides with the heterochromatic block (Fig. 1) and the site of rDNA 18S (Fig. 3). FISH with rDNA 5S probe showed a single marking on the pericentromeric region of a submetacentric chromosome (Fig. 3). FISH with telomeric probe demonstrated the occurrence of ITS, which was located on the short arm of a small submetacentric pair and in the short arm of the first metacentric pair of the karyotype (Fig. 4).
Harttia carvalhoi
The 24 specimens of H. carvalhoi (13♂ and 11♀) showed diploid number equal to 52 chromosomes (16m+16sm +12st +8a) for females and 53 chromosomes (15m+16sm +12st +10a) for males, and NF = 96 for both (Fig. 1 and Table 2). This difference in diploid number between males and females was observed by the presence of two additional acrocentric chromosomes in males, besides the absence of one chromosome of the first metacentric pair, consistent with the sex chromosome system XX/XY1Y2. The heterochromatin was mainly located in the pericentromeric and terminal regions of the chromosomes (Fig. 2). The chromosome pairs 1 and 9 show conspicuous pericentromeric bands. In the pair 23 was detected three heterochromatic blocks, one in the centromeric region, a small in the interstitial region on the long arm adjacent to the secondary constriction, and one in the terminal region of the short arm (Fig. 2). Staining with silver nitrate and FISH with rDNA 18S probe showed simple NOR, which was coincident with the secondary constriction and intercalated with the heterochromatic block (Figs. 2 and 3). FISH with rDNA 5S probe revealed two chromosome pairs that carry these cistrons. The largest site was located in the interstitial region of the long arm of the pair 23, just below the 18S sites (syntenic marks) and the minor site was located on the short arm of the metacentric pair 3 (Fig. 3). FISH with telomeric probe demonstrated the occurrence of ITS in the first submetacentric pair, and a second site located on the short arm of the largest metacentric pair (chromosome X) of the karyotype (Fig. 4).
Discussion
Harttia species analyzed in this work show considerable differences in the karyotype formula, NF and especially in the diploid number (Table 2), which varies from 52 chromosomes, found in females of H. carvalhoi,5,7 to 62 chromosomes, present in H. absaberi cit. Harttia sp. 9 This group has cases of B chromosomes, as in H. longipinna, 10 and cases of sex chromosomes systems: XX/XY1Y2 present in H. carvalhoi5,8 and X1X1X2X2/X1X2Y in H. punctata. 11 Centric fusions and fissions are the main events for karyotype differentiation in Loricariidae, 24 and chromosomal translocations and inversions are also important in the diversification of this group. 25 Thus, it is evident that the interspecific variation of the Harttia species here analyzed is the result of extensive chromosomal rearrangements like it was postuladed for Loricariidae in general.
Phylogenetic analysis with mitochondrial and nuclear data in many Harttia species show that the species analyzed in the present work are organized into two distinct groups: (1) composed of H. punctata and (2) composed of H. torrenticola, H. carvalhoi, H. gracilis, H. loricariformis, H. kronei, and H. longippina.26,27 The relations of the species are showed in the Figure 5. In Loricariidae, 2n = 54 chromosomes is considered a basal character.24,28 Chromosome analysis showed that H. punctata, belonging to the group (1) and H. kronei, the basal species from the group (2), present 58 chromosomes, and that the most common diploid number in this genus is 58. Thus, it is likely that the basal diploid number for the genus Harttia is 58 chromosomes. In this case, a hypothetical ancestral Loricariinae with 54 chromosomes would have resulted in the formation of a basal karyotype for Harttia with 58 chromosomes by fission events.

Using the phylogenetic relations26,27 as scenario and the ancestral of Harttia with 58 chromosomes and without sex chromosomes, the chromosome evolution of the group occurred by two distinct events of 2n reduction, one in the lineage of H. loricariformis, and the other in the lineage of the ancestral of the group H. torrenticola e H. carvalhoi. In this context, a third event of 2n reduction occurred in the lineage of H. carvalhoi (Fig. 5). The first chromosome pair of H. torrenticola and H. carvalhoi is larger than the corresponding pair of the remaining species. The presence of this large chromosome in these two species, associated to the ITS site in this pair, and the lack of this chromosome in H. loricariiformis suggests that the events of 2n reduction to 56 chromosomes is due to distinct events, reinforcing the hypothesis proposed. In this context, H. punctata differed by the appearance of the multiple sex chromosomes system of the type X1X1X2X2/X1X2Y, which originated by a pericentric inversion, with subsequent partial translocation. 11 H. kronei, on the other hand, differed by pericentric inversions and translocations, which altered the morphology of the chromosome pairs. The differentiation of H. loricariformis was caused by an event of centric fusion, confirmed by the ITS site in a chromosome pair, leading to the diploid number reduction to 56 chromosomes. To H. gracilis and H. longippina, the differentiation occurred by inversions, translocations, and deletions. The appearance of the cluster of H. torrenticola e H. carvalhoi occurred by an event of centric fusion, leading to the generation of a big chromosome pair and to the reduction of the diploid number to 56 chromosomes. H. carvalhoi differed by subsequent events of fusion, which reduced the chromosome number to 52, and by the appearance of the sex chromosome system XX/XY1Y2, which was due to a centric fission of the pair 1. 5
The separation of the groups (1) and (2) has strong statistical support, but the remaining relations among the species are uncertain.26,27 Nevertheless, the similarity of the molecular results of both works and the adequacy of cytogenetic data to this scenario are strong evidences of the veracity of these relations.
A single chromosome pair with NOR sites is considered a basal characteristic for Loricariidae.24,29 Staining with silver nitrate and FISH with rDNA 18S probe showed simple NORs in all species of Harttia. This feature could be a possibly primitive condition within the genus. Despite the maintenance of the number of 18S sites, there is variation in the position and in the type of chromosome that carry these cistrons between the species. H. longipinna, H. gracilis, H. punctata, H. torrenticola, and H. carvalhoi have the rDNA 18S in the proximal region of the first acrocentric pair, characteristic that can be considered conserved for this group of species. Exceptions of this position can be observed in three species: in H. kronei4,5 and H. absaberi cit. Harttia sp. 9 NORs were observed on the short arm of metacentric chromosomes; and in H. loricariformis these were observed in the short arm of the first pair acrocentric. These variations are possibly due to pericentric inversions. The physical mapping of 5S rDNA sequence has revealed that this cistron is variable in position and conserved in number, occurring in different chromosomes of the analyzed species. H. carvalhoi is the only species that have multiple sites of 5S rDNA. Thus, it is possible to infer that one pair of sites of this ribosomal sequence is a basal feature in Harttia.
The synteny of the rDNA sites can be considered a probable primitive condition in Loricariidae. 28 Thus, it is possible that this feature in Harttia effectively represents the basal character or simply be a byproduct of rearrangements that led to diversification of this chromosome. If we consider the synteny as a basal pattern for the Harttia genus, H. loricariformis and H. carvalhoi maintain the basal condition for these genes.
The presence of small amounts of heterochromatin is probably an intrinsic characteristic of the species of Harttia. Heterochromatic regions are recognized as “hot spots” for chromosome rearrangements. 29 In the Harttia species analyzed, heterochromatic blocks were preferentially located in the pericentromeric and terminal regions of some chromosomes. Possibly, the presence of repetitive DNAs in the heterochromatic regions in the chromosomes of Harttia has acted to diversify the genomes, contributing to the occurrence of chromosomal rearrangements that promote the differentiation of isolated populations in the headwaters or in specific stretches of rivers, similar to what occurs in other species of Neotropical fishes.
In general, the chromosomal data for the Harttia genus indicate the occurrence of great diversity, represented by differences in diploid number, karyotype formula, number and position of the 18S and 5S ribosomal genes, and distinct sex chromosome systems. All this chromosomal variations, which is accompanied by robust taxonomic characters, may be result of the behavior of these organisms. The Harttia species have limited capacity of dispersion and inhabit headwaters or specific portions of the rivers, allowing that process of chromosomal and morphological differentiation occur independently in each population, leading to the origin of new species.
Footnotes
Acknowledgments
The authors are grateful to P.L. Gallo and L.H. da Silva for the collecting effort, to the Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis (IBAMA/MMA - number licenses 10538–1 and 38532–2) and Instituto Estadual de Florestas (IEF/MG) for the authorization to collect the material. This study was financed by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), and Fundação Araucária de Apoio ao Desenvolvimento Científico e Tecnológico do Estado do Paraná (Fundação Araucária).
Disclosure Statement
No competing financial interests exist.