
Abstract
Abstract
Some species of Characiformes are known for their high economic value, such as Colossoma macropomum and Piaractus mesopotamicus, and are used in aquaculture programs to generate hybrid tambacu (interbreeding of C. macropomum females and P. mesopotamicus males). The present work aimed to investigate the location of the Rex3 and Rex6 transposable elements in the hybrid and in the species, in addition to checking the genomic organization of the 18S and 5S rDNA in tambacu. The diploid number found for the hybrid was equal to 54 chromosomes, with heterochromatic blocks distributed mainly in the centromeric portions. The chromosomal location of the mobile elements Rex3 and Rex6 in C. macropomum, P. mesopotamicus, and in the hybrid between these species enabled knowledge expansion and the generation of data on such mobile elements. In addition, the location of such elements is not related to the distribution of ribosomal DNA sites. The mapping of the 18S rDNA was shown to be effective in cytogenetic identification of the hybrid tambacu, allowing for differentiation from the parent species and from the hybrid between C. macropomum and the other species from Piaractus (P. brachypomus).
Introduction
T
There have been cytogenetic analyses on C. macropomum, P. mesopotamicus, P. brachypomus, and their hybrids.5,7–9 Interspecific hybrids are important models for cytogenetic studies because they inherit one chromosome set of each parental species, which allows identification of divergences between the chromosomes of parental species.10–12 However, there are few analyses involving molecular cytogenetics in this fish group, and most of them are karyotypic descriptions.
Cytogenetically, 5S and 18S repetitive sequences from ribosomal DNA are the most studied and have gained in prominence mainly in studies on the evolutionary relationships between species and in the characterization of populations and cytogenetic markers as well as genome structure.13–15 Transposable elements (TE) are other sequences widely studied for chromosomal mapping in this group of fish.16–18 Studies of the isolation and mapping of repetitive sequences (18S rDNA, 5S rDNA, and TE) have been performed in different fish species of commercial interest, 19–29 including species of family Serrasalmidae such as the tambaqui.8,9
Retrotransposons are a class of TE that use the reverse transcriptase enzyme to “jump in DNA” via an intermediate RNA.16,17,29 This group contains the Rex3 and Rex6 elements that were first isolated from Xiphophorus maculatus.16,30 The transposition mechanism of these elements within the host genome is not random and is mediated by host cell signals as well as external factors such as thermal shocks, attack of parasites, stress, or self-regulation. The endogenous element restricts its movement according to its abundance in the genome.31–34 The evolutionary dynamics of retroelements in the fish genome has revealed a subtle association between its distribution and lineage differentiation within the group.16,30 The knowledge of the composition and distribution of these elements in the genome of species with commercial interest such as tambaqui and pacu is essential to elucidate the evolutionary trends covered by these species along the evolutionary process.
Thus, the aim of the present work was to map the distribution of retrotransposable elements Rex3 and Rex6 as well as rDNA genes in hybrid tambacu. We then analyzed the distribution of these sequences in parental C. macropomum and P. mesopotamicus and correlated the location of such mobile sequences with the 5S and 18S rDNA sequences as well as the heterochromatic blocks in tambacu.
Materials and Methods
In this work, 17 specimens of C. macropomum, 16 of P. mesopotamicus, and 7 of tambacu (interbreeding of C. macropomum females and P. mesopotamicus males) were analyzed—all from fish farms. The specimens were anesthetized with eugenol (5 mL in 12 L of water) to obtain chromosomal preparations according to the protocol by Bertollo et al. 35 The constitutive heterochromatin was detected with the C-band technique. 36
The extraction of total DNA was made from muscle tissue samples of C. macropomum, 37 and quantification used agarose gel and comparison to a standard lambda marker. The amplifications of the 18S and 5S rDNA genes were performed by polymerase chain reaction (PCR) using the primers 18Sf (5′-CCG CTT TGG TGA CTC TTG AT-3′) and 18Sr (5′-CCG AGGACC TCA CTA AAC CA-3′), 38 5Sa (5′-TAC GCC CGA TCT CGT CCG ATC-3′) and 5Sb (5′- CAGGCT GGT ATG GCC GTA AGC-3′), 39 respectively. For the amplification of the TE, we used the primers RTX3-F3 (5′-CGG TGA YAA AGG GCA GCC CTG-3′) and RTX3-R3 (5′-TGG CAG ACN GGG GTG GTG GT-3′) 40 as well as Rex6-Medf1 (5′-TAA AGC ATA CAT GGA GCG CCA C-3′) and Rex6-Medr2 (5′-GGT CCT CTA CCA GAG GCC TGG G-3′). 41
All PCRs were performed in a final volume of 25 μL containing genomic DNA (200 ng), buffer 10 × with 1.5 mM MgCl2, Taq DNA polymerase (5 U/μL), dNTPs (1 mM), a pair of primers (5 mM), and Milli-Q water. The profile of reaction for 18S rDNA was 1 min at 95°C, 35 cycles of 1 min at 94°C, 1 min at 56°C, and 1 min 30 s at 72°C. This was followed by 5 min at 72°C. The conditions for amplification of the 5S rDNA were 1 min at 95°C followed by 30 cycles of 1 min at 94°C, 1 min at 59°C, and 1 min 30 s at 72°C. The final extension was 5 min at 72°C. To amplify Rex3 and Rex6 elements, we used 95°C for 5 min, 35 cycles of 95°C for 1 min, 55°C for 40 s, and 72°C for 2 min. This was followed by a final extension of 72°C for 5 min. The PCR products were checked in 1% agarose gel quantified in a NanoVue Plus spectrophotometer (GE Healthcare). We used the probes for fluorescence in situ hybridization (FISH).
Homologous and heterologous hybridizations were carried out according to the protocol described by Pinkel et al. 42 at high stringency of 77% (2.5 ng/μL of 18S rDNA, 5S DNA, Rex3, and Rex6, 50% formamide, 10% dextran sulfate, and 2 × SSC at 37°C for 18 h). The chromosomes were stained with DAPI (2 mg/mL) in mounting medium Vectashield (Vector). The metaphases with FISH were analyzed in an epifluorescence microscope Olympus BX51, and the images were captured with an attached digital camera (Olympus DP71) using Image-Pro MC 6.3 software. The metaphases were processed with Adobe Photoshop CS3, and the chromosomes were measured with ImageJ and sorted according to the nomenclature proposed by Levan et al. 43
Results
The diploid number observed for the tambacu was equal to 54 chromosomes, including 27 metacentric and 27 submetacentric chromosomes (Fig. 1a). The karyotype was organized by chromosome type and in descending order of size because chromosomes do not form homologous pairs. In some cases, there is a great morphological similarity. The distribution of constitutive heterochromatin took place mainly in the centromeric and terminal portions of some complement chromosomes—especially chromosome numbers 21, 32, 35, 41, and 46. In chromosome 21, we noted a heterochromatic block at the end of the long arm, while in the chromosomes 32, 35, 41, and 46 there was a big heterochromatic block in the short arm (Fig. 1b). Besides heterochromatic blocks, the pericentromeric regions were seen in chromosomes 13, 29, 50, and 53.

Tambacu karyotype:
The mapping of the 5S and 18S rDNA ribosomal genes resulted in multiple markings in the hybrid as noted in the parental species. 9 The 18S rDNA was located in the end portion of five chromosomes (metacentric and submetacentrics) and is probably chromosomes 4, 5, 6 10, and 12. Moreover, the 5S sites were observed in interstitial position in the short arms of two chromosomes and in the long arms of two others (Fig. 2).
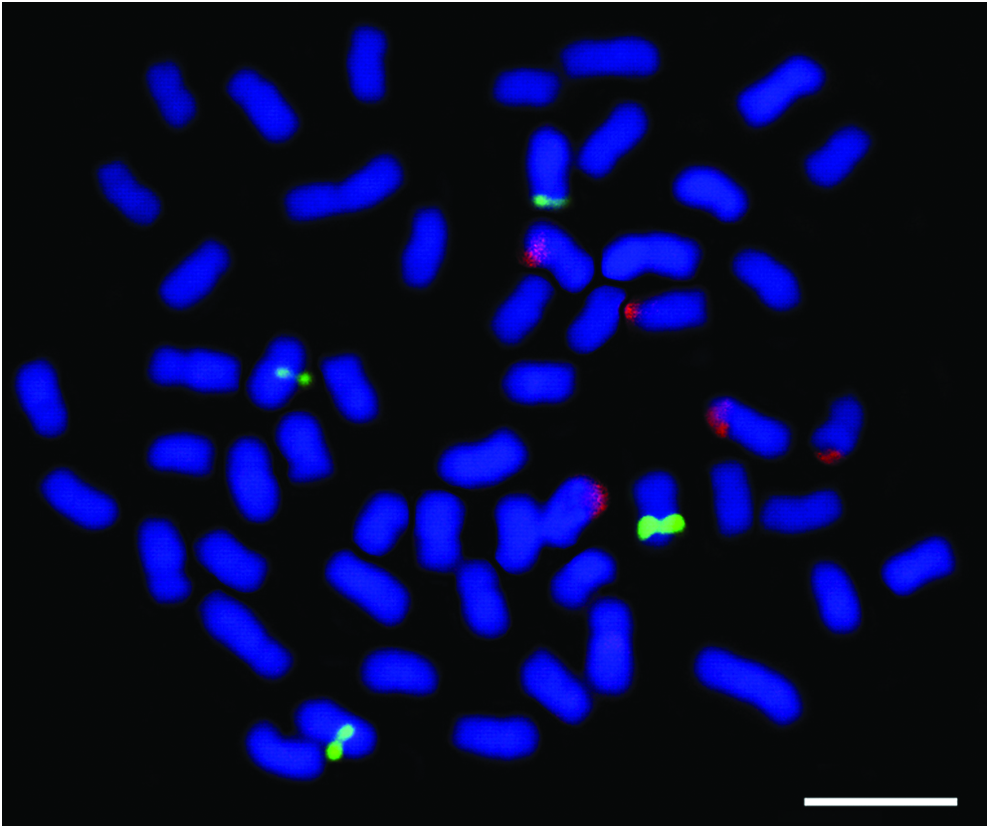
Tambacu metaphase showing 18S rDNA (red) and 5S rDNA (green) mapping. Bar: 10 μm. Color images available online at www.liebertpub.com/zeb
With respect to mapping the Rex3 and Rex6 retrotransposons, only low-intensity markings were observed and dispersed in heterochromatic and euchromatic portions. This showed that there is no preferential location of these sequences in the animals. In C. macropomum, we noted a few markings for Rex3. This was diffuse in several chromosomes. The Rex6 markings were more intense, but they were also scattered in several chromosomes (Fig. 3a). More Rex3 was observed in P. mesopotamicus than Rex6. This second element showed scattered distributions in some chromosomes and in a smaller quantity (Fig. 3b). The Rex3 element has an interstitial and dispersed distribution in several chromosomes of C. macropomum and P. mesopotamicus.
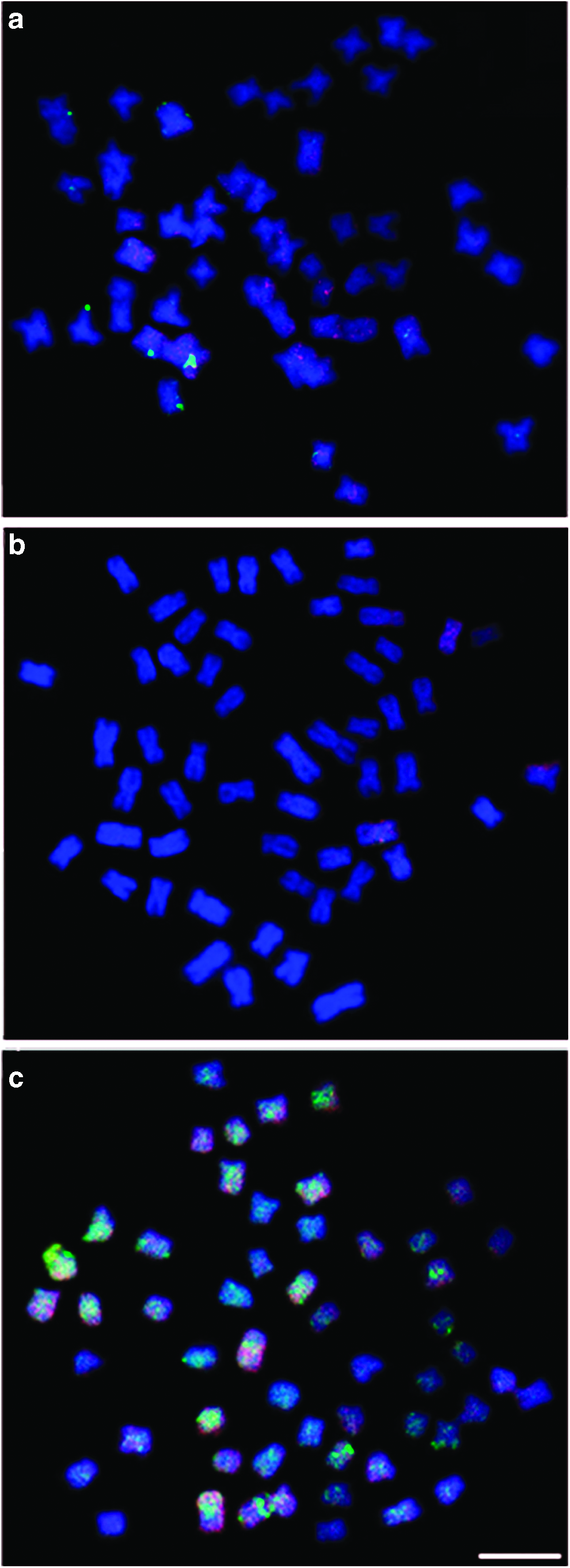
Metaphases showing Rex3 (red) and Rex6 (green) retrotransposable element distribution.
Discussion
Previous studies involving hybrids of other species show that they may present karyotypes very similar to the parental,44–46 as observed here. The C. macropomum and P. mesopotamicus, as well as their hybrids, have karyotypes comprising 54 chromosomes meta- and submetacentric. 9 However, subtle differences regarding the amount and distribution of heterochromatin were observed versus cytogenetic descriptions before tambacu hybrids. The first relates to the absence of chromosomal markers and lower amounts of constitutive heterochromatin.
Almeida-Toledo et al. 7 showed big heterochromatic blocks on centromeric portions, on the short arms of some chromosomes, and smaller blocks in terminal position. In this study, heterochromatic blocks were observed only in the centromeric portion. These results can be explained by the influence of several factors, including environmental factors, because epigenetic variations due to hybridization processes or environmental stresses can be inherited throughout generations. 47
The mapping of 18S rDNA was useful to distinguish between hybrids and pure species. C. macropomum has six chromosomes carrying 18S rDNA, while P. mesopotamicus presents only four. 9 The tambacu, in turn, has five marked chromosomes by FISH with 18S rDNA probe. This result confirmed that all individuals were hybrids and that each chromosome with 18S rDNA site was inherited from one specific parental species. In situ hybridization in tambacu indicated that the 18S ribosomal genes were not lost or altered. This was also seen in other interspecific hybrids of Leporinus and Pseudoplatystoma genus.11,48 The location of 18S rDNA allowed the differentiation of hybrids resulting from the interbreeding between C. macropomum and P. mesopotamicus and from hybrids from the interbreeding between C. macropomum and P. brachypomus. These are present in smaller numbers of 18S rDNA sites in different chromosomal pairs. 5
Although hybridization is a common procedure, the hybrids exhibit a potential biological hazard for the environment. If they are fertile, they can contaminate genetically natural stocks and breeding grounds and can compete with the parental species. 10 A high degree of homology in the chromosomal constitution of the parental species frequently results in correct pairing and segregation during meiosis as observed in other fish 48 and in this study. The distribution of Rex3 element was similar to that described for other species of order Characiformes such as Erythrinus erythrinus, 49 Brycon amazonicus, and Brycon orbignyanus. 28 For Rex6, the less frequently studied element in teleosts and the most restricted occurrence is absent in several species. 28 This work shows the first chromosomal location of this element for the order Characiformes. Although less abundant, the scattered distribution pattern of Rex6 was observed in the parent species C. macropomum and P. mesopotamicus, and this matches the pattern described for species of the order Perciformes, including Astatotilapia latifasciata, Chaetobranchus flavescens, Oreochromis niloticus, and Satanoperca jurupari. 50
For Rex3 and Rex6, fluorescent signals did not have great intensity in the examined individuals; however, the distribution of such elements was clearly seen in several chromosomes. The most evident markings of Rex6 in C. macropomum can be associated with a greater quantity of these elements that are located closely. In addition, this diffuse distribution pattern may be because these TEs have structural and regulatory functions that justify their dispersion in both types of chromatin. The accumulation of TE has great importance in regulatory functions of the genome and in terms of targeting gene expression.51,52
The association of Rex3 and Rex6 sequences with 5S and 18S rDNA was not observed in tambacu. Hybrids have also been observed to colocalize the retroelement Rex3 and 5S rDNA. This TE is related to the dispersion of 5S rDNA and unrelated to 45S rDNA. 53 In tambacu, no relationship was seen between the mapped retroelements and the ribosomal sequences because there was no increase in the site number of 5S and 18S rDNA in the analyzed animals. This maintained the pattern described for the parent species. 9
On the contrary, there is an increase in the amount of retrotransposons in the hybrid tambacu probably due to the need for adjustments in the cell division processes. The hybridization process is known to induce TE transposition because it changes the TE control system and requires a reorganization of the entire genome while modifying the recombination patterns. One of the explanations for the origin and dispersion of mobile elements is the introgressive hybridization involving parent species.54,55 After the origin of TE, a rapid dispersion occurs with an increase in the number of these mobile sequences in the host genome, which leads to a diversity of insertion sites.55–57 Generally, the junction of two different genomes caused by the hybridization process leads to several consequences such as changes in the control of gene expression, DNA methylation, chromosome rearrangements, and mobilization of TE. 58
In fish, the increase in Rex3 sites has been observed in natural hybrids of Cyprinidae, 53 which presented a higher amount of Rex3 clusters with independent distribution in heterochromatic and euchromatic portions. There are a few differences between homologous chromosomes as seen here.
It is possible that the TE markings observed in tambacu represent a mechanism of hybrid dysgenesis,59,60 with a propagation of Rex elements that can be active and related with gene regulation processes. This would explain the large increase in the quantity of Rex3 and Rex6 markings compared to the parent species. However, analyses involving the gene expression of these elements are necessary to validate this hypothesis.
We conclude that the 18S rDNA is a cytological marker effective for monitoring the cultivation of these fish, and that the chromosomal location of Rex3 and Rex6 mobile elements in C. macropomum, P. mesopotamicus, and tambacu is important in the expansion of knowledge and the generation of data on such mobile elements. Accordingly, studies of identification, characterization, and genetic monitoring of hybrids are required to achieve sustainable use of these fishery resources without prejudice to the wild species and the natural environment. Furthermore, the development of genetic techniques that allow the correct manipulation and making of diagnostic markers of stocks is a promising area for the expansion of Amazon fish farming.
Footnotes
Acknowledgments
The authors are grateful to Alexandre Honczaryk for support and access to the Santo Antonio (AM) farm, José A. Esteves and Antônio C. Diniz (CETaq - Technical Education Center of Monte Aprazível). This work was supported by the Conselho Nacional de Pesquisa e Desenvolvimento Tecnológico (CNPq—475943/2009-0), the Instituto Nacional de Pesquisas da Amazônia/Genética, Conservação e Biologia Evolutiva (INPA/GCBEv), the Fundação de Amparo de Pesquisas do Amazonas (PRONEX FAPEAM/CNPq and FIXAM/FAPEAM process number 1828/2013), and the Centre for Studies of Adaptation to Environmental Changes in the Amazon (INCT ADAPTA, FAPEAM/CNPq 573976/2008-2). This manuscript was edited for proper English language, grammar, punctuation, and spelling by the expert staff at American Manuscript Editors (project number 13178/certificate verification key 900-572-786-138-667).
Disclosure Statement
No competing financial interests exist.