
Abstract
Abstract
Various species and lineages that until recently were identified as Microsternarchus bilineatus (Hypopomidae, Gymnotiformes) have a widespread distribution in the Amazon and Orinoco River basins and across the Guiana shield. Recent molecular studies show five distinct lineages for Microsternarchus from different localities. These results suggest that this previously monotypic genus actually consists of more than one species. Here, we describe the karyotype of M. aff. bilineatus from the Cururutuia River (Bragança, Pará, Brazil). The diploid number of 48 chromosomes (14 meta-submetacentric/34 subtelo-acrocentric) is found for males and females, with an XX/XY sex chromosome system. The nucleolar organizer region is found in the short arm of pair 9. Constitutive heterochromatin occurs in the pericentromeric region of all chromosomes, in the distal region of 3p, 5p, 7p, 8q, 9q, 16q, and Xq, in the interstitial region in 2p, 10q, 11q, and 12q and all along 4p, and in a large block of the Y chromosome. These results indicate extensive karyotype divergence between this population and samples from Igarapé Tarumã Grande (Negro River, Amazonas, Brazil) studied by other researchers. Moreover, despite the diversity of sex chromosome systems found in Gymnotiformes, the XX/XY sex chromosome system of M. aff. bilineatus is the first case of Y chromosome degeneration in this order. The present data are valuable to help understand karyotype evolution in Hypopomidae.
Introduction
T
One aspect that has been extensively explored in this group is its karyotype diversity. Besides sporadic cases of polyploidy, great intra- and interspecific variation has been found in the number and morphology of the chromosomes, localization of the NOR (Nucleolar Organizing Region), occurrence of supernumerary chromosomes, and sex chromosome systems.4–25 These data have been extensively used in the identification of populations and/or species and to understand the process of evolution in this group.9,13,26. Moreover, the wide diversity of sex chromosome systems makes this group an interesting model to explore the mechanism of sex chromosome evolution.19,22
The phylogenetic relationships of Gymnotiformes have been revised several times, but are well resolved only at the family level.1,27–29 The families Hypopomidae and Rhamphichthyidae are closely related and constitute the superfamily Rhamphichthyoidea.1,28,30,31 According to recent phylogenies Akawai openak is basal among the Hypopomidae and is a sister group to the clade that includes both the Microsternarchinae and Brachyhypopominae31–33 (Fig. 1).

Among the Hypopomidae, cytogenetic data are known for Hypopomus artedi, 34 four species of Brachyhypopomus,7,22,24,34 and a population of Microsternarchus bilineatus from Igarapé Tarumã Grande (a left margin tributary of the Negro River, Amazonas, Brazil). 35
The genus Microsternarchus has until recently been considered as monotypic, with the type locality of M. bilineatus in the San Jose River, Venezuela 36 (Fig. 2). The genus is distributed across the Amazonas and Orinoco River Basins and the Guiana shield. Molecular studies with mitochondrial DNA (Cytochrome oxidase I) in Microsternarchus from different Amazonian localities demonstrate at least five distinct lineages, suggesting the existence of new species. 37 A new genus and species, Procerusternarchus pixuna 33 and a new species Microsternarchus brevis, 38 both from the Negro River were traced from these five lineages. As type specimens of M. bilineatus are currently lost, a population from the confluence of the Casiquiare and Orinoco Rivers has been assumed to represent the original species based on original measurements and proximity to the type locality. 38 However, cytogenetic data in the genus are known only for a population named as M. bilineatus 37 from the Negro River. 35 Therefore, as chromosome data are valuable for diagnosing many populations and/or species of Gymnotiformes, it is relevant to describe the karyotypes of other populations of Microsternarchus, to help resolve taxonomic questions in this group.

In the present work we describe the karyotype of a population of M. aff. bilineatus from the Cururutuia River, Pará State, to provide useful information that will help the understanding of mechanisms of karyotype evolution in the Hypopomidae and Gymnotiformes in general.
Materials and Methods
We analyzed 15 specimens (6 males and 9 females, lot UFPACIT-P-L00001, UFPA Cytogenetic reference collection [UFPACIT]—a registered collection for genetic patrimony in Brazil) of M. aff. bilineatus from the Cururutuia River (Bragança, PA, Brazil: S01°04′42.0"/W046°44′18.6"; Fig. 2). We used the classification M. aff. bilineatus due to the distance from the type locality of M. bilineatus and the existence of multiple divergent molecular lineages, and the impossibility of association of our sample to any published lineage. Samples were sequenced for a CO1 gene fragment following the protocols described in Cardoso et al. 39 and submitted to BOLD under the project Barcoding and Cytogenetics of Gymnotiformes (BCG). Within the BOLD species identification tree (Supplementary Data; Supplementary Data are available online at www.liebertpub.com/zeb) our sample clearly falls within a lineage delimited as Microsternarchus sp. rather than with Procerusternarchus or M. brevis. However, despite the authors of Maia and Alves-Gomes 37 described the five lineages using barcoding, they never deposited those data, turning impossible to do any comparison. Field collection was authorized by Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio), permit 020/2005 (IBAMA Registration: 207419) and the taxonomic identification of these samples was confirmed by Dr. William G.R. Crampton. Eugenol was used to anesthetize the animals following euthanasia guidelines approved by the Federal University of Pará Committee for the Ethical Use of Animals (CEUA). Metaphase chromosome preparations were obtained from kidney cells 40 and analyzed by conventional staining, C-banding, 41 silver staining for NOR with AgNO3, 42 and fluorescence in situ hybridization with probes of 45S rRNA genes. Chromosomes have been classified according to Levan et al. 43
Results
All the specimens of M. aff. bilineatus have 2n (diploid number) = 48 chromosomes. The karyotype consists of 14 metacentric/submetacentric (m/sm) plus 34 subtelocentric/acrocentric (st-a) chromosomes and the FN (fundamental number) = 62, with m/sm considered bi-armed and st-a as one-armed. Heteromorphism of size related to the male sex was detected in a chromosome pair, which indicates the presence of a XX/XY sex chromosome system (Fig. 3A, B). Both are subtelocentric/acrocentric chromosomes, but the X is larger than Y chromosome. Both chromosomes have heterochromatic short arms. However, the Y chromosome shows an exclusive heterocromatic block that extends from the centromeric region to the middle region of the long arm.
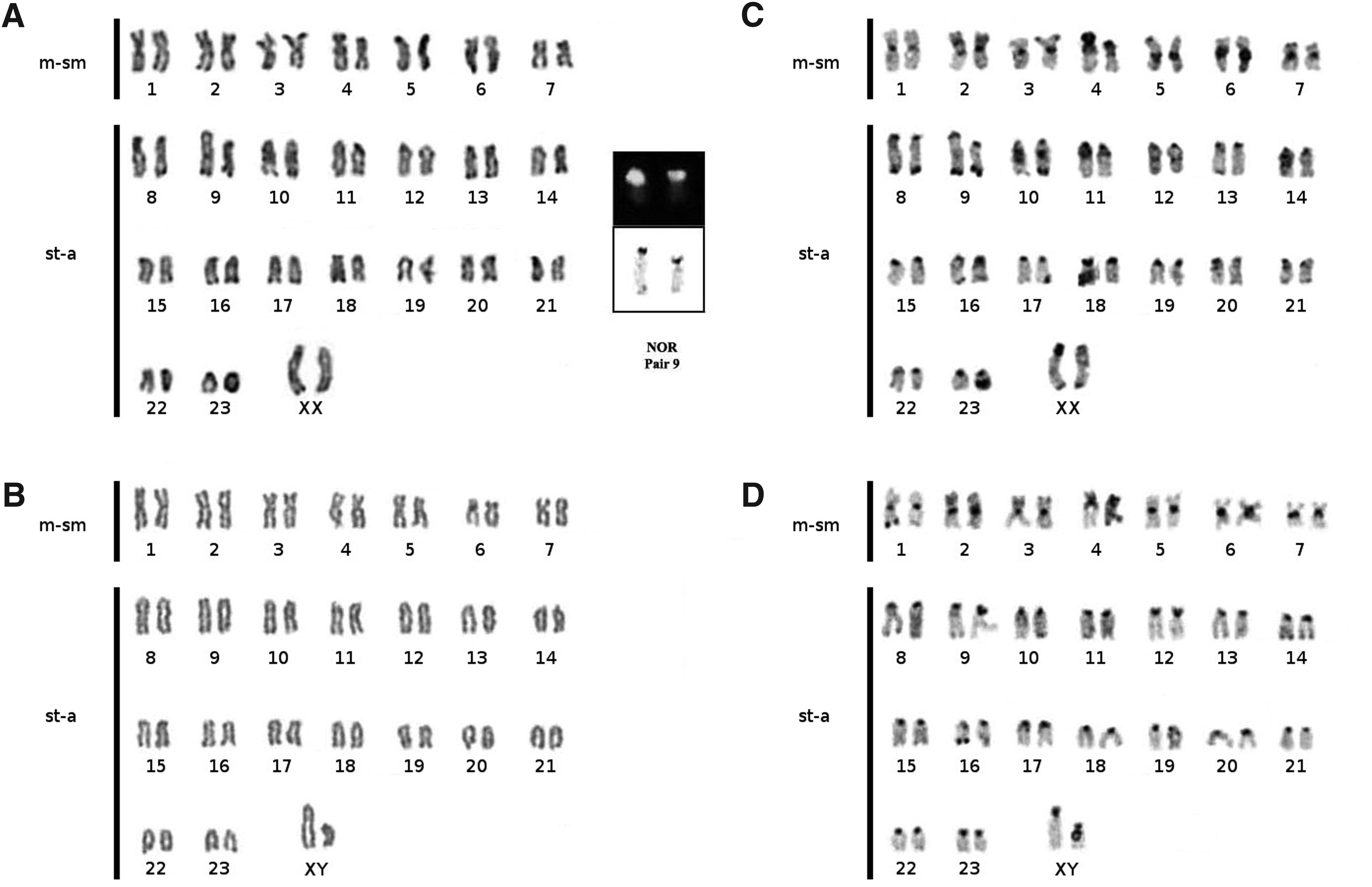
Karyotype of M. bilineatus: conventional Giemsa staining in female
NOR/45S rDNA sites were found in the short arm of chromosome pair 9 (Fig. 3, box), which shows size heteromorphism. Constitutive heterochromatin was detected in the pericentromeric region of all chromosomes, in the distal region of 3p, 5p, 7p, 8q, 9q, 16q, and Xq, in the interstitial region in 2p, 10q, 11q, and 12q and all along 4p. The Y chromosome is almost entirely heterochromatic (Fig. 3C, D). Pair 4 shows size heteromorphism due to difference of the heterochromatic block in the short arm between the homologs.
Discussion
Sex chromosomes and mechanisms of differentiation
Male heterogametic sex chromosome systems are common among Neotropical electric fish, covering more than 70% of the systems described in this group (Table 1). Our data demonstrate a XX/XY sex chromosome system in our sample of M. aff. bilineatus. The large heterochromatic block extending from the centromeric to the interstitial region of the Y chromosome, which is absent in the X chromosome, indicates events of heterochromatinization. This mechanism is often described during the evolution of sex chromosomes and results from the absence of recombination in rearranged regions. 44 It has been observed in the evolution of several types of sex chromosomes of Gymnotiformes, including X, Y, and W chromosomes.5,17–19,21
SCS, sex chromosome system.
Apart from heterochromatinization, we also find that the Y of M. aff. bilineatus studied here is smaller than the X chromosome. This indicates it has been degenerating, probably by gene loss, as is frequent in Y evolution. 45 Among the XY sex chromosome systems identified in Gymnotiformes this is the only case of Y chromosome degeneration (Fig. 4). Interestingly, in other species of Gymnotiformes with XY sex systems, the X is reduced, as clearly observed in Gymnotus carapo (Catalão Lake) and Gymnotus sp. (Negro River). 19 In Eigenmannia virescens and Eigenmannia sp. B the X chromosome is under heterochromatinization that may be followed by size reduction.18,19 We therefore conclude that the sex chromosome system of M. aff. bilineatus from the Cururutuia River has reached an advanced level of differentiation.
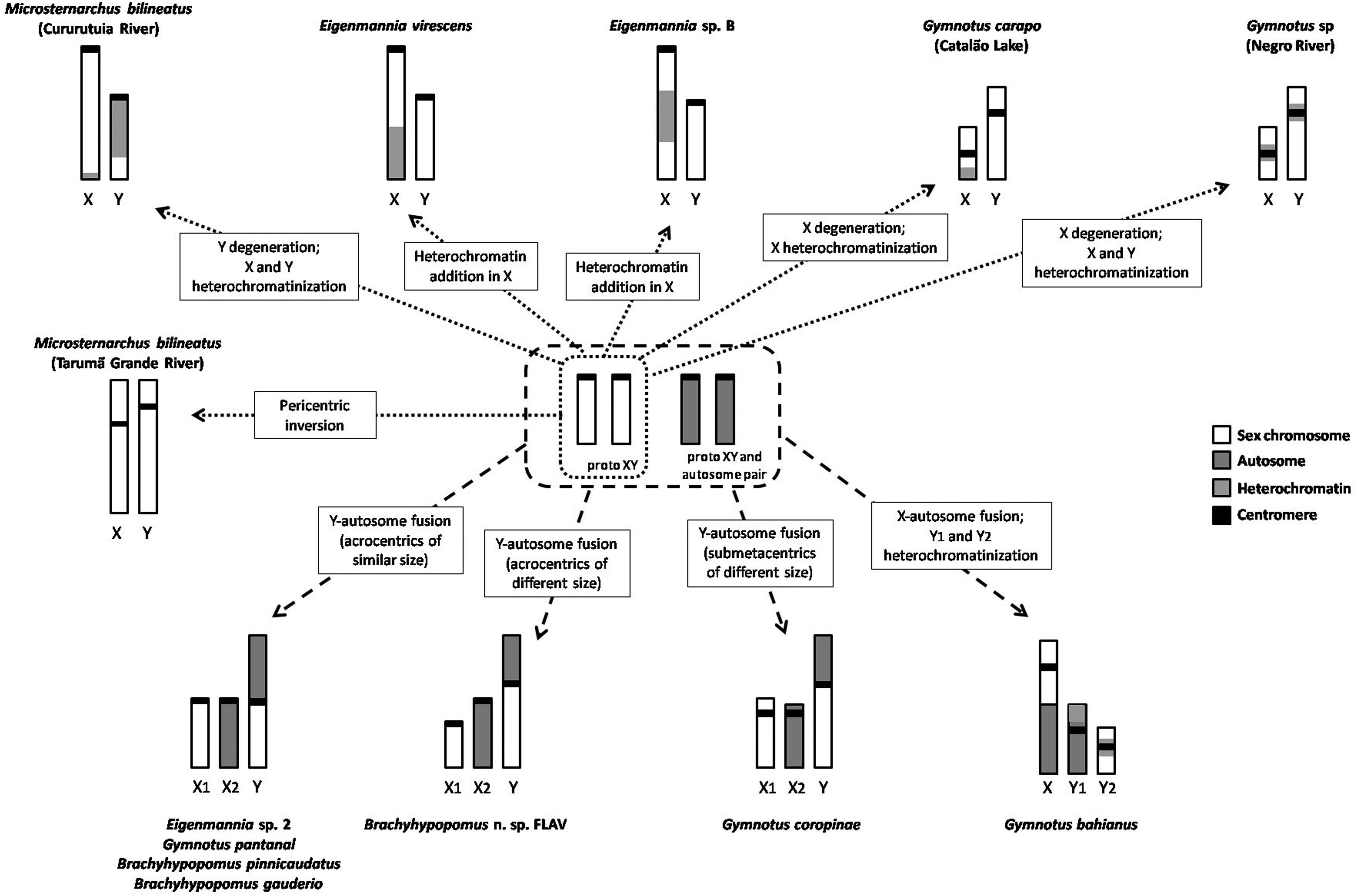
Mechanism of evolution of male heterogametic sex chromosome systems in Gymnotiformes.
In the population of M. bilineatus (lineage C) from Igarapé Tarumã Grande in the Negro River (Fig. 5), 35 a heteromorphic chromosome pair is observed in males. This is evidence of a simple sex chromosome system of the XX/XY type, which should have evolved from a pericentric inversion, since the X chromosome is subtelocentric and the Y chromosome is submetacentric. The sex chromosome system of the population from the Cururutuia River probably evolved independently and earlier than the population from Igarapé Tarumã Grande in the Negro River. Three species of Brachyhypopomus, another genus in the Hypopomidae along with Microsternarchus, show X1X2Y sex chromosome systems.5,22,24 If the XY system of M. bilineatus is common to all lineages and other Microsternarchinae, then it represents the status that gave rise to the X1X2Y systems of Brachyhypopomus. However, not all Brachyhypopomus have differentiated sex chromosomes, so it is more parsimonious to assume that sex chromosome systems of M. bilineatus do not have homeology with the proto-XY system that originated these X1X2Y systems in Brachyhypopomus. In fact, it is difficult to trace the trends of sex chromosome evolution in the Hypopomidae due to the absence of chromosome data for other species of this family. Anyway the emergence of male heterogametic sex chromosome systems could represent a trend in this family, contrasting with the emergence of ZW systems observed in the Rhamphichthyidae. 21

Comparison between ideograms of the M. bilineatus taxa from Negro River and Cururutuia River.
The present data increase the knowledge about the diversity of types of sex chromosome systems and their mechanisms of evolution in Gymnotiformes and reinforce the occurrence of an intense process of turnover of sex chromosomes in this order.19,22
Karyotype evolution in Hypopomidae
Among Gymnotiformes, the family Hypopomidae has cytogenetic data available only for few representatives of the Brachyhypopominae. Therefore, the karyotype description of lineages of M. bilineatus is useful to better understand the karyotype diversity and evolution in this family in the light of previous phylogenies. The population of M. aff. bilineatus studied here (2n = 48; 14 m-sm/34st-a) has the same diploid number (2n = 48; 21 m-sm/27st-a ♂, 20 m-sm/28st-a ♀) present in the population from the Negro River, 35 however, these populations are different in karyotype formula between males and females and in relation to the chromosomes bearing the NOR (multiple NOR in the Negro River vs. a simple NOR in the Cururutuia River) (Fig. 5). These data indicate that rearrangements such as pericentric inversions and translocations are involved in karyotype divergence, promoting changes to the chromosome morphology and localization of the NOR. Moreover, in respect to NOR sites, it is possible that mechanisms of transposition are involved in expansion of these sites in M. bilineatus (lineage C) from the Negro River, although events of degeneration may have happened in the M. aff. bilineatus population from the Cururutuia River. These results, together with Cytochrome oxidase I studies 37 and recent species descriptions33,38 strongly support the view of a significant overlooked diversity in what was previously considered a monotypic genus. However, more detailed taxonomic analyses will be necessary to clarify the real number and traits of the species of this genus.
While Rhamphichthyidae have an evolutionary history marked by chromosome inversions changing the morphology of the chromosomes20,21,24,34 the cytogenetic diversity observed in Hypopomidae indicates that karyotype evolution in this group was influenced by all the types of chromosome rearrangements that are able to modify the number and morphology of the chromosomes. Given this more prominent karyotype reorganization, it is challenging to identify chromosome homeologies and try to trace karyotype evolution in this group. Therefore, future work using methods that are able to detect chromosome homeologies need to be conducted and other species of this family need to be cytogenetically investigated (especially among the Microsternarchinae) aiming to understand their karyotype evolution.
Footnotes
Acknowledgments
This research was support by CNPq through the Edital Universal (Proc. 475013/2012-3) on a project coordinated by C.Y.N.; by FAPESPA (Pará Foundation for Supporting Science) through the National Excellence on Research Program (PRONEX, TO 011/2008) and Banco Nacional de Desenvolvimento Economico e Social—BNDES (Action 2.318.698.0001) on a project coordinated by J.C.P. C.Y.N. (308428/2013-7) and J.C.P. (308401/2013-1) are grateful to CNPq for Productivity Grants. We also thank the Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio) for the collection permit 020/2005 (Registration: 207419).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.