
Abstract
Abstract
Cytogenetic studies in fish of the Rineloricaria genus have already shown a high variation in diploid number (2n). Along with fusion/fission events for 2n alteration, inversions contribute to the diversification of chromosome formulae within this group. The present study assessed different populations/species of the Rineloricaria aiming to describe the karyotype organization of its members and understand the mechanisms that lead to the variation of chromosome numbers. Cytogenetic data showed distinct karyotype organization among Rineloricaria populations/species studied, ranging in diploid number from 46 to 64 chromosomes, syntopic species and two karyomorphs in Rineloricaria pentamaculata. Using ribosomal DNAs (rDNAs) and TTAGGGn probes by fluorescence in situ hybridization in species with low diploid numbers, we detected sites of 18S rDNA, 5S rDNA, and TTAGGGn in centromeric regions of metacentric chromosomes, which participated in chromosome rearrangements like centric fusions. In species with high 2n, centric fissions probably occurred in karyotypic diversification. In this study, we assessed the telomeric instability, chromosomal breaks, and rearrangements due to interstitial telomeric site vestiges detection, in addition to the probable role of rDNAs in chromosome fusions in karyotypic diversification of this group.
Introduction
L
Loricariinae is the second largest subfamily in Loricariidae and most of its species present highly differentiated characteristics in relation to their ancestral characteristics. For example, the diploid number variation ranges from 30 to 70 chromosomes within the genus Rineloricaria6–8 and the presence of B chromosomes and heteromorphic sex chromosome systems in Harttia.9–12
The main chromosomal rearrangement involved in karyotypic diversification of Loricariinae are the events of centric fusion and fission,7,8,11,13 combined with inversions.7,14 Rineloricaria is an example of the karyotype plasticity observed in this group, and in addition to the high variation of the diploid number among the species, there are numeric polymorphisms in Rineloricaria latirostris (Boulenger, 1900) (2n = 36–48 chromosomes), Rineloricaria lima (Kner, 1853) (2n = 66–70), and Rineloricaria lanceolata (Gunther, 1868) (2n = 48–45 chromosomes).6–8 In R. lima, there were vestiges of interstitial telomeric sites (ITSs) and 5S rDNA in chromosomes fused, in addition to the different gametic combinations for the occurrence of different karyotype organizations. 7
In most cases, the karyotype organization is unique, as a reflex of the evolutionary history of this group, being a good method to differentiate the specimens. Thus, considering that some species of Loricariidae have high karyotypic diversity and repetitive DNAs are known for presenting hotspots for chromosomal break and rearrangement, this study aimed to analyze karyotypic diversity in different species/populations of Rineloricaria, using traditional cytogenetic analysis, chromosome markers of ribosomal DNAs, and TTAGGGn sequence to understand the probable chromosomal rearrangements within this group.
Materials and Methods
Eighty-one specimens of different species/populations of the Rineloricaria genus captured in different hydrographic basins of Paraná river system were cytogenetically analyzed: R. latirostris (Boulenger, 1900) (Laranjinha river); R. latirostris and R. pentamaculata (Langeani and de Araujo, 1994) (Barra Grande river); R. pentamaculata (Juruba river); R. stellata (Ghazzi, 2008) and R. capitonia (Ghazzi, 2008) (Uruguay river) with sample sites as shown in Figure 1. Sample data and species identification are reported in Table 1. Capture and transportation of fish specimens in nature were according to the license of Instituto Chico Mendes de Conservação da Biodiversidade (permanent license for collection of zoological material ICMBio/SISBIO: 15117-1).
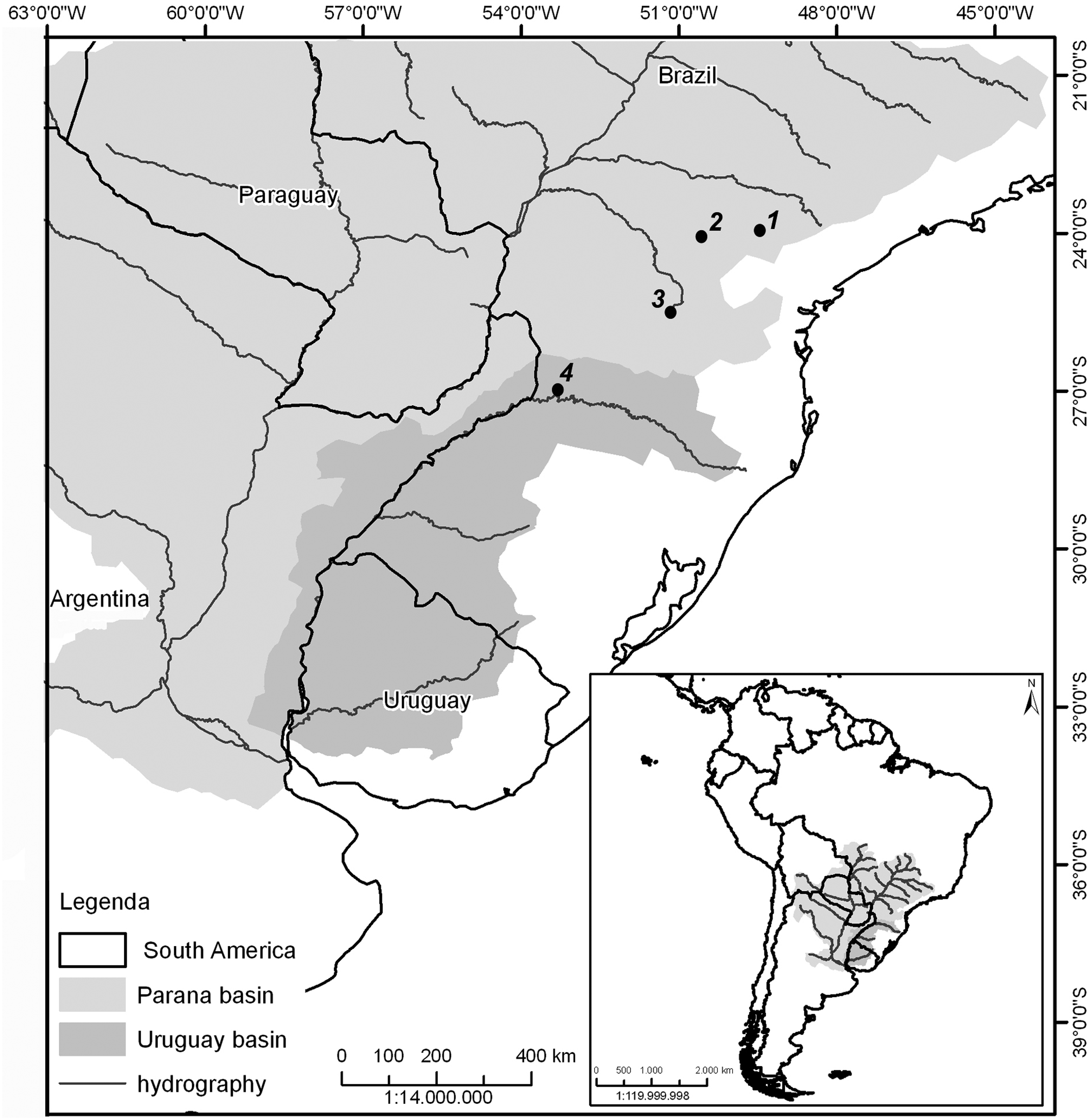
Partial map of South America, highlighting Paraná river basin, with the collecting points of Rineloricaria populations. (
Species identification was accomplished by Dr. Cláudio Henrique Zawadzki and deposited in the Icthiology Museum “Museu de Zoologia do Núcleo de Pesquisas em Limnologia, Ictiologia e Aquicultura da Universidade Estadual de Maringá (NUPELIA)” under number NUP 17120–17123 and 17140–17141.
♂, number of males; ♀, number of females; GPS, global position system; PR, Paraná; SC, Santa Catarina.
Procedures for animal care in research were according to the Ethics Committee of Animal Usage (process CEUA 007/2014) of Universidade Estadual de Ponta Grossa and Brazilian laws. The mitotic chromosomes were obtained following the methods described by Blanco et al. 15 Detection of the constitutive heterochromatin was accomplished according to Sumner. 16
Fluorescence in situ hybridization (FISH) was performed following Pinkel et al. 17 instructions, using 18S rDNA 18 and 5S rDNA probes, 19 and the general telomeric sequence (TTAGGG)n of vertebrates. 20 For amplification of the telomeric sequence, two amplification conditions were used. The first one was performed with low stringency (4 min at 94°C, 12 cycles of 1 min at 94°C, 45 s at 52°C, and 1 min 30 s at 72°C), followed by 35 cycles of high stringency (1 min at 94°C, 1 min 30 s at 60°C, and 1 min 30 s at 72°C). The 18S rDNA and the telomeric sequence probes were labeled with biotin through the nick translation technique, using the biotin 16-2′-deoxyuridine-5′-triphosphate (16-dUTP) compound (Nick Translation Biotin; Roche Applied Science); whereas 5S rDNA probe was labeled with digoxigenin through the nick translation technique, using the digoxigenin 11-dUTP compound (DIG Nick Translation Mix; Roche Applied Science). All hybridization mixtures had an approximate stringency of 77% (1 μg of each probe, 50% deionized formamide, 10% dextran sulfate, 2 × saline-sodium citrate, 37°C for 16 h). For probe detection, we used antibodies Streptavidin Alexa Fluor 488 (Molecular Probes) and anti-digoxigenin rhodamine (Roche Applied Science). Chromosomes were counterstained with 4′,6-diamidino-2-phenylindole (DAPI 0.2 μg mL−1) in mounting medium VECTASHIELD (Vector) and analyzed under epifluorescence microscope Zeiss Axio Imager A2 coupled with the ZEISS pro 2011 software in Zeiss AxioCam MRm camera of monochromatic capture with CCD sensor and 1.4 megapixel resolution (Carl Zeiss®).
For karyotype assembly, homologous chromosomes were paired and divided into metacentric (m), submetacentric (sm), and subtelocentric/acrocentric (st/a) according to the arm ratio. 21 To establish the fundamental number (FN), we considered the chromosomes st/a as a single chromosome arm, as previously determined for Loricariidae. To settle the number of FISH marks, ∼30 cells with chromosomes in metaphase were analyzed.
Results
Giemsa staining and C-banding
We obtained cytogenetic data for four species morphologically identified (Supplementary Table S1; Supplementary Data are available online at www.liebertpub.com/zeb). For the populations of R. latirostris collected from the Laranjinha and Barra Grande rivers, 2n = 46 chromosomes was observed, subdivided in 10 m + 4 sm +32 st/a and FN = 60 (Fig. 2a, b and Supplementary Fig. S1a, b). In the Barra Grande river, by collecting sympatric and syntopic specimens to R. latirostris, we captured two karyomorphs of R. pentamaculata species. The karyomorph A of R. pentamaculata presented 2n = 56 chromosomes, karyotype formula 4 m + 10 sm +42 st/a, and FN = 70 (Fig. 2c and Supplementary Fig. S1c). The karyomorph B of R. pentamaculata presented 2n = 54 chromosomes subdivided in 6 m + 4 sm +44 st/a and FN = 64 (Fig. 2d and Supplementary Fig. S1d). R. pentamaculata from the Juruba river presented 2n = 56 chromosomes, karyotype formula 4 m + 10 sm +42 st/a and FN = 70 (Fig. 2e and Supplementary Fig. S1e). R. stellata from the Uruguay river presented 2n = 54 chromosomes subdivided in 6 m + 14 sm +34 st/a, FN = 74 (Fig. 2f and Supplementary Fig. S1f). Also captured in Uruguay river, R. capitonia specimen showed 2n = 64 chromosomes, karyotype formula 4 m + 2 sm +58 st/a, and FN = 70 (Fig. 2g and Supplementary Fig. S1g).

Karyotypes of Rineloricaria populations/species submitted to C-band.
C-banding showed the heterochromatin located preferentially on the pericentromeric region of all chromosomes for both R. latirostris populations, except for the m chromosome pair 2 of the Laranjinha river specimens, the block of which was larger (Fig. 2a, b). Karyomorph A of the R. pentamaculata revealed small pericentromeric blocks in chromosomes, in addition to the short arm of sm chromosome pair 3, which is entirely heterochromatic (Fig. 2c). C-banding of the karyomorph B showed evident blocks of heterochromatin in the centromeric regions, as well as the short arm of sm chromosome pair 4, which is entirely heterochromatic (Fig. 2d). R. pentamaculata from the Juruba river showed small pericentromeric blocks, in addition to the short arm of sm chromosome pair 3, which is entirely heterochromatic (Fig. 2e). C-banding showed the heterochromatin located preferentially on the small pericentromeric heterochromatic blocks in R. stellata (Fig. 2f). In R. capitonia were visualized pericentromeric heterochromatin blocks on the chromosomes, and the whole short arm of sm chromosome pair 3 was heterochromatic (Fig. 2g).
Ribosomal DNA sites and TTAGGGn sequence location
FISH mapping of 5S rDNA probe in chromosomes of R. latirostris population from the Laranjinha river showed six pairs marked, with m chromosome pair 3 in the centromeric region and five st/a pairs (10, 11, 21, 22, and 23) in the terminal region (Fig. 3a). The population of R. latirostris from the Barra Grande river presented four pairs carrying 5S rDNA, one m pair (pair 3) in the centromeric region, and three st/a pairs (10, 11, and 21) in the terminal region (Fig. 3b). The 18S rDNA site was located in the pericentromeric region of m chromosome pair 2 in both R. latirostris populations (Fig. 3a, b, in details). The karyomorph A of R. pentamaculata from the Barra Grande river had five acrocentric pairs (8, 9, 10, 15, and 16) with 5S rDNA sites in the terminal region (Fig. 3c), whereas karyomorph B presented four chromosome pairs (7, 8, 10, and 21) carrying 5S rDNA in the terminal region (Fig. 3d). R. pentamaculata from the Juruba river presented six acrocentric pairs (8, 9, 10, 11, 13, and 14) with 5S rDNA sites in the terminal region (Fig. 3e). Karyomorphs A and B of R. pentamaculata from the Barra Grande river and R. pentamaculata from the Juruba river presented 18S rDNA sites in the short arm of the major sm chromosome pair (Fig. 3c–e, in details). In the Uruguay river, R. stellata had five chromosome pairs (3, 8, 11, 13, and 15) carrying 5S rDNA (Fig. 3f), and four chromosome pairs (4, 6, 9, and 10) bearing 18S rDNA sites (Fig. 3f, in detail). R. capitonia presented four st/a pairs (16, 17, 18, and 19) bearing terminal 5S rDNA sites (Fig. 3g), and 18S rDNA site was located in the short arm of sm chromosome pair 3, which represents the variation in cistron size among the homologs (Fig. 3g, in detail).

Karyotypes of Rineloricaria populations/species submitted to FISH using 5S rDNA, 18S rDNA, and TTAGGGn probes.
FISH mapping of TTAGGGn sequence showed telomeres marked as well as ITS vestiges in species of R. latirostris and karyomorph B of R. pentamaculata (Fig. 3, in box). Populations of R. latirostris (Laranjinha and Barra Grande rivers) evidenced two m chromosome pairs (1 and 3) with vestiges of ITS, with one pair colocated with the 5S rDNA (Fig. 3h, i). Karyomorph B of R. pentamaculata from the Barra Grande river also presented ITS vestiges in the centromeric region of the m pair 1 (Fig. 3j). The other species studied presented only terminal sites for the telomeric location (data not shown).
Discussion
Loricariidae is known for presenting 2n = 54 chromosomes as an ancestral characteristic.2,22 Among subfamilies of Loricariidae, Loricariinae has the major range of diploid number: from 2n = 36 to 74 chromosomes, 3 and only the genus Rineloricaria presents a variation from 2n = 36 to 70 chromosomes. 13 In this study, we detected a variation from 2n = 46 chromosomes in populations of R. latirostris to 2n = 64 chromosomes in R. capitonia. These results corroborate the proposition that events of Robertsonian fusions/fissions and pericentric inversions are the main processes of karyotypic diversification observed in the Loricariinae and are responsible for the high variability of numerical and structural changes.3,6–8 Moreover, repositioned centromeres are centromeres that move to a new position along the chromosome without any change in marker order and are now accepted as an important mechanism of genome evolution ranked on equal grounds with traditional chromosome rearrangements, such as inversions, translocation, deletions, and insertions. 23 However, little is known about the mechanisms that lead to chromosomal breaks and rearrangements in Loricariidae.
R. latirostris presents numeric polymorphisms and variation of diploid number among its populations ranging from 2n = 36 to 48 chromosomes. 6 Two populations of R. latirostris in this study (Laranjinha and Barra Grande rivers) had 2n = 46 chromosomes and the same karyotype organization. Even keeping the diploid number, the analysis of ribosomal DNAs (rDNAs) and TTAGGGn probes using FISH mapping indicated that Robertsonian fusion events occurred in the evolutionary history of this species. Rineloricaria species with diploid number superior to 54 chromosomes presented 18S rDNA in the terminal region of st chromosome pairs7,24,25 and 5S rDNA in the terminal region of acrocentric chromosomes. 7 In R. lima, it was detected 5S rDNA sites in the fusion point of metacentric chromosomes. 7 In populations of R. latirostris, the occurrence of one large m chromosome pair carrying 18S rDNA and one large m chromosome pair carrying 5S rDNA colocated with TTAGGGn sequences, in addition to a third large m chromosome pair carrying ITS vestiges, all with centromeric location, may indicate fusions between subtelocentric or acrocentric chromosome pairs. These results support that numerous fusion events occurred in the karyotype differentiation within this species.
Few fish species have already shown telomeric DNA sequences located far from the chromosomal ends, which are usually observed at the pericentromeric regions and between centromeres and the real telomeres. 26 There are different kinds of ITS, like heterochromatic ITSs (het-ITSs), that is, extended blocks of telomeric-like DNA mainly in centromeric or pericentromeric chromosomes, considered as unstable regions, where fissions and inversions might occur during karyotype evolution, and could represent possible fission points at which new telomeres can be seeded by the pre-existing telomeric repeats. 27 Short ITSs (s-ITSs) are stretches of limited numbers of telomeric hexamers distributed at internal positions and presumably present in all vertebrate species. The s-ITS insertion events could be explained by telomeric repeats that were inserted by telomerase at the intrachromosomal sites through the repair of double-strand breaks that occurred in the germline during evolution.27,28 Recently, Schmid and Steinlein 29 found a new category of large ITSs in anuran species, named euchromatic-ITS or eu-ITS, which was divided into: (1) restricted eu-ITS, restricted to few euchromatic regions with a random distribution pattern, and (2) dispersed eu-ITS, presented in many euchromatic regions. In this study, FISH using TTAGGGn probes revealed the presence of het-ITSs in R. latirostris species (chromosome 1 with vestiges of ITS and chromosome 3 presented colocated ITS/5S rDNA sites), and in karyomorph B of R. pentamaculata from the Barra Grande river (chromosome 1 with vestiges of ITS).
Among specimens of R. pentamaculata assessed, 2n = 56 chromosomes and karyotype organization were the same for populations from the Barra Grande and Juruba rivers. This diploid number has already been described for other populations of this species in the Upper Paraná river, with some differences in the karyotype formula caused by pericentric inversions24,25,30 and seems to be the most widely distributed. However, the karyomorph B (2n = 54 chromosomes) occurs in sympatry and syntopy with karyomorph A (2n = 56 chromosomes) in Barra Grande river, without any evidence of hybrid formation. The analysis of chromosome markers showed a small variation of 5S rDNA sites and occurrence of ITS traces in a metacentric chromosome pair of karyomorph B, indicating Robertsonian fusion and numeric reduction to 2n = 54 chromosomes. In addition to the numeric reduction, the comparison of chromosome formula and FN indicate other chromosomal rearrangement in the karyomorph differentiation.
Our study presented the first cytogenetic results for R. stellata and R. capitonia, which were captured in sympatry and syntopy in the Uruguay river. A comparison of karyotype of these two species evidenced divergent karyotype organization, consistent with reproductive isolation. R. stellata presented 2n = 54 chromosomes, whereas R. capitonia had 2n = 64 chromosomes. This 2n variation is correlated to the increase of the number of acrocentric chromosome pairs in R. capitonia, an event that may be due to centric fissions. Species of Rineloricaria with high diploid numbers presented most of their st/a chromosomes when compared with species of low diploid number.7,30 R. stellata had 20 m/sm and 34 st/a pairs, whereas R. capitonia presented 6 m/sm and 58 st/a pairs with FN = 74 and 70, respectively, which corroborates the Robertsonian events in the species diversification and the pericentric inversions for alterations of FN.
R. stellata had multiple 18S rDNA sites, which was located in only 1 sm chromosome pair in R. capitonia. On the other hand, all species analyzed presented multiple 5S rDNA sites. The high number of 18S rDNA sites in R. stellata and the high number of 5S rDNA sites may be associated with transpositions of these gene families in the karyotypes or the association of transposable elements causing its dispersion, as it has already been described for fish.31–33 Symonová et al. 33 proposed that in some cases rDNAs use the spread mechanism of transposable elements to increase the number of copies, affect recombination rates, and ease the karyotype differentiation among close genomes. Besides the species of this study, differences in the number and phenotypes of rDNA sites were also detected in R. lanceolata, 8 indicating that distribution and dispersion of these repetitive DNAs may contribute to the genomic diversification within this genus.
In addition to the movement of repetitive DNAs and their power of mutation to generate chromosomal rearrangements,34,35 there is another factor that can be correlated to the numerous Robertsonian rearrangements in Rineloricaria, which is the probable instability of telomeres. Perry et al. 36 proposed that ITS turn chromosomes susceptible to new breaks and occurrence of chromosomal rearrangements, like fissions. Chromosome fusions are assigned to mechanisms of telomere shortening and inactivation, the presence of DNA decondensed regions, and the occurrence of terminal repetitive sequences of chromosomes similar to that found in centromeres.37,38 In R. lima, the mechanisms of telomere instability and fusion of acrocentric pairs, in addition to the fusion of acrocentric pairs carrying 5S rDNA in the terminal region, were used to explain chromosome variation. 7
In this study, we proposed that terminal location in st/a chromosome pairs of repetitive sequences for some species, like rDNAs, provides decondensed regions leading to Robertsonian fusion event. This fact is corroborated by the centromeric location of these rDNAs in m chromosomes in species with low diploid number. In addition to these rearrangements, end-to-end fusions of acrocentric chromosomes support the karyotype organization of species with low diploid number in Rineloricaria. On the other hand, species tending to have an increase of their diploid number would present centric fissions as the main mechanism of karyotype differentiation. Thus, cytogenetic data in the genus Rineloricaria indicate that telomeric instability, inversions, and Robertsonian events supported the mechanisms of karyotypic diversification of species, with the likely occurrence of repetitive DNAs in hotspots for chromosomal rearrangements.
Footnotes
Acknowledgments
This study was financed by CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior), Fundação Araucária (Fundação Araucária de Apoio ao Desenvolvimento Científico e Tecnológico do Estado do Paraná), CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico), and SETI/UGF (Secretaria de Estado da Ciência, Tecnologia e Ensino Superior/Unidade Gestora do Fundo do Paraná).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.