
Abstract
Abstract
The aim of this study was to investigate lithium and 2,4-dichlorophenol (2,4-DCP)-induced circadian rhythm disorder and their genome-wide effects in zebrafish. Zebrafish larvae were exposed to 250 ppm LiCl (n = 40) or 20 ppm 2,4-DCP. RNA was subsequently extracted and determined quantitatively. The mRNA levels of circadian clock-related genes, including clock1a, bmal1b, per2, and per1b, were determined. Microarray datasets were generated and the differentially expressed genes (DEGs) were identified. The mRNA levels of some upregulated and downregulated DEGs were examined by quantitative real-time polymerase chain reaction (RT-PCR). Finally, gene ontology (GO) enrichment analysis was applied to determine the roles of the DEGs. The mRNA expression levels of circadian rhythm-related genes in the daily cycle were significantly affected after incubation of zebrafish with LiCl and 2,4-DCP. Many genes were differentially expressed during the light phase (97 h) and RT-PCR validation tests revealed that the expression patterns of DEGs were in accordance with those obtained by microarray analysis. GO functional enrichment analysis showed that the DEGs in LiCl- and 2,4-DCP-treated groups were associated with signal transduction and development. Collectively, our findings indicate that LiCl and 2,4-DCP could affect signal transduction pathways and immune response, thereby inducing circadian rhythm disorder.
Introduction
E
Chronic administration of lithium modulates cognitive judgment bias, 2 affects renal acid excretion, 3 and is involved in diseases such as bipolar disorder. 4 Lithium has been shown to regulate behavior and mood in mice, 5 and has also been found to be associated with bone regeneration, 6 Ebstein's cardiac anomaly in infants, 7 and renal failure. 8 Moreover, it has been used in the treatment of neuropsychiatric disorders and can also affect circadian rhythms. 9 These studies have contributed to our understanding of the relationship between lithium exposure and psychopathological and physiological disorders.
2,4-DCP is typically used as a key intermediate to synthesize herbicides, pesticides, preservatives, disinfectants, and medicines. 10 It is commonly present in the environment and has been recognized as a priority pollutant in aquatic environments in China 11 and the United States. 12 Short-term 2,4-DCP exposure can induce global DNA hypermethylation, thereby causing major toxic effects in fish. 13 It also disrupts endocrine function in zebrafish 14 and induces endoplasmic reticulum stress-mediated apoptosis in vitro 15 and hepatotoxicity in vivo. 16 However, it has not been ascertained whether 2,4-DCP can affect circadian rhythm.
Circadian rhythm disorder is associated with neuropsychiatric illnesses, including manic depression, 17 bipolar disorder, 18 schizophrenia, 19 and adult attention-deficit hyperactivity disorder, 20 as well as tumor progression, 21 infectious and inflammatory diseases. 22 In addition to the effects of genetic and physiological factors, the circadian rhythms of plants and animals can become disrupted or arrhythmic following exposure to certain environmental pollutants, such as light pollution, 23 metals, 24 pesticides, 25 and other organics. 26
In the present study, an experimental model using the zebrafish, which is a sentinel for aquatic pollution, was used to validate and determine the effects of lithium and 2,4-DCP on living organisms. Circadian clock-related genes, including clock1a and bmal1b, two crucial clock genes that participate in circadian modulation, 27 and the clock genes per1b and per2, which are involved in regulating the physiological effects of sleep disruption, 28 were analyzed to identify circadian rhythm abnormalities caused by lithium and 2,4-DCP. The abnormal gene expressions caused by lithium and 2,4-DCP exposure and the associated biological processes were further analyzed to identify the underlying mechanisms of toxicity in living organisms.
Materials and Methods
Animals and RNA isolation
A total of 80 zebrafish larvae (60 h post hatching) were maintained in 250 ppm lithium chloride (LiCl) (n = 40) or 20 ppm 2,4-DCP (n = 40) in a closed recirculating water system with a 14 h light/10 h dark photoperiod. Simultaneously, control zebrafish larvae (n = 40) were maintained in dechlorinated running tap water in the same light and photoperiod conditions. Total RNA was extracted from larvae at different time points during incubation (72–86 h: light phase; 86–96 h: dark phase) using the methods described by Shi et al. 29 RNA concentration was determined quantitatively using a NanoDrop ND-2000 Spectrophotometer (Thermo Fisher Scientific, Inc.), and RNA integrity was assessed using an Agilent 2100 Bioanalyzer (Agilent Technologies).
mRNA expression assay of clock1a, bmal1b, per2, and per1b
To confirm the effects of LiCl and 2,4-DCP on changes in the expression of the circadian clock system genes, the mRNA levels of circadian clock-related genes, including clock1a, bmal1b, per2, and per1b, were determined by quantitative real-time polymerase chain reaction (RT-PCR). This analysis was conducted using a SYBR Green PCR kit (Toyobo, Tokyo, Japan), in accordance with the manufacturer's recommended protocols, in conjunction with an ABI 7300 Real-time PCR System (Applied Biosystems). The primers used in this study were the same as those used by Amaral et al. 30
Array hybridization
To identify the genome-wide effects of LiCl and 2,4-DCP exposure on zebrafish, RNA samples were labeled and hybridized to the Agilent zebrafish (V3) gene expression microarray 4 × 44K (Design ID 026437) and eluted according to the manufacturer's standard protocols. In brief, total RNA was reverse-transcribed to double-stranded cDNA, and then cyanine-3-CTP-labeled cRNA was synthesized. The labeled cRNAs were fragmented and hybridized to the microarray. The arrays were then washed and subsequently scanned using an Agilent G2505C Scanner (Agilent Technologies).
Microarray data analysis
Raw data from the array images were generated using Feature Extraction software (version 10.7.1.1; Agilent Technologies). Data analysis was performed using GeneSpring (version 11.0; Agilent Technologies). Initially, the raw microarray data were normalized using the quantile normalization method. The probes having a 100% match were collected for further data analysis. The differentially expressed genes (DEGs) were then identified using the criteria of a threshold of fold change (FC) ≥2.0 (log2FC ≥1) and a p-value <0.05.
Differential gene expression assayed by RT-PCR analysis
The mRNA levels of some significantly upregulated and downregulated DEGs obtained from a comparison of the control and LiCl-treated groups, as well as those from a comparison of control and 2,4-DCP-treated groups, were determined by quantitative RT-PCR measurement. The cycle threshold (CT) values obtained from three replicates within each assay were generated and compared with the log2FC values. This enabled us to assess the prediction accuracy of the microarray dataset.
Functional enrichment analysis of DEGs
The gene ontology (GO, www.geneontology.org) consortium is a project designed to facilitate the biologically meaningful annotation of genes. 31 GO enrichment analysis was applied in the present study to determine the functional characteristics of the identified DEGs.
Results
The effects of LiCl and 2,4-DCP on the mRNA levels of circadian rhythm-related genes
The mRNA expression levels of circadian rhythm-related genes, particularly those of clock1a and bmal1b, were significantly affected during the daily cycle of zebrafish incubated with LiCl (Fig. 1A, C). In addition, significant changes were found in the mRNA levels of per2 and per1b during the light cycle, but not during the dark cycle (Fig. 1E, G). This difference may be due to the low expression levels of per2 and per1b in darkness. Similar results were obtained for zebrafish incubated with 2,4-DCP (Fig. 1B, D, F, H).

Quantitative analysis of the expression levels of clock1a, bmal1b, per2, and per1b in control, LiCl-
DEGs obtained from the microarray datasets
To explore the molecular mechanism underlying the LiCl- and 2,4-DCP-induced toxic effects in zebrafish models, the gene expression profiles of zebrafish after 97 h of incubation were generated and analyzed.
Compared with the control group, a total 1105 downregulated DEGs and 1041 upregulated DEGs were obtained in the LiCl-treated group, whereas 1211 downregulated DEGs and 1088 upregulated DEGs were screened from the 2,4-DCP-treated group.
RT-PCR validation of DEGs
To validate the accuracy of mRNA expression in the microarray datasets, some top DEGs with large log2FC values were analyzed using RT-PCR. We determined the mRNA levels of DEGs, including ppp2r5cb (log2FC = −6.86), atp5c1 (log2FC = −5.40), and ccnt2a (log2FC = −8.54), and plxna4 (log2FC = 7.73) and pank2 (log2FC = 5.95), obtained from the comparison between the control and LiCl-treated groups (Fig. 2A). Similarly, for the 2,4-DCP-treated group, we analyzed the upregulated genes lgals2b (log2FC = 5.64), rpl17 (log2FC = 5.70), and ireb2 (log2FC = 6.94), and the downregulated genes cyb561a3b (log2FC = −5.77) and ccnt2a (log2FC = −7.69) (Fig. 2B). The primers used are shown in Table 1. The RT-PCR results showed that although the CT values were not completely consistent with the log2FC values, all the gene expression patterns were in accordance with those obtained by microarray analysis. Thus, the datasets were considered suitable for the following analyses.
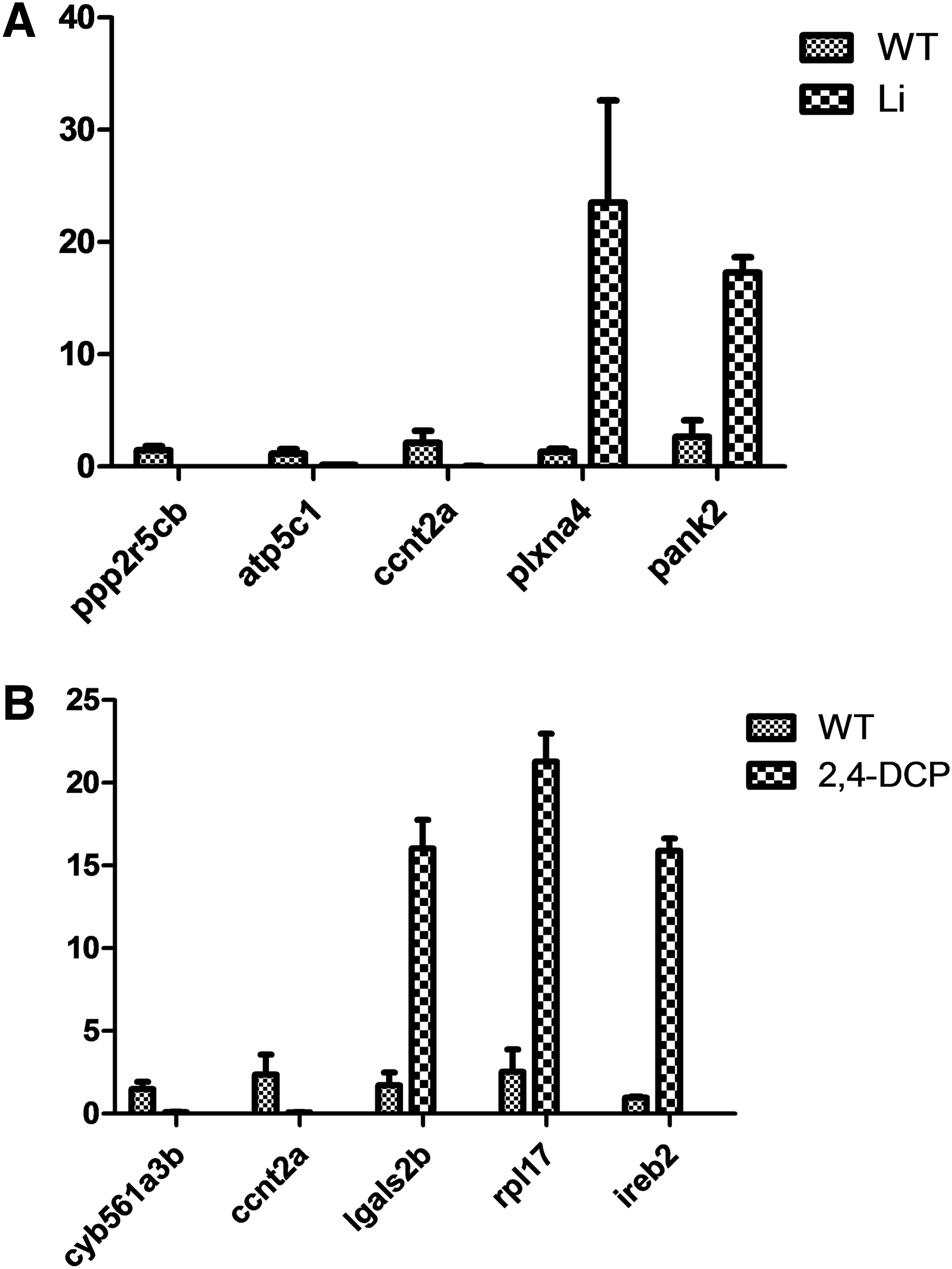
Quantitative analysis of the expression levels of the differentially expressed genes (DEGs) ppp2r5cb, atp5c1, and ccnt2a (downregulated), and plxna4 and pank2 (upregulated) in the LiCl-treated group
2,4-DCP, 2,4-dichlorophenol; WT, wild-type.
GO functional terms enriched by DEGs
The GO-BP (biological process) terms enriched by DEGs in the two treatment comparisons are shown in Figure 3. The upregulated DEGs in the LiCl-treated group (Fig. 3A) were mainly associated with fin regeneration, signal transduction, immune response, and frontonasal suture morphogenesis, whereas the downregulated DEGs (Fig. 3B) were primarily related to biological process, signal transduction, skeletal muscle fiber development, and embryonic skeletal system development. In the 2,4-DCP-treated group, upregulated DEGs (Fig. 3C) were mainly enriched in intracellular signal transduction, biological process, and regulation of synaptic vesicle lumen acidification, whereas the downregulated DEGs (Fig. 3D) were mainly associated with biological process, signal transduction, motor neuron migration, and face development.
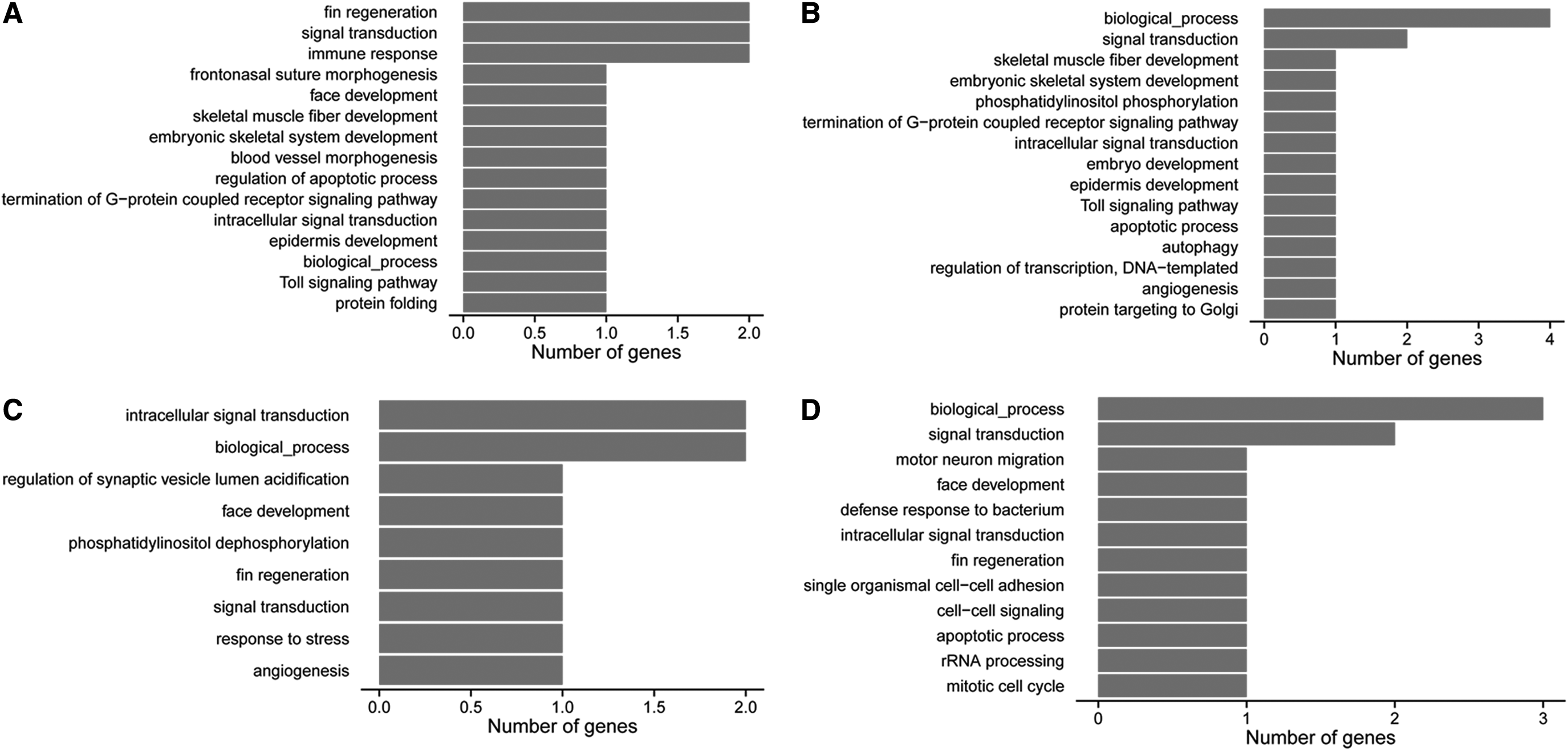
Gene ontology (GO) enrichment analysis of DEGs in the LiCl-treated group (upregulated DEGs
Discussion
Environmental pollutants, including lithium and 2,4-DCP, can have potentially adverse effects on living organisms. In this study, the abnormal expression levels of circadian rhythm-related genes, including per1a, per1b, bmal1b, and clock1a, were identified after LiCl and 2,4-DCP treatment. The accuracy of the gene expression profile was subsequently validated by analysis of DEGs. Functional enrichment analysis indicated that the DEGs identified in LiCl- and 2,4-DCP-treated group were mainly associated with signal transduction, morphogenesis and development, and immune response. These results provide important insights into circadian rhythm abnormalities and will facilitate elucidation of the mechanisms underlying the toxicity induced by lithium and 2,4-DCP.
In our study, mRNA levels of per1a, per1b, bmal1b, and clock1a in both LiCl- and 2,4-DCP-treated groups differed from those in the control group, notably during the light phase. Moreover, changes in the mRNA expression of per1a and per1b were more pronounced than those in the mRNA levels of bmal and clock1a. Previous studies have indicated that the expression levels of per1a and per1b are associated with acute stress response, whereas no concomitant responses were identified in the mRNA levels of bmal1b and clock1a, the transcripts of which showed significant changes only upon extended exposure. 32 This indicates that the mRNA expression levels of circadian rhythm-related genes were under the influence of both LiCl and 2,4-DCP.
The later validation of DEG expression by RT-PCR revealed the accuracy of the gene expression profiles. Subsequent functional analysis showed that signal transduction was enriched by DEGs in both LiCl- and 2,4-DCP-treated groups. Signal transduction pathways are crucial for physiological effects and circadian rhythms via multiple factors, including neuropeptide Y, 33 melatonin receptors, 34 and intermittent hypoxia/reoxygenation. 35 Signal transduction also controls genome-wide transcription in the circadian clockworks that underlie circadian oscillations. 36 Therefore, LiCl and 2,4-DCP could influence signal transduction pathways to induce both circadian rhythm and physiological dysfunction.
In this study, GO-BP terms such as fin regeneration, skeletal muscle fiber development, and face development were enriched by DEGs in both LiCl- and 2,4-DCP-treated groups. Previous studies have indicated that the environmental pollutants LiCl and 2,4-DCP can adversely affect tissue development and regeneration in larval zebrafish.37,38 Lithium has been found to be associated with bone regeneration, 6 axonal regeneration, 39 and caudal fin regeneration, 40 and 2,4-DCP14,15 has been observed to have a similar toxic effect.
In addition, immune responses could be activated after LiCl treatment. Previous studies have indicated that lithium, which is an anti-inflammatory agent and diminishes aggression, could contribute to the suicide-linked traits. 41 Lithium acts on the immune system that plays both inhibitory and stimulatory roles in disorders of immunity and inflammation. 42 In addition, the inflammatory immune response has been shown to be mediated by the circadian clock.43,44 The bidirectional relationship between circadian rhythms and the immune system indicates that immune response is sensitive to alterations in circadian rhythms. 45 However, the causal relationship between immune response and the circadian system remains to be explored.
As a result, lithium and 2,4-DCP can induce circadian rhythm abnormality. The gene expression profiles obtained in the present study had good accuracy and precision. Collectively, signal transduction pathways could be affected by exposure to LiCl and 2,4-DCP, accompanied by damage to tissue development and regeneration. Moreover, LiCl-induced toxic effects are closely related to immune responses. However, there are certain limitations to our study. In future studies, we will need to further identify the crucial genes and biological processes involved in the induced responses and validate the experimental data.
Footnotes
Acknowledgment
This work was supported by the National (Youth) Natural Science Foundation of China (grant no. 21277115, 41302272, 41501339), the China Postdoctoral Science Foundation (no. 2013M540461), and the Youth Natural Science Foundation of Jiangsu Province (no. BK20140330).
Disclosure Statement
No competing financial interests exist.