
Abstract
Abstract
Antibodies raised against mammalian proteins may exhibit cross-reactivity with zebrafish proteins, making these antibodies useful for fish studies. However, zebrafish may express multiple paralogues of similar sequence and size, making them difficult to distinguish by traditional Western blot analysis. To identify the zebrafish proteins that are recognized by an antimammalian antibody, we developed a system to screen putative epitopes by cloning the sequences between the yeast SUMO protein and a C-terminal 6xHis tag. The recombinant fusion protein was expressed in Escherichia coli and analyzed by Western blot to conclusively identify epitopes that exhibit cross-reactivity with the antibodies of interest. This approach can be used to determine the species cross-reactivity and epitope specificity of a wide variety of peptide antigen-derived antibodies.
H
We have used the recombinant SUMO solubility tag expression system to experimentally determine (1) whether a commercially available anti-human antibody recognizes zebrafish homologues of a human protein and (2) whether the antibody discriminates among zebrafish paralogues. To accomplish this, we first modified the pET28-SUMO vector, by deleting the N-terminal 10xHis tag, creating the vector pET28-SUMOΔNHis (Supplementary Fig. S1). This modification makes only the C-terminal 6xHis tag available for purification and detection. The putative zebrafish epitope was then inserted between the N-terminal SUMO tag and the C-terminal 6xHis tag (Fig. 1A). 4 Fusion proteins were recombinantly expressed in and purified from Escherichia coli (Supplementary Materials and Methods section and Supplementary Figs. S1 and S2) before analysis by western blotting with both an anti-His probe and the antibody of interest to test cross-reactivity.
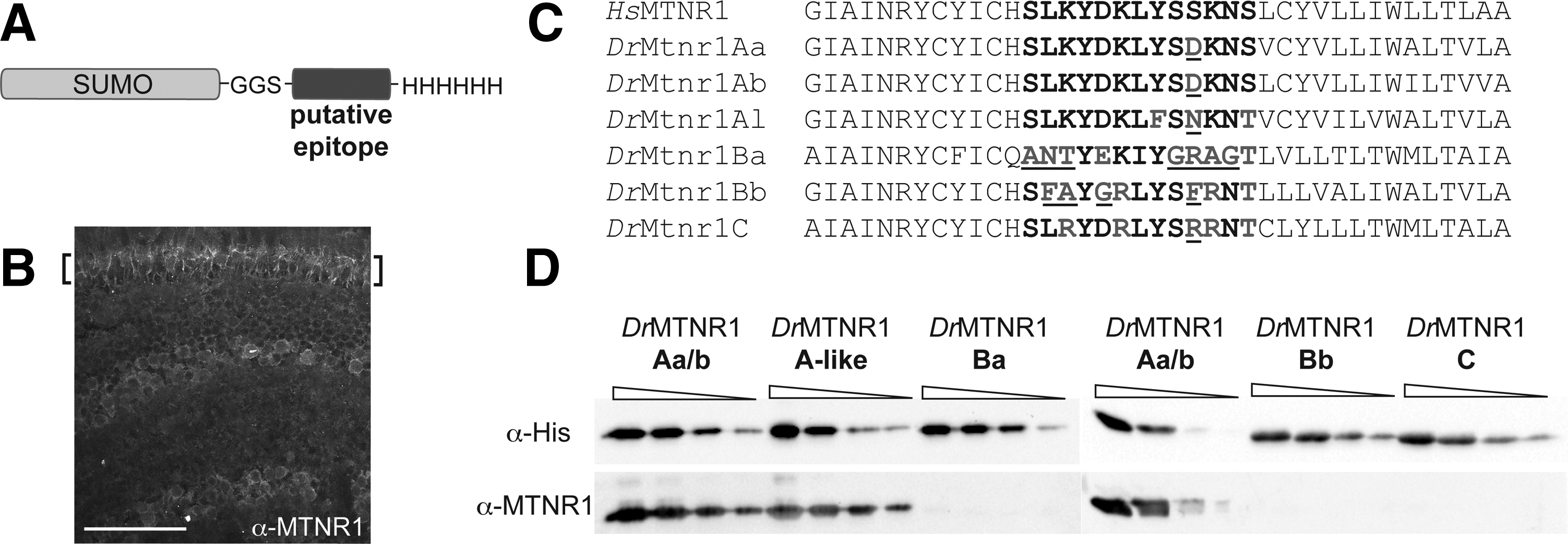
Identification of immunoreactive MTNR1 peptides in zebrafish retina.
The C-terminal location of the 6xHis tag affords multiple advantages beyond facilitating rapid purification. A control western blot that probes the 6xHis tag serves as a direct loading control for the test antibody western blot. In addition, it provides a mechanism for monitoring the integrity of the putative epitope peptide. If the recombinant protein is not able to bind to either the affinity resin or the anti-His antibody, then there is reason to suspect that both the C-terminal 6xHis and the putative epitope have been degraded during cell lysis and/or purification. In that case, a new protein preparation should be made before performing the test antibody western blot.
We have demonstrated the facility of this approach using a commercial antibody (ab87639; Abcam) raised against a 13-residue peptide within the human melatonin receptor 1 protein (HsMTNR1). Immunohistochemistry experiments clearly show antibody reactivity with a protein in zebrafish retinas (Fig. 1B) 5 ; however, the zebrafish genome contains six different mtnr1 paralogues (Fig. 1C). Of the proteins encoded by these genes, two (DrMtnr1Aa and DrMtnr1Ab) share the exact same putative epitope sequence with one another. We have recombinantly expressed and purified SUMO fusion peptides of the five unique putative epitopes from the zebrafish Mtnr1 proteins (DrMtnr1Aa/b, DrMtnr1A-like, DrMtnr1Ba, DrMtnr1Bb, and DrMtnr1C). Titrations of these proteins were analyzed by SDS-PAGE and two western blots (anti-His and anti-HsMTNR1) (Fig. 1D and Supplementary Materials and Methods section and Supplementary Fig. S3). These data clearly show that the anti-HsMTNR1 antibody reacts with both the DrMtnr1Aa/b epitope and the DrMtnr1A-like epitope. The antibody does not recognize the putative epitope within the DrMtnr1Ba, DrMtnr1Bb, or DrMtnr1C paralogues. It is notable that this approach allows us to probe how precisely the antibody recognizes the chemical features of the epitope. DrMtnr1C contains only one nonconservative chemical difference between the two sequences (Ser10→Arg10 within the epitope), whereas the other four differences with the antigenic sequence are very chemically conservative differences (Lys→Arg and Ser→Thr). Even so, the antibody does not recognize the DrMtnr1C sequence at all, demonstrating that the size, shape, and charge of these amino acid side chains matter greatly. This approach can, therefore, also be used to probe the features of a putative epitope that are necessary for antibody binding.
Footnotes
Acknowledgments
We thank Benjamin Euhus, Katie Shaw, and Leticia Gonzalez for assistance with protein expression and purification; the Texas State University Student Undergraduate Research Fund (to M.A.V. and B.J.R.); and Chris Lima (Rockefeller University) and Cornell University for the pET28-SUMO expression plasmid. Confocal microscopy was performed on an instrument supported by NSF grant DBI-0821252 (to J. Koke and D.M.G.).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.