
Abstract
Abstract
Serotonin (5-hydroxytryptamine [5-HT]) is a bioactive monoamine that acts as a neurotransmitter in the central and peripheral nervous system of animals. Teleost fish species have serotonergic neurons in the raphe nuclei of the brainstem; however, the role of 5-HT in the raphe neurons in teleost fish remains largely unknown. Here, we established a medaka (Oryzias latipes) strain with targeted disruption of tryptophan hydroxylase 2 (tph2) gene that is involved in the 5-HT synthesis in the raphe nuclei. Immunohistochemistry and mass spectrometry analysis revealed that the homozygous mutants (tph2Δ13/Δ13) lacked the ability to synthesize 5-HT in the raphe neurons. To investigate the effects of 5-HT deficiency in adult behaviors, the mutant fish were subjected to five behavioral paradigms (diving, open-field, light–dark transition, mirror-biting, and two-fish social interaction). The homozygous mutation caused a longer duration of freezing response in all examined paradigms and reduced the number of entries to the top area in the diving test. In addition, the mutants exhibited a decreased number of mirror-biting in the males and an increased contact time in direct social interaction between the females. These results indicate that this tph2-knockout medaka serves as a good model to analyze the effects of 5-HT deficiency in the raphe neurons.
Introduction
S
Teleost fishes also have serotonergic neurons in both their central and peripheral nervous systems. 4 Major sites for serotonergic neurons in the teleost brains are mainly divided into two areas: the raphe nuclei in the brain stem and posterior tuberculum/hypothalamic populations.5–9 As distinct from mammals, there are at least three tph genes (tph1a, tph1b, and tph2) in teleost fishes. 4 Previous studies in the adult brains of zebrafish (Danio rerio) and medaka (Oryzias latipes) reported that the tph2 gene, an orthologue of the Tph2 in mammals, is expressed by serotonergic neurons of the raphe nuclei, whereas the tph1a is expressed by those of the tuberculum and hypothalamic areas.10,11 In zebrafish, serotonergic neurons in the raphe nuclei send axonal projections to a number of brain regions, including telencephalon, diencephalon, and mesencephalon, 12 which indicates that those neurons would be involved in a wide range of behaviors in teleost fishes, similar to mammals. Recent studies in zebrafish demonstrated that genetic ablation or optogenetic controls of tph2-expressing neurons affected arousal or light–dark preference, respectively.13,14 However, there are no reports on fish whose tph2 gene is genetically disrupted; therefore, the function of tph2 protein in the raphe serotonergic neurons and the behavioral regulation by 5-HT that can be synthesized by tph2 in the neurons has remained largely unknown in teleost species.
Here, to investigate the function of tph2 and its metabolite 5-HT in the raphe neurons in teleost fish, we generated a tph2-knockout medaka strain by a genome editing technology. The 5-HT synthesis activity in brains of the mutants was assessed by immunohistochemistry and mass spectrometry analysis. Subsequently, we evaluated the behavioral differences between wild-type and tph2 homozygous mutant fish. Among the wide range of behaviors that can be controlled by the raphe serotonergic neurons, we investigated the effects on anxiety-related and social behaviors that have been well characterized in medaka and zebrafish by several behavioral paradigms, such as diving, open-field, light–dark transition, mirror-biting, and two-fish social interaction.15–20
Methods
Fish and housing conditions
The d-rR strain (closed colony) 21 of medaka was used in this study. Embryos were raised in embryo culture medium (0.1% NaCl, 0.003% KCl, 0.004% CaCl2 · 2H2O, and MgSO4 · 7H2O) on a 14/10 h light/dark cycle at 26°C. Hatched fish were raised in groups of 8–10 fish per 2 L tank, with recirculating, filtered water at 27°C. The fish tanks were illuminated by a white fluorescent tube on a 14/10 h light/dark cycle (07:00–21:00 light). The fish were fed newly hatched Artemia sp. nauplii and commercial powdered foods (for larval fish: Sweetfish Super Gold #0; Oriental Yeast, Tokyo, Japan; for juvenile and adult fish: Otohime B2; Marubeni Nisshin Feed, Tokyo, Japan), once or twice a day. The sex of each fish was determined by the shape of the anal fin.
Targeted gene disruption using transcription activator-like effector nucleases
Potential transcription activator-like (TAL) effector nuclease (TALEN) target sites in the sixth exon of tph2 gene (Ensembl Transcript ENSORLT00000000279.1) were searched by using the TALEN Targeter program (https://tale-nt.cac.cornell.edu/node/add/talen). 22 TAL repeats were assembled by a modified Golden Gate assembly method, as previously described. 23 Capped RNA encoding each TALEN was transcribed by using the mMessage mMachine SP6 kit (Life Technologies, Carlsbad, CA), followed by purification with the RNeasy mini kit (Qiagen, Hilden, Germany). A pair of TALENs (150 ng/μL) was injected into the fertilized eggs of the d-rR strain by a microinjection method. 24
The tph2 mutants were genotyped by the heteroduplex mobility assay with an automated microchip electroporation system 25 by using PCR primers 5′-TGACCCATACTCCCCACAGT-3′ and 5′-GCCACAGTGTTTGGTGAGAA-3′. Sequences of the identified mutant alleles were determined by direct sequencing of the PCR product amplified by using primers 5′-AGGGTTTCAAGGACGAGGTT-3′ and 5′-TCTCGAGGAGACAGGTAGCC-3′.
Immunohistochemistry
For whole-mount staining of embryos, embryos whose chorion was removed by using the medaka hatching enzyme (National BioResource Project medaka, Okazaki, Japan) were fixed overnight with 4% paraformaldehyde in phosphate-buffered saline (PBS) at 4°C. Fixed embryos were rinsed in PBS containing 0.05% Tween-20 (PBST) and then dehydrated in a graded methanol series (25%, 50%, 75%, and 100%). The embryos were rehydrated in a reverse methanol series, rinsed in water, permeabilized by cold acetone at −20°C for 20 min, rinsed in water, and rinsed in PBST. The embryos were then blocked at 4°C overnight in PBS with 0.3% Triton X-100, 1% dimethyl sulfoxide, 2% normal goat serum, and 0.1% bovine serum albumin. The blocked embryos were incubated in a primary antiserum containing a serotonin rabbit antibody (ImmunoStar, Hudson, WI; #20080, 1:500) overnight at 4°C, washed with PBST, and finally incubated in a secondary antiserum containing a donkey anti-rabbit immunoglobulin G (IgG) secondary antibody, Alexa Fluor® 488 conjugate (Life Technologies; R37118, 1:500) at 4°C overnight. The embryos were washed with PBST and cleared in a glycerol series (30%, 70%, and 100%). The fluorescent images were captured by using a fluorescence stereomicroscope MZFLIII with a GFP2 filter set (Leica Microsystems, Wetzlar, Germany) with a digital CCD camera VB-7010 (KEYENCE, Osaka, Japan).
For adult brains, fish were terminally anesthetized with MS-222 (Sigma, St. Louis, MO). The dissected heads were fixed with Bouin's solution for a day at room temperature, dehydrated with a series of the Tissue Dehydration Solution A (Wako Pure Chemical Industries, Osaka, Japan) (50%, 70%, and 100%), and embedded in paraffin. Paraffin-embedded heads were sectioned at 10 μm. Paraffin sections were deparaffinized with xylene, dehydrated with the Tissue Dehydration Solution A, and finally rehydrated with PBS. The sections were blocked in PBS with 0.25% Triton X-100 containing 2% normal goat serum for an hour at room temperature; incubated in a primary antiserum containing a serotonin rabbit antibody (ImmunoStar; #20080, 1:2000) for an hour at room temperature; washed with PBS with 0.25% Triton X-100; and finally incubated in a secondary antiserum containing an anti-rabbit IgG, alkaline phosphatase-linked antibody (Cell Signaling Technology, Danvers, MA; 1:1000) for 30 min at room temperature. Signals were developed by using nitroblue tetrazolium and 5-bromo-4-chloro-3-indolyl phosphate in buffer containing 100 mM NaCl, 100 mM Tris-HCl (pH 9.5), 50 mM MgCl2, and 0.1% Tween-20. The sections were photographed by using a light transmission microscope (BX51; Olympus, Tokyo, Japan) with a digital CCD camera (DP72; Olympus).
Neurochemical assessment by liquid chromatography with tandem mass spectrometry
Adult fish were terminally anesthetized with MS-222 (Sigma), and then each brain was dissected and immediately stored at −80°C until the extraction step. The brain was homogenized in 100 μL of ice-cold extraction buffer containing 4% HClO4, 0.2% ethylenediaminetetraaceticacid, and 0.05% NaHSO3, followed by centrifugation at 20,000 g, for 20 min. The pellet was solubilized in 100 μL of 0.5 N NaOH at 60°C for 20 min, and the protein content was quantified by using Micro BCA Protein Assay Kit (Thermo Scientific, Rockford, IL). The supernatant was subjected to the neurochemical quantification. Liquid chromatography with tandem mass spectrometry (LC-MS/MS) analysis was performed by using a high-performance liquid chromatograph Prominence system coupled with a triple quadrupole mass spectrometer LCMS-8040 with an ESI interface (Shimadzu, Kyoto, Japan). The neurochemicals were separated on a reverse-phase porous ODS column Unison UK-C18 (particle size: 3 μm, internal diameter: 2.0 mm, length: 150 mm; Imtakt, Kyoto, Japan) at 40°C with a binary gradient elution by using 0.1% formic acid (solvent A) and 80% acetonitrile (solvent B) at a flow rate of 0.2 mL/min. The gradient profile was as follows: 0–15 min, 0%–25% B; 15–20 min, 25%–100% B; 20–25 min, 100% B; 25–25.01 min, 100%–0% B; and 25.01–40 min, 0% B. 5-HT, tryptophan, 5-HTP, and dopamine were detected in each brain homogenate by the multi-reaction monitoring, whose conditions were optimized by using the standard chemicals obtained from Nacalai Tesque (Kyoto, Japan). Each neurochemical was quantified by the total ion current of three fragment ions as follows: 5-HT: m/z 177.20 > 115.10, 117.10, 132.10; 5-HTP: m/z 221.20 > 162.00, 134.10, 77.05; tryptophan: m/z 205.25 > 146.05, 118.10, 91.10; dopamine: m/z 154.20 > 91.05, 65.05, 119.05. The amount of each neurochemical was calibrated by using a standard curve plotted for four different amounts of the standard chemical (0.5, 0.1, 0.05, 0.01 ng).
Micro-computed tomography imaging
Anesthetized fish were fixed with 10% formalin neutral buffer solution (Wako Pure Chemical Industries) overnight at room temperature and were then washed with PBS. X-ray images were captured by using an in vivo micro X-ray computed tomography (CT) system (R_mCT2; Rigaku, Tokyo, Japan). Scanned images were reconstructed by using OsiriX (Pixmeo SARL; Bernex, Switzerland).
Behavioral testing
Fish
Both wild-type (tph2+/+) and homozygous mutant (tph2Δ13/Δ13) fish were subjected to behavioral testing at 8 weeks post-hatching (wph). The body length of each fish was measured with a ruler; images of the fish were captured by using a digital camera (CX2; Ricoh, Tokyo, Japan); and the number of pixels was quantified by using ImageJ software (National Institutes of Health, Bethesda, MD; http://imagej.nih.gov/ij/).
Acquisition of behavioral data
Behavioral data were acquired and analyzed by a custom-made software previously described. 15 Briefly, fish images captured were every 100 ms by using a USB webcam. The images were subtracted from the background and were binarized. The gravity points of the binarized fish were used for fish positions. All behavioral parameters were calculated by the time course of the fish position.
Testing procedures
The wild-type and the mutant fish were maintained as separate groups of 8–10 fish during the testing period. Each fish was subjected to one of five behavioral paradigms (diving, open-field, light–dark transition, mirror-biting, and social interaction) in a day. All the tests were performed from 10:00 to 18:00. To reduce the effects of the diurnal rhythm of behavioral activity, the wild-type and the mutant groups were subjected to the behavioral test, one by one.
Diving test
The diving test is a well-established paradigm that quantifies the diving response to assess stress, fear, and anxiety-related behaviors in zebrafish. 16 Fish were individually introduced into a test tank (200 mm length × 50 mm width × 150 mm height) that was filled with water to a height of 120 mm, and the behavior of each fish was recorded for 10 min from the side. To measure behaviors in the top and bottom area of the tank, the captured images were vertically divided into five areas (24 mm height each) and then the tracking data were separately analyzed in each area.
Open-field test
The open-field test is a paradigm that assesses the locomotor activity and anxiety level in zebrafish. 19 Fish were individually introduced into a test tank (200 mm length × 200 mm width × 75 mm height) that was filled with water to a height of 20 mm, and its trajectories was recorded for 15 min from above. The captured images were virtually divided into center (120 mm × 120 mm, 60% of length and width) and peripheral (the area within 80 mm from the walls) areas.
Light–dark transition test
The light–dark transition test is a behavioral paradigm that is used to assess anxiety in both rodents and teleost fishes, which is based on the conflict between the innate aversion to a brightly illuminated environment and spontaneous exploratory behavior.26–28 A test tank (200 mm length × 50 mm width × 100 mm height) that was filled with water to a height of 30 mm was divided equally into two compartments (100 mm length each). The left half (dark side) consisted of black walls and bottom and was shielded from light with an opaque lid, whereas the right half (light side) consisted of white walls and bottom and was illuminated by an LED panel. Each fish was introduced into the light compartment, and their behaviors were recorded for 10 min from the side.
Mirror-biting test
The mirror-biting test is a well-established model of social behavior, including socially induced anxiety, in medaka. 20 The test tank (200 mm length × 90 mm width × 70 mm height) with a mirror (90 mm width × 70 mm height) on the left wall was filled with water to a height of 20 mm. We divided the captured images into 20 virtual areas (10 mm length × 90 mm width each) and defined stay in an area closest to the mirror as the “mirror-biting” behavior. Fish were individually introduced into the center of the tank, and their trajectories were recorded for 10 min from above.
Social interaction test
The two-fish social interaction test is a behavioral paradigm that is used to evaluate social interactions between two fish, such as contacts and distance between the fish. This paradigm was conceptually inspired by a reciprocal social interaction test in mice. 29 Since a previous study in medaka demonstrated that the distance between two individuals by the coordinated behavior is based on a conspecific social interaction, 30 this paradigm can reflect the changes in sociability of medaka. The test apparatus was the same as the open-field test. A pair of two congeneric fish (male-to-male or female-to-female) was introduced into the tank, and their trajectories were recorded for 10 min from above. Each captured frame at which two individuals were detected as “one” fish object (with “zero” distance) was defined as the “contact.”
Statistical analysis
All data in the graphs are presented as mean ± standard error of the mean. Statistical analyses were performed by using GraphPad Prism 6 for Windows (GraphPad Software, La Jolla, CA) or R version 3.3.1 (https://r-project.org/). Figures were drawn by using Adobe Illustrator CS6 (Adobe Systems, San Jose, CA). Differences between treatments over time were analyzed by using two-way repeated measures (RM) analysis of variance (ANOVA) followed by Sidak's multiple-comparison test. Differences between treatments were analyzed by using Welch's t-test, except latency outcomes, which were analyzed by using survival curves and the log-rank (Mantel-Cox) test.
Results
Targeted disruption of the tph2 gene using TALENs
By searching a medaka genome database, we found at least three genes encoding tph, tph1a (Ensembl gene number ENSORLG00000005962), tph1b (ENSORLG00000015541), and tph2 (ENSORLG00000000223). A previous study showed that the tph2 gene is an orthologue of the human TPH2 and is expressed in the raphe nuclei of the brain stem, dorsal posterior nucleus in the thalamus, and the anterior part of the medulla oblongata. 11 Therefore, for an understanding of the raphe serotonergic controls of teleost fish behaviors, the tph2 gene was selected as a target for a loss-of-function analysis. We designed a pair of TALENs that can recognize and cleave a target site on the sixth exon of the tph2 gene (Supplementary Fig. S1A; Supplementary Data are available online at www.liebertpub.com/zeb) and injected capped RNAs encoding these TALENs into the fertilized eggs. Each injected fish was mated with a wild-type fish, and then, by genotyping of the F1 generation, the founder fish (G0) harboring a 13 bp deletion (tph2Δ13) on the target exon were identified (Supplementary Fig. S1B). This deletion causes a frame shift and results in additional five residues of an altered sequence followed by a premature stop codon (Supplementary Fig. S1B), which would be expected to truncate the tph within its catalytic domain (Biopterin_H, NCBI accession number pfam00351) (Supplementary Fig. S1C).
Spinal deformity in tph2Δ13/Δ13 adult fish
By intercrossing between F1 fish that are heterozygous carriers for the deletion (tph2+/Δ13), an F2 family containing homozygous mutant fish (tph2Δ13/Δ13) was obtained. The homozygous mutant fish were viable and fertile (data not shown); however, they began to display spinal deformity at the adult stage (around 3 months post-hatching). A radiograph acquired by the X-ray CT system showed that scoliosis occurred in 4 month-old homozygous mutants (Fig. 1C, D) but not in the wild-type (Fig. 1A, B).
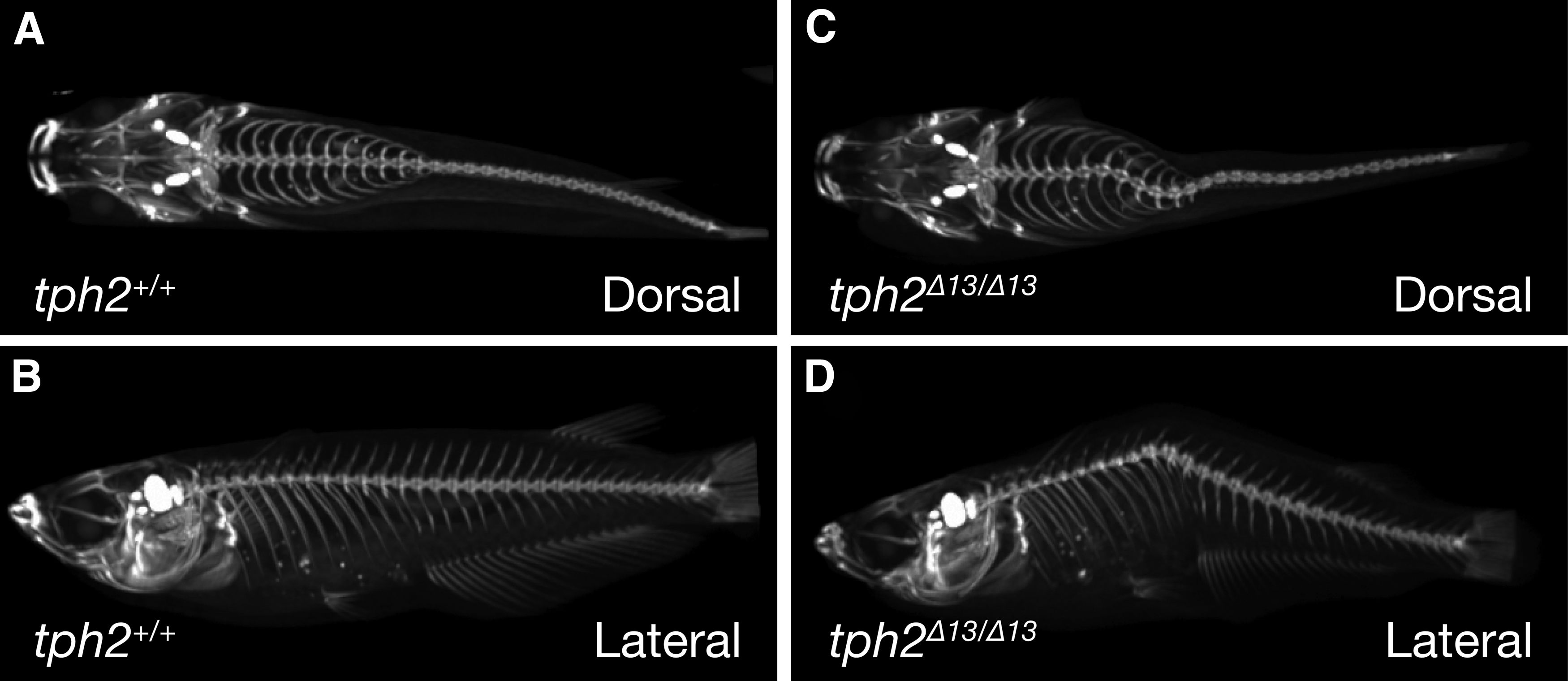
Reconstructed three-dimensional radiography of the skeletal structures captured by X-ray micro-computed tomography. Radiography of tph2+/+
Deficiency of 5-HT synthesis in the raphe nuclei of tph2Δ13/Δ13 fish
To evaluate the enzymatic activity of the truncated tph2 protein, 5-HT in the serotonergic neurons of the brain was detected in both tph2+/+ and tph2Δ13/Δ13 fish by immunohistochemistry using an anti-5-HT antibody. Whole-mount immunostaining of 4-day-post-fertilization embryos showed that the tph2Δ13/Δ13 embryos lost a 5-HT-positive population of cells in the raphe nuclei of the brain stem but not in the pineal gland (Fig. 2A, B). For further analysis of 5-HT distribution in the brain, the coronal sections of the adult brains were immunohistochemically stained. The tph2+/+ brain exhibited the 5-HT-positive signals in the neuronal cells of the raphe nuclei (Fig. 2C), whereas the tph2Δ13/Δ13 brain did not (Fig. 2D). In addition, a number of 5-HT-positive axons were found in the telencephalon of the tph2+/+ brain (Fig. 2E), but not of the tph2Δ13/Δ13 brain (Fig. 2F). On the other hand, there were no differences in the 5-HT-positive signals in the pineal gland (Fig. 2G, H), the pituitary gland (Fig. 2I, J), and the hypothalamus, such as in the posterior parvocellular preoptic nucleus (Fig. 2K, L) or in the posterior recess nucleus (Fig. 2M, N), where other tph genes (tph1a and/or tph1b) are expressed.4,11

Immunohistochemical staining of 5-HT by using anti-serotonin antibody.
Subsequently, for the quantitative assessment of the tph2 enzymatic activity, two metabolites (5-HTP and 5-HT) and a substrate (tryptophan) of the tph were quantified in each wild-type or mutant brain by using the LC-MS/MS with a multi-reaction monitoring method. The tph2Δ13/Δ13 fish showed significant reductions of the metabolites 5-HT (males: t9.479 = 7.855, p < 0.0001; females: t7.845 = 5.530, p = 0.0006; Fig. 3A) and 5-HTP (males: t9.933 = 2.872, p = 0.0167; females: t9.979 = 3.639, p = 0.0046; Fig. 3B). On the other hand, the amount of tryptophan was significantly increased in the tph2Δ13/Δ13 brain (males: t9.969 = 4.080, p = 0.0022; females: t9.737 = 2.292, p = 0.0455; Fig. 3C). The dopamine levels in the brain were not affected by the tph2 mutation (males: t8.552 = 2.002, p = 0.0781; females: t9.864 = 0.3051, p = 0.7666; Fig. 3D). These results indicate that the enzymatic activity is lost in the truncated tph2 protein translated from the tph2Δ13 allele.

Quantification of neurochemicals by liquid chromatography with tandem mass spectrometry in the whole brain of tph2+/+ and tph2Δ13/Δ13 fish. 5-HT
Behavioral phenotyping of the tph2Δ13/Δ13 fish
For behavioral phenotyping, tph2+/+ or tph2Δ13/Δ13 fish were obtained by matings between F2 tph2+/+ and tph2Δ13/Δ13 fish, respectively, and the hatched larvae of the two genotypes were separately reared under the same conditions. The fish at 8 wph were subjected to the following behavioral assessment because they were younger than the starting age for the spinal deformity. There were no significant differences between genotypes in the body length of both males (tph2+/+: 21.59 ± 0.2060 mm, n = 36; tph2Δ13/Δ13: 21.72 ± 0.3175 mm, n = 31; t52.59 = 0.3294, p = 0.7432) and females (tph2+/+: 21.83 ± 0.2124 mm, n = 30; tph2Δ13/Δ13: 21.81 ± 0.3408 mm, n = 27; t44.15 = 0.06154, p = 0.9512).
We first assessed the diving response as an indicator for stress, fear, and anxiety-related behaviors in teleost species. The tph2Δ13/Δ13 fish showed a decrease in the vertical swimming activity in both males (Fig. 4A, C) and females (Fig. 4B, C). The duration of freezing behavior was significantly increased by the mutation in both males (Fig. 4D, F) and females (Fig. 4E, F). Time spent in the bottom area was significantly increased in the mutant females (Fig. 4H, I) but not in the mutant males (Fig. 4G, I). Number of entries into the top area was significantly decreased by the mutation in both males (Fig. 4J, L) and females (Fig. 4K, L). The latency to the first entry into the top area was not affected by the mutation (male: χ2 = 0.06380, p = 0.8006, Fig. 4M; female: χ2 = 0.8050, p = 0.3696, Fig. 4N). Further, there were no differences in the maximum swimming velocity in the 10 min test between the genotypes (male: t32.238 = −0.07432, p = 0.9412; female: t19.047 = 1.8192, p = 0.07801; Fig. 4O). The statistical results of Welch's t-test and two-way RM ANOVA are listed in Supplementary Table S1.
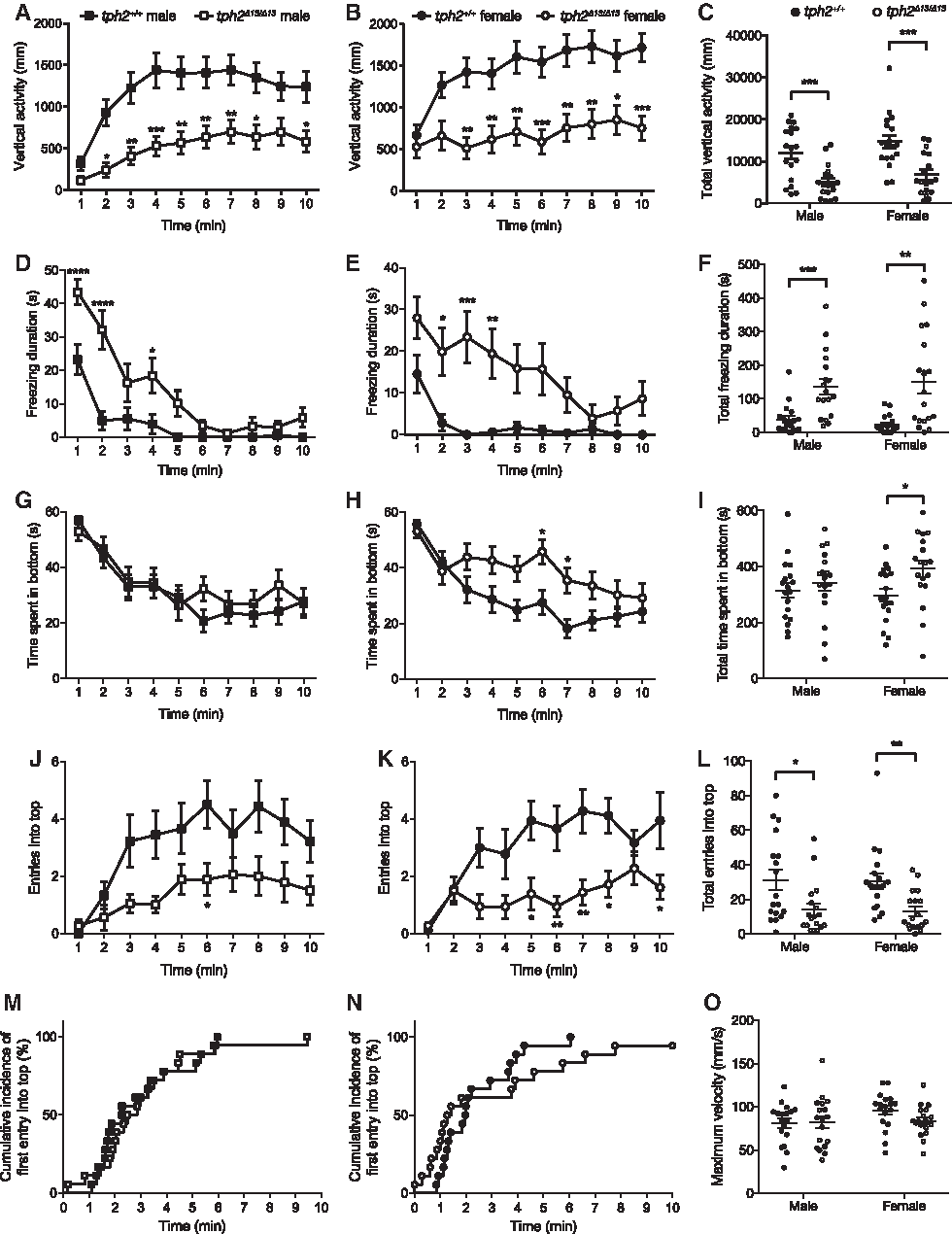
Behavioral phenotypes of tph2Δ13/Δ13 fish tested in the diving test.
In the open-field test, there were no significant differences between the two genotypes in the horizontal swimming activity (Fig. 5A), time spent in the center area (Fig. 5B), and numbers of entries into the center (Fig. 5C). The durations of freezing behavior were significantly increased by the mutation during the first 1 or 2 min in both males (Sidak's multiple-comparison test: 0–1 min, t = 3.651, p = 0.0043; 1–2 min, t = 3.710, p = 0.0034; Fig. 5D) and females (Sidak's multiple-comparison test: 0–1 min, t = 9.639, p < 0.0001; Fig. 5E), whereas the total duration of the freezing was significantly increased in the mutant female (Fig. 5F), but not in the mutant male (Fig. 5F). There were no differences in the maximum swimming velocity in the 15 min test between the genotypes (male: t31.379 = 0.6793, p = 0.5019; female: t17.42 = −0.8593, p = 0.3962; Fig. 5G). The statistical results of Welch's t-test and two-way RM ANOVA are shown in Supplementary Table S1.
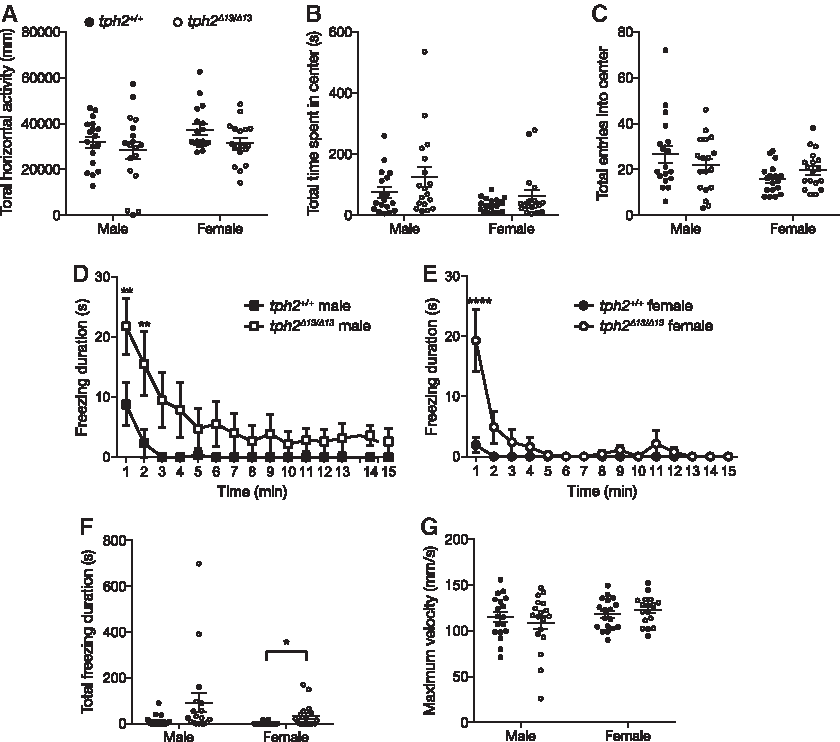
Behavioral phenotypes of the tph2Δ13/Δ13 fish tested in the open-field test.
In the light–dark transition test, the tph2 genotype had no significant effect on time spent in both light (Fig. 6A) and dark areas (Fig. 6B), numbers of transitions between the areas (Fig. 6C), latency to the first entry into the dark area (males: χ2 = 0.9656, p = 0.3258, Fig. 6D; females: χ2 = 0.8650, p = 0.3523, Fig. 6E), and the total swimming activity in the 10 min test (Fig. 6F). The freezing duration during the first 1 min was significantly increased in both tph2Δ13/Δ13 males (Sidak's multiple-comparison test: 0–1 min, t = 5.792, p < 0.0001; Fig. 6G) and females (Sidak's multiple-comparison test: 0–1 min, t = 3.286, p = 0.0113; Fig. 6H). Total duration of freezing behavior in the 10 min test was significantly increased in males but not in females (Fig. 6I). For the statistical results of Welch's t-test and two-way RM ANOVA, see Supplementary Table S1.

Behavioral phenotypes of tph2Δ13/Δ13 fish tested in the light–dark transition test.
Subsequently, alterations of social behaviors were measured with two behavioral paradigms. In the mirror-biting test, the tph2Δ13/Δ13 fish did not exhibit any significant changes in the duration of mirror-biting in the 10 min test (Fig. 7A), total swimming activity in the test (Fig. 7B), and latency to the first mirror bite (males: χ2 = 3.674, p = 0.0553; females: χ2 = 0.8650, p = 0.6065; Fig. 7C). The number of mirror bites was significantly decreased by the mutation in males (Sidak's multiple-comparison test: 1–2 min, t = 3.152, p = 0.0175; Fig. 7D, F) but not in females (Fig. 7E, F). The tph2Δ13/Δ13 fish showed a significantly increased duration of freezing behavior in both males and females (Fig. 7I). The statistical results of Welch's t-test and two-way RM ANOVA are listed in Supplementary Table S1.
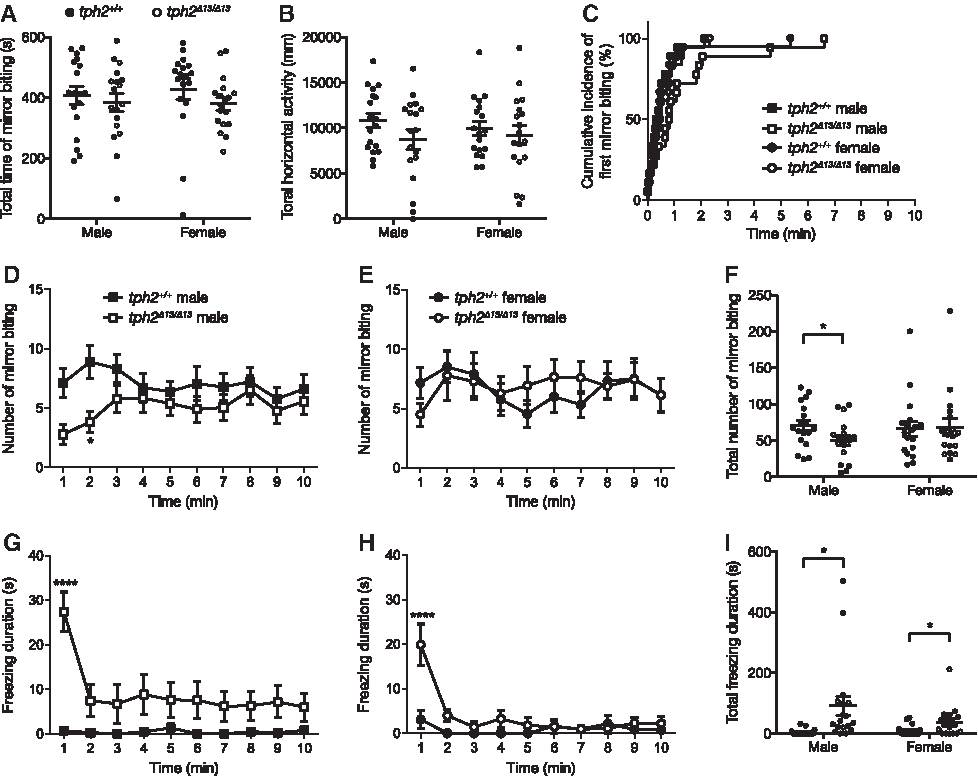
Behavioral phenotypes of tph2Δ13/Δ13 fish tested in the mirror-biting test.
In the two-fish social interaction test, the tph2 genotype did not affect the average distance between individuals for the 10 min test (Fig. 8A), latency to the first contact (males: χ2 = 0.04152, p = 0.8385; females: χ2 = 0.5151, p = 0.4730; Fig. 8B), and total numbers of contacts between the individuals (Fig. 8C). The contact time was significantly increased by the mutation in females (Sidak's multiple-comparison test: 8–9 min, t = 3.590, p = 0.0040; Fig. 8E, F) but not in males (Fig. 8D, F). The statistical results of Welch's t-test and two-way RM ANOVA are shown in Supplementary Table S1.
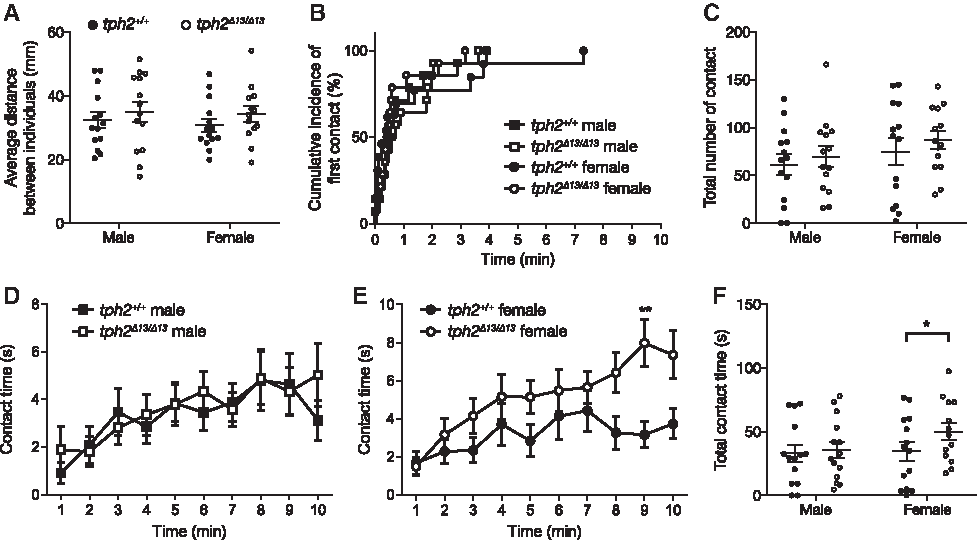
Behavioral phenotypes of tph2Δ13/Δ13 fish tested in the two-fish social interaction test.
Discussion
Previous studies revealed that Tph2-deficient mice were deficient in 5-HT of the raphe serotonergic neurons and exhibited alterations in a wide range of behaviors, such as aggression, depression, anxiety, fear memory, social interaction, and maternal care.31–36 Although teleost fishes also have the serotonergic neurons in the brain, 4 the role of 5-HT as a neurotransmitter in controlling their behaviors was largely unknown because the effects of genetic inhibition of serotonin synthesis have not been analyzed in teleost fishes thus far. In this study, we established a tph2-deficient strain of medaka by targeted mutagenesis using TALENs, which lacked the neurotransmitter 5-HT in serotonergic neurons of the raphe nuclei. Behavioral analysis with five paradigms elucidated that the 5-HT deficiency induced behavioral alterations in the adult fish, including significant increases in the freezing and bottom-dwelling behaviors and decreases in mirror-biting and social interaction. This is the first report that directly shows the effects of 5-HT deficiency in the raphe nuclei on behaviors of teleost fish.
The tph2Δ13/Δ13 fish showed a significantly increased freezing duration, especially during the first minutes of each test, in all four behavioral paradigms that quantify the freezing behavior of each fish (Figs. 4D–F, 5D–F, 6G–I, and 7G–I). Since the hypolocomotion for the first minutes when introduced into a novel environment is considered an index of stress and fear reflexive behavior in medaka, 17 these results suggest that the 5-HT deficiency in the raphe nuclei would induce elevated stress and fear-related behaviors in medaka. In addition to the freezing behavior, an increased diving (or bottom-dwelling) behavior, one of the most reliable indices of elevated stress and fear response in medaka and zebrafish,16,17 was demonstrated by the longer time spent in the bottom area in the tph2Δ13/Δ13 female (Fig. 4H, I) and the decreased number of entries into the top area in both sexes of the mutant fish (Fig. 4J–L). The increased stress and fear-related behaviors in the diving test was also detected in zebrafish injected with a tph inhibitor para-chlorophenylalanine (pCPA), 37 which indicates that the 5-HT deficiency may cause fear- and stress-related responses in a wide range of teleost species.
Such hypolocomotion in the tph2-knockout fish might also result from the impaired mobility. In zebrafish, previous studies showed that tph2 would be involved in 5-HT synthesis in the spinal neurons 38 and the 5-HT could regulate development and regeneration of spinal motor neurons. 39 We found 5-HT-positive neurons in the spinal cord of medaka embryos (data not shown), suggesting that the tph2 mutant medaka has the potential to show dysfunctions in the spinal motor neurons and their motility. In addition, the impaired motility can be caused by spinal deformity at the adult stage (Fig. 1). Although spinal deformity caused by 5-HT deficiency was also reported in chum salmon (Oncorhynchus keta),40,41 the effects on motor functions of teleost species have been unknown. In the open-field, light–dark transition, and mirror-biting tests, the tph2 homozygous mutation had significant effects on freezing duration but not on total swimming activity (Figs. 6A, 7F, and 8B). The mutant fish displayed a decreased vertical swimming activity in the diving test (Fig. 4C), whereas there were no significant differences in maximum swimming velocity of each fish in the diving test (Fig. 4O) as well as the open-field test (Fig. 5G). These results support the fact that the tph2 mutation does not show severe defects in motor function of the medaka. However, further analysis, such as quantification of the stress level by cortisol measurement and assessment of functions of the spinal motor neurons, will be required to investigate the relationship between the hypolocomotion and the elevated fear and stress levels.
Previous studies showed that inhibition of 5-HT synthesis by pCPA administration affected light–dark preference of zebrafish. 37 Also, optogenetic manipulations of tph2 serotonergic neurons altered the light–dark preference in zebrafish larvae. In this study, although we examined the effects of the tph2 mutation in the light–dark preference of medaka, we could not find any significant changes (Fig. 6). In principle, the light–dark transition paradigm can measure the anxiety level as the conflict between spontaneous exploratory behavior and innate aversion to a brightly illuminated environment that is widely found in zebrafish and other teleost species.18,27 However, both wild-type and mutant fish spent longer in the light area than the dark (Fig. 6A, B), which indicates that medaka originally lost the innate aversion of illuminated environments. This suggests that, unlike mice and zebrafish, it would be difficult to measure the anxiety level of medaka by the light–dark paradigm. Interestingly, loss of the innate aversion to illuminated environments was also reported in other fish species, such as bluegill (Lepomis macrochirus), 27 Cardinal-tetras (Paracheirodon axelrodi), 28 and mosquitofish (Gambusia holbrooki). 18 Thus, this finding emphasizes the importance of species differences in neurobehavioral studies using teleost species.
Behavioral assessment in the Tph2 knockout mice showed that the raphe serotonergic neurons were also involved in social behaviors, such as increased aggressive behaviors31,33,35 and autism spectrum disorders-like behaviors (social impairments, communication deficits, and repetitive behaviors). 34 The serotonergic system is known to be involved in social behaviors of teleost species, such as social status in a cichlid fish (Astatotilapia burtoni) 42 and evolutionary shift of aggressive behavior in blind cavefish (Astyanax mexicanus). 43 In this study, significant changes by the tph2 homozygous mutation were detected in both the mirror-biting test and the two-fish social interaction test. These results indicate that the 5-HT deficiency also has some effects on socially evoked behaviors of medaka adult fish. Remarkably, the effects, including a decreased number of mirror-biting (Fig. 7D–F) and an increased time of contact between individuals (Fig. 8D–F), differed between sexes, which suggests that the regulation of social behaviors by the serotonergic neurons might differ between sexes. Although there is little known about the mechanism of the sex differences, increased plasma testosterone level induced by the Tph2 mutation, found only in male but not in female mice, 33 suggests that sex steroids might be involved in the sex-specific effects of the mutation.
It is currently well known that the raphe serotonergic neurons control sleep–awake behaviors in mammals. 44 In fish, although a previous study exhibited that the raphe neurons could modulate sensory responsiveness during aroused states, 13 the effects on the sleep–awake behaviors and their diurnal rhythm of behavioral activity have remained largely unknown. To reveal the effects of the tph2 mutation on the diurnal activity pattern, basal behavioral activity of the mutants will need to be assessed by another behavioral paradigm such as 24 h of a home-tank test.
Recent studies demonstrated that medaka can serve as a suitable model to study the genetic and neuronal mechanisms of complex adult behaviors, such as mating preference, mate-guarding, aggression, and startle response. However, it remains largely unknown as to whether 5-HT and serotonergic neurons are involved in those behaviors. The tph2 mutant strain established in this study will become a good model to elucidate the serotonergic regulations of behaviors and other complex traits in medaka.
Footnotes
Acknowledgments
The authors thank Dr. Masatomo Tagawa for support in histological analysis and Drs. Tomonori Deguchi and Susumu Uji for helpful discussions. They also thank Drs. Tetsushi Sakuma and Takashi Yamamoto for pFUS_A2A and pFUS_A2B vectors, Dr. Kazuyuki Hoshijima for the pCS2TAL3DD and pCS2TAL3RR vectors, Dr. Daniel F. Voytas for the Golden Gate TALEN and TAL Effector Kit (Addgene #1000000016), Dr. Akihiro Fujikawa for advice on micro-computed tomography imaging, and National BioResource Project medaka for the hatching enzyme. This work was supported by a Grant-in-Aid for Japan Society for the Promotion of Science (JSPS) Fellows (no. 13J01682 to S.A.) and the National Institute for Basic Biology Individual Collaborative Research Project (no. 14-345 to M.K.).
Disclosure Statement
No competing financial interest exists.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.