
Abstract
Abstract
Pseudoloma neurophilia, a microsporidium that primarily infects neural tissues, is a common pathogen in laboratory zebrafish. The risk of parasite transmission with different spawning apparatuses and the effectiveness of disinfection are unknown. In this study, we spawned uninfected zebrafish with P. neurophilia-infected zebrafish in either 50 L mass spawning chambers (MSCs) or 1 L standard breeding tanks (BTs). Fish were spawned once or thrice, with and without chamber disinfection between uses, to evaluate risk of vertical and horizontal transmission. Six disinfection protocols were tested to determine which effectively eliminated residual spores. We demonstrated that three consecutive uses of an MSC significantly increased the risk of transmission to other fish when compared to the use of BTs or only one spawning event in an MSC (both p < 0.0001). Vertical transmission was not detected with any method. Disinfection with ∼100 ppm bleach soak (pH ∼7.0), 75 ppm Wescodyne® soak, and 175 ppm Wescodyne Plus spray was 100% effective in eliminating spores from the MSCs. Disinfection of MSCs before spawning did not decrease P. neurophilia transmission when infected fish remained present in the breeding population. Researchers should avoid using endemically infected fish in MSCs to minimize transmission of pathogens within their colonies.
Introduction
Z
The demand for synchronized zebrafish embryos to fulfill research objectives prompted the development and use of mass spawning chambers (MSCs) that maximize the reproductive potential of adult zebrafish. MSCs hold large numbers of zebrafish, and some create shallow water conditions to promote group mating and production of thousands of embryos.3,4 Fish originating from different tanks are temporarily co-housed in the MSC to spawn in recirculating, flow-through, or static water.
Pseudoloma neurophilia, a microsporidium that primarily infects neural tissues, is one of the most common pathogens present within contemporary zebrafish colonies causing chronic infections.5,6 Disease can be subclinical or may present as emaciation, skeletal deformities, and/or death. 7 Aggregates of infectious spores form multiple parasitic clusters (PC) in the hindbrain and ventral aspect of the spinal cord, but have occasionally been observed in other locations, including the somatic musculature and ovary. 8 Inflammation is usually absent or minimal in the brain and spinal cord, but severe, chronic inflammation can be present when free spores are dispersed within other infected tissues, such as the skeletal muscle, swim bladder, and ovary.6,7,9,10
P. neurophilia infection in a research zebrafish facility was first described in 1980 by de Kinkelin and since then, awareness, colony health monitoring, and the availability of sensitive diagnostic tools for detecting this pathogen, such as real-time polymerase chain reaction (PCR), have continued to increase.7,11–13 A recent global survey revealed that 7 of 12 zebrafish facilities reported the presence of microsporidian organisms, including P. neurophilia, during the previous year. 5 Initially P. neurophilia was thought to have a narrow host range as it was only observed in zebrafish; however, it is now known that it can infect other species, including the goldfish (Carassius auratus). 14 It remains unclear whether P. neurophilia is a natural pathogen of zebrafish, since infection has not been described in wild-caught zebrafish. Although there is no effective treatment, the disease can be diagnosed using histopathology and/or PCR.15,16
Horizontal transmission of P. neurophilia occurs through cannibalism of fish carrying mature infective spores, exposure to contaminated tank debris, cohabitation with infected fish, and exposure to infective spores released during spawning events.8,9,13 P. neurophilia may also be transmitted vertically to offspring when spores are present within the developing embryos or on the outer surface of the embryo's chorion.7,8,17 Therefore, the use of MSCs, which hold large numbers of fish when compared to the use of smaller breeding tanks (BTs), could potentially increase the transmission of P. neurophilia. Infected fish could release numerous spores during spawning, infecting greater numbers of fish and embryos. Subsequent cohorts spawned in the MSC could also be infected if the contaminated chamber is inadequately or improperly sanitized. The risk of cross-contamination with MSCs has not been evaluated, nor has an effective disinfection protocol been described or evaluated.
Given the high prevalence of P. neurophilia in laboratory zebrafish, the aims of this study were to determine if P. neurophilia transmission rates differ when using MSCs versus BTs; ascertain if risk correlated to the number of spawning events; develop an effective disinfection protocol for MSCs; and evaluate whether disinfection of MSCs between uses decreases transmission. We hypothesized that the use of MSCs would result in an increased risk of P. neurophilia transmission to unaffected adult zebrafish and their offspring compared with BTs, and that this risk would increase with additional spawning events; the use of chemical disinfectants, including bleach, Virkon® Aquatics, Wescodyne®, and Wescodyne Plus for a contact time of 10 min, would effectively eliminate all P. neurophilia spores; and the risk of P. neurophilia transmission to unaffected adult fish and their offspring subjected to recurrent spawning events could be mitigated by adequate disinfection of the spawning apparatus between uses.
Materials and Methods
Fish: conventional AB
Wild-type zebrafish embryos of the AB line were obtained from the Zebrafish International Resource Center (ZIRC), Eugene, Oregon, at 48 h postfertilization (HPF). Health surveillance reports from ZIRC indicate that this colony is endemically infected with P. neurophilia and Mycobacterium spp., and are free of cestodes, encysted metacercariae (digenetic trematodes), fungal organisms, gram-negative bacteria, Pseudocapillaria tomentosa, Pleistophora hyphessobryconis, Ichthyophthirius multifiliis, Myxidium streisingeri, Piscinoodinium sp., and Tetrahymena sp. Before shipment, embryos were surface sanitized at ZIRC using 50 ppm sodium hypochlorite for a 10-min contact time. Upon receipt, embryos were maintained in 60 × 15 mm Petri dishes filled with an embryo medium (15.8 ppm sea salt [Instant Ocean® Aquarium Sea Salt Mixture; Instant Ocean Spectrum Brands, Blacksburg, VA] in reverse-osmosis [RO] water) and 0.15 ppm methylene blue, and incubated at 28°C until 5 days postfertilization (DPF). At 5 DPF, larvae were moved into 2.8 L tanks on a recirculating rack (Aquaneering, Inc., San Diego, CA) in a quarantine facility, where they were maintained until study termination.
Fish: specific pathogen-free AB
Wild-type zebrafish embryos of the AB line were obtained from the P. neurophilia-specific pathogen-free (SPF) zebrafish colony housed at the Sinnhuber Aquatic Research Laboratory (SARL) at Oregon State University (OSU), Corvallis, OR, at 48 HPF. Health surveillance reports from SARL indicate that this colony is endemically infected with Mycobacterium chelonae and is free of P. neurophilia, P. hyphessobryconis, P. tomentosa, M. streisingeri, Piscinoodinium sp., Edwardsiella ictaluri, and other Mycobacterium spp. All embryos were bleached and reared as described above for the conventional AB fish. At 5 DPF, larvae were housed in tanks on a dedicated stand-alone recirculating rack (Tecniplast, Exton, PA) in our zebrafish core facility until study termination.
All research procedures were approved by the Memorial Sloan Kettering Cancer Center (MSK) IACUC. MSK's animal care and use program is AAALAC accredited and operates in compliance with the Guide for the Care and Use of Laboratory Animals 8th edition.
Housing and husbandry
All adult zebrafish were housed at a density of 5–10 fish/L in mixed-sex groups in either the quarantine facility (conventional) or on a dedicated rack (SPF) in a core facility, with a 14:10-h light:dark cycle. Both systems are recirculating water holding systems that are supplied with calcite-filtered RO/deionized (DI) water with 15% (conventional) or 50% (SPF) water exchanges per day. Recirculated water is passed through either 25-μm (conventional) or 50-μm (SPF) mechanical filters, carbon filters, biological filtration using either a fluidized bed (conventional), or ceramic beads (SPF), and is exposed to UV lamps providing a minimum of 100,000 (conventional) or 180,000 mW/cm2/s (SPF). Water quality parameters on both systems were maintained at 27–29°C, with conductivity 700–900 μS, pH 7.2–7.8, ammonia <0.2 ppm, nitrite <0.1 ppm, and nitrate <30 ppm.
Starting at 5 DPF, zebrafish larvae were fed twice daily with an irradiated <50 μm larval powder diet (Larval AP100 < 50 μm; Zeigler Brothers, Gardners, PA). At 15 DPF, the larvae were fed twice daily with an irradiated 100–150 μm larval powder diet (Larval AP100 100–150 μm; Zeigler Brothers) and thrice daily with Artemia nauplii (Brine Shrimp Direct, Ogden, UT). At 30 DPF, fish were transitioned onto an irradiated commercial pelleted diet (Adult Zebrafish Complete Diet; Zeigler Brothers) and Artemia nauplii fed twice daily.
Establishing P. neurophilia-infected fish
A colony of P. neurophilia-infected zebrafish (n = 200) was established by exposing naive 3-month-old or 7-month-old conventional zebrafish to P. neurophilia-infected central nervous system (CNS) tissue-contaminated water during feeding. The infected CNS tissue was harvested from adult fish from a known infected colony located at OSU or from fish exposed to effluent water at MSK, which has historically been a source of P. neurophilia-infected fish. Nervous tissue harvest was performed using methods described by Murray et al., except CNS tissue was collected from fresh (not frozen) euthanized fish and forceps were not disinfected between fish. 7 The tissue was then cut into 2–3 mm long pieces, placed in 2.0 mL gamma-irradiated screw-cap microcentrifuge tubes (VWR, Radnor, PA), and mixed with 1.5 mL RO water. CNS tissue, suspended in ∼6.0 mL RO water, was mixed with 6.0 mL of brine shrimp and added to the exposure tank twice daily for 2 days. During exposures, the fish were housed in a static 9.5 L tank (Aquaneering, Inc., San Diego, CA) with complete water changes every 24 h. Approximately 15 weeks postexposure (WPE), a subset of fish (n = 75) was euthanized by exposure to 2–4°C water for 10 min and tissues evaluated microscopically to confirm the prevalence of P. neurophilia.
The remaining fish (n = 125) were utilized in Aims 1 and 3 (described below). Over the span of 14 months, 30 P. neurophilia-infected zebrafish were euthanized (as described above) before the scheduled experimental endpoint (at conclusion of Aim 3) due to the development of clinical signs of disease (i.e., emaciation and difficulty swimming), and were evaluated by histology to determine prevalence of P. neurophilia. A total of 51 P. neurophilia-infected fish were found dead over the course of the study and could not be evaluated microscopically due to autolysis. At the end of the study, the remaining infected fish (n = 44) were euthanized (as described) and tissues evaluated microscopically to determine the final P. neurophilia prevalence.
Animal identification
To differentiate between infected (conventional inoculated) and uninfected (SPF) adult zebrafish, the P. neurophilia-infected zebrafish were identified with a visible implantable elastomer tag (Northwest Marine Technology, Shaw Island, WA) near the base of the dorsal fin ∼4–6 months after P. neurophilia exposure following the instructions for minimal volume use (Fig. 1).18,19 The anesthetic solution bath (100 mg/L) consisted of Tricaine-S (MS 222; Western Chemical, Inc., Ferndale, WA) that was added to system water, buffered to a pH of 7–7.8 (Sodium Bicarbonate Certified ACS; Fischer Chemical, Fair Lane, NJ), and maintained at room temperature.

Gross image of tagged Pseudoloma neurophilia-infected fish. Dorsal view of female (top) and male (bottom) with an elastomer tag near the dorsal fin (white arrows). Color images available online at www.liebertpub.com/zeb
Postfiltration sentinels
To ensure that tank-to-tank transmission of P. neurophilia did not occur during the study, four tanks, each containing 6–15 experimentally naive SPF zebrafish, were placed in random locations on the SPF rack housing experimental animals to act as postfiltration sentinels. At the conclusion of both Aim 1 and Aim 3, naive SPF zebrafish (n = 15) were randomly collected and submitted, and assessed microscopically for P. neurophilia.
P. neurophilia quantitative PCR assay
Extraction of DNA and quantitative PCR (qPCR) were performed according to the methods of Sanders and Kent. 13 Water filter samples were processed by using sterile forceps to transfer each filter to a 15 mL conical tube containing 7.0 mL of acetone. The tubes were vortexed periodically over 15 min until the nitrocellulose filter dissolved, centrifuged for 10 min at 3000 g, and the acetone supernatant decanted. The pellet was resuspended in 800 μL of 100% ethanol, vortexed, and the suspension transferred to a 2.0 mL screw-cap tube. Tubes were centrifuged at 12,000 g for 3 min, ethanol supernatant carefully removed by pipette, and the pellet air dried for ∼10 min. For embryo samples, any water remaining was removed by pipetting. Embryos and the dissolved filter sample pellets were placed in 2.0 mL screw-cap tubes with 200 μL of phosphate-buffered saline and sonicated on a Branson Sonifier 250 (Branson, Danbury, CT) at 15% amplitude (5 W) for 5 min. Subsequently, the Qiagen DNeasy Blood & Tissue Kit (Qiagen, Inc., Valencia, CA) was used to extract DNA, first adding 180 μL of ATL and 35 μL Proteinase K, and digesting overnight at 45°C as described by the manufacturer. Extracted DNA was aliquoted into strip tubes and the concentration measured using a DNA spectrophotometer (NanoDrop™ Technologies, Wilmington, Delaware). Quantitative PCR was run on a QuantStudio® 3 Real-Time PCR machine (Applied Biosystems®, Foster City, CA) using primers (PN10F/PN10R and probe) as described by Sanders and Kent, added to TaqMan® Environmental Master Mix 2.0 (Applied Biosystems). 13 All samples were run in triplicate and for each plate, a dilution series (50 fg/μL, 5 fg/μL, 500 ag/μL, 50 ag/μL, and 5 ag/μL) of a synthetic target P. neurophilia fragment (gBlocks® Gene Fragments; Integrated DNA Technologies, Coralville, IA) was run in triplicate as a standard.
Histopathology
Fish were euthanized as described above. The coelomic cavity was incised and the fish fixed in Bouin's solution for 48 h. Fish were sectioned longitudinally along the midline, processed, and embedded in paraffin tissue blocks. Two 5-μm thick serial sections, taken 100 μm apart, were deparaffinized and stained with Luna stain using Luna's method for staining erythrocytes and eosinophil granules. 20 Tissues were evaluated for evidence of intraneural and extraneural P. neurophilia infection using a binocular light microscope (Olympus Corporation, Shinjuku, Tokyo, Japan) by S.P. Tissue sections in which P. neurophilia infection was not observed were examined a second time by a board-certified veterinary pathologist (J.W.).
Experimental design
AIM 1: P. neurophilia transmission without spawning device disinfection
Five-month-old uninfected zebrafish (n = 120) were randomly assigned into one of four groups. Seven-month-old P. neurophilia-infected zebrafish (n = 60) were randomly assigned and used in at least two of the following groups below:
(1) Group BTx1 (n = 30 infected and 30 uninfected fish). These fish were randomly assigned into 24 separate 1.0 L standard BTs (Aquaneering, Inc., San Diego, CA) at a density of 2 or 3 fish/L and allowed to spawn once. (2) Group BTx3 (n = 30 infected and 30 uninfected fish). These fish were randomly assigned into 24 separate 1.0 L standard BTs and spawned every 7 days for a total of three spawnings. (3) Group MSCx1 (n = 30 infected and 30 uninfected fish). These fish spawned once in a custom-made 50 L acrylic MSC (dimensions: 47.5″ × 11″ × 8″; Aquaneering, Inc., San Diego, CA) (Fig. 2) at a density of 2.5 fish/L. The bottom of the chamber was layered with red glass marbles (1/2″ diameter) to minimize consumption of embryos, and a valve was mounted at the bottom side of the tank to facilitate drainage. (4) Group MSCx3 (n = 30 infected and 30 uninfected fish). These fish spawned once every 7 days for a total of three spawnings in a 50 L MSC at a density of 2.5 fish/L.

Spawning apparatuses used in the study.
While all MSC groups included at least 30 infected fish (n = 15 males, n = 15 females), fish in the BT groups were randomly assigned from either the infected or uninfected colony, to reflect common laboratory practices. Following randomization and placement in tanks, a visual count of elastomer tags confirmed that 40%–60% of the BTs included at least one infected and uninfected fish. Each BT contained at least one male and female fish. The chamber/tanks were not disinfected between use; they were drained and air-dried. For all groups, the fish were placed in their respective BT or MSC in the afternoon and males and females physically separated overnight by a polyvinyl chloride divider, which was removed the following morning and the fish allowed to spawn for 60 min.
The presence of P. neurophilia in uninfected fish was evaluated at two time points, baseline and 15 WPE, at which time, 13–15 adult uninfected zebrafish per group were euthanized and submitted for analysis. Fish examined at baseline were collected from the BT or MSC immediately following the last spawning event. In addition, the water from each BT and MSC was filtered through a 3″ nylon mesh strainer (HIC Harold Import Company, Lakewood, NJ) to collect embryos following each spawning event. For the last spawning event, 3159, 1155, 1459, and 571 embryos were collected from groups BTx1, MSCx1, BTx3, and MSCx3, respectively. Fifteen pooled embryo samples from each group were randomly collected and evaluated by qPCR to test for the prevalence of P. neurophilia. Each pooled sample contained 15 embryos and was placed into a 2.0 mL screw-cap microcentrifuge tube and stored at −20°C until analyzed. Fifty embryos from each group were also randomly collected and maintained until they were 15 weeks of age. The remaining embryos that were collected and not reared were euthanized using a dilute bleach solution (one part 8.25% bleach [Clorox Concentrated Germicidal Bleach, Oakland, CA] to five parts housing system water). Embryos remained in this solution for at least 5 min to ensure death.
AIM 2: MSC disinfection
Due to the higher risk of P. neurophilia transmission detected in MSCs from Aim 1, only disinfection protocols for MSCs (and not BTs) were evaluated. MSCs were contaminated with P. neurophilia before disinfection by spawning P. neurophilia-infected zebrafish (n = 20), which had not spawned for at least 7 days before use, in 8.0 L of system water within one of three MSCs. After spawning, the water from each MSC was completely drained and filtered through a 3″ nylon mesh strainer, to remove large debris and embryos. The water was then filtered through a 1.2 μm nitrocellulose filter (RAWG04700; EMD Millipore Corporation, Darmstadt, Germany) using a 4.0 L filtering flask (Fisher Scientific, Hampton, NH). The filters were removed using sterile forceps or hemostats, rolled and placed into a 2.0 mL screw-cap microcentrifuge tube, and stored at −20°C until analyzed by qPCR to confirm the presence of P. neurophilia within the chambers. Embryos were euthanized as previously described. The drained MSCs were randomly assigned to one of six disinfection protocols:
(1) Air dry; The MSC was allowed to dry at room temperature for 60 min. (2) RO water soak. (3) 90–120 ppm sodium hypochlorite solution soak: the sodium hypochlorite solution was prepared by adding 80–100 mL 8.25% bleach (Clorox® Concentrated Germicidal Bleach, Oakland, CA) to 50 L RO water. The solution was buffered to a pH of 6.8–7.4 with pH Down Water Conditioner (API Fishcare, Chalfont, PA), using a pH probe (Hanna HI98130 pH/Conductivity/TDS tester, Woonsocket, RI). The chlorine concentration was verified using a portable chlorine meter (Extech CL200, Camarillo, CA) and chlorine test strips (LaMotte, Chestertown, MD). (4) 1.0% Virkon Aquatics (Syndel Laboratories, Ltd., Ferndale, WA) solution soak: prepared according to manufacturer's instructions with RO water.
21
(5) 75 ppm Wescodyne (STERIS Corp., Mentor, OH) solution soak: prepared according to manufacturer's instructions with RO water.
22
(6) 175 ppm Wescodyne Plus (Fischer Scientific, Hampton, NH) solution spray: prepared according to the manufacturer's instructions with RO water and administered from a 16-ounce spray bottle, which allowed the solution to be dispersed on the MSC's inner surfaces.
23
All solutions were freshly prepared on the day of the study, and the study was repeated with each disinfectant eight times. Soaks were performed by filling the entire chamber (50-L) with each solution, allowing it to stand for 10 min. The MSC was then drained and its inner surfaces rinsed thrice with 10–15 L RO water to remove residual disinfectant. The chambers were then air dried at room temperature for 60 min. The spray method involved spraying the chamber's inner surfaces with the disinfectant, allowing 10 min of contact time. The inner surfaces were subsequently rinsed and the chamber was allowed to air dry as described above.
Following disinfection and drying, the MSC was filled with 8 L RO water to obtain a postdisinfection water sample. The water remained in the chamber for 1 min, and was subsequently decanted, filtered (1.2-μm nitrocellulose filter), and the filter analyzed by qPCR for P. neurophilia.
AIM 3: P. neurophilia transmission following MSC disinfection
Uninfected (n = 60; 12 months old) and P. neurophilia-infected (n = 60; 14 months old) zebrafish were randomly assigned into one of two groups.
(1) Group DMSCx1 (n = 30 infected and 30 uninfected fish). These fish spawned once in a 50 L MSC at a density of 2.5 fish/L.
(2) Group DMSCx3 (n = 30 infected and 30 uninfected fish). These fish spawned once every 7 days, thrice in a 50 L MSC at a density of 2.5 fish/L. The MSC was disinfected with 175 ppm Wescodyne Plus as described in Aim 2 immediately after each spawning event.
The presence of P. neurophilia in the uninfected fish was determined as described in Aim 1. Uninfected zebrafish (n = 12–15/group) were euthanized and submitted for histology at two time points (baseline and 10 weeks postexposure). Fish examined at baseline were collected from the MSC immediately following the last spawning event. After the last spawning event, embryos were collected and 15 pooled samples of 15 embryos each were collected and tested for P. neurophilia by qPCR as described in Aim 1. Furthermore, 50 embryos from each group were randomly selected and maintained until they were 15 weeks of age, at which point, they were evaluated for P. neurophilia microscopically as described in Aim 1. All remaining embryos were euthanized as described above.
Statistics
Differences between groups were evaluated using Fisher's exact test for categorical variables and the Kruskal-Wallis test for continuous variables. Differences in gene copy number between predisinfection and postdisinfection samples were evaluated using the Wilcoxon signed-rank test. All analyses were performed using SAS version 9.4 (SAS Institute, Inc., Cary, NC). p Values less than or equal to 0.05 were considered statistically significant.
Results
Infected colonies: P. neurophilia prevalence
Microscopic evaluation of the subset of the infected fish colony revealed that 63 of 75 fish (84%) were infected with P. neurophilia. All positive fish showed evidence of intraneural infections, with PC in the brain, spinal cord, and/or nerve roots. Twenty-nine of the 63 positive fish (46%) had evidence of extraneural infections, with spores and PC in the skeletal muscle, kidney, testis, and/or ovary (Fig. 3).
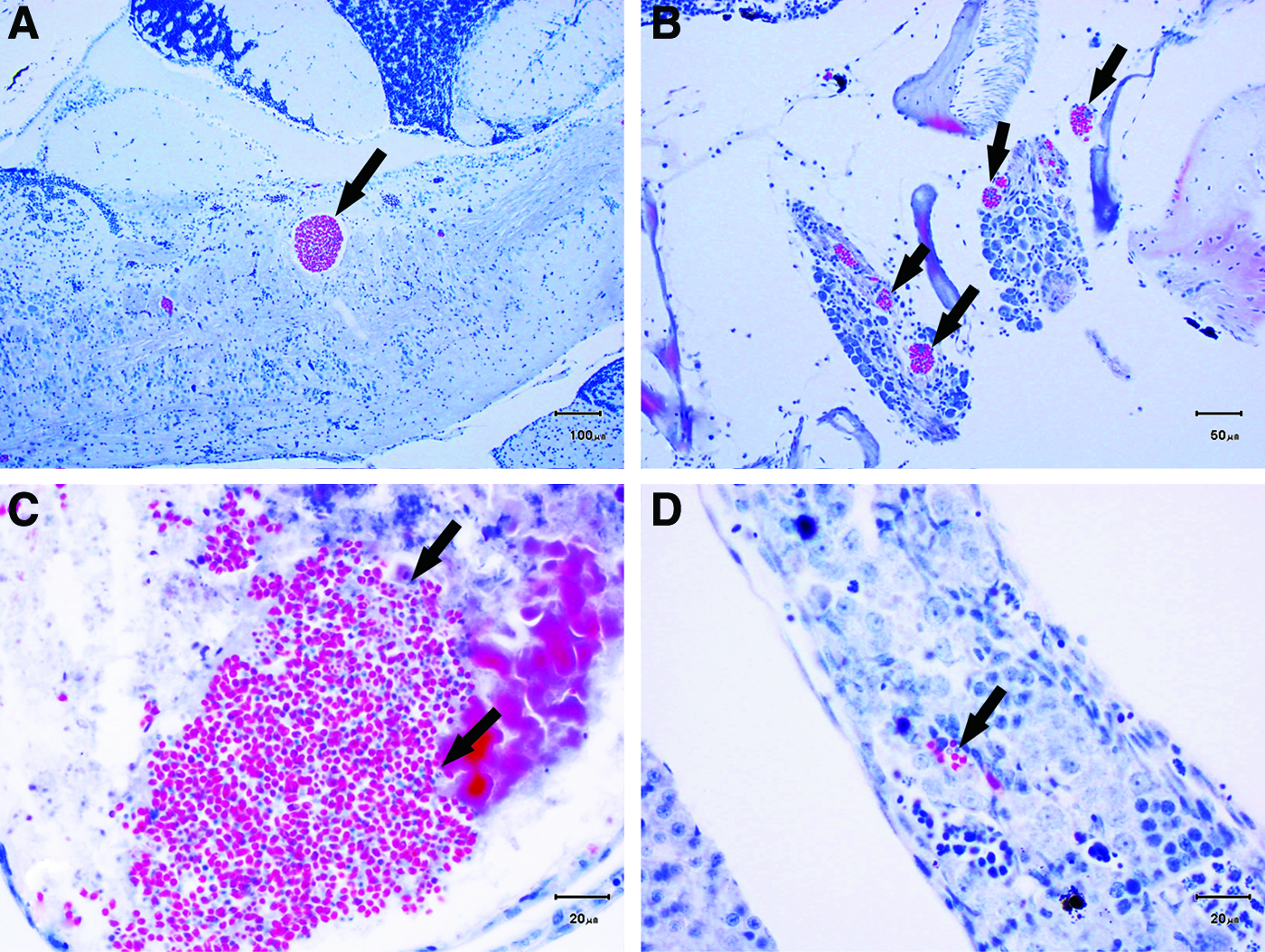
Histologic characteristics of intraneural and extraneural P. neurophilia in the P. neurophilia-infected colony.
Twenty-nine of the 30 fish (96.7%) that required euthanasia due to clinical signs showed histologic evidence of P. neurophilia infection. All positive fish showed evidence of intraneural infections, whereas 62% (n = 18) of the positive fish also had evidence of extraneural infections, with PC and free spores present in the kidney, ovary, skeletal muscle, and/or notochord.
At the conclusion of the study, the fish remaining from the infected colony (n = 44) were euthanized (46–80 weeks postinfection), to determine final P. neurophilia prevalence. All fish had evidence of neural infections, and of those, 38.6% (n = 17) had evidence of extraneural infections, with PC and free spores present in the kidney, notochord, muscle, and/or ovary.
Postfiltration sentinel evaluation
None of the experimentally naive SPF zebrafish placed as sentinels had microscopic evidence of P. neurophilia infection.
AIM 1: P. neurophilia transmission without spawning device disinfection
SPF fish from all groups were negative for P. neurophilia histologically at the baseline evaluation. At 15 WPE, none of the fish evaluated microscopically in groups BTx1 or BTx3 were found to be P. neurophilia positive. One (6.7%) of the group MSCx1 and all of the group MSCx3 SPF fish were P. neurophilia positive with evidence of intraneural infections. In addition, 4 (30.7%) of the MSCx3 fish had evidence of extraneural infection at 15 WPE with P. neurophilia present in the kidney, ovary, and notochord. When comparing the 15 WPE results from group MSCx3 with either group MSCx1 or BTx3, the differences seen were statistically significant (Fisher's exact test, both p < 0.0001). At baseline, one pooled embryo sample from group BTx1 was P. neurophilia positive by qPCR. The remaining pooled embryo samples across all groups were negative for P. neurophilia. The embryos that were allowed to hatch and maintained to 15 WPE were negative for P. neurophilia when examined microscopically.
AIM 2: MSC disinfection
The predisinfection water samples (n = 8 per protocol) for all six disinfection protocols were P. neurophilia positive as determined by qPCR. Seven of eight (87.5%) of the water samples were P. neurophilia positive after the MSCs were allowed to dry for 1 h (control). For the RO soak, four of eight (50%) of the postfiltration water samples were positive for P. neurophilia, while one of eight (12.5%) of the postfiltration samples was positive for P. neurophilia following a Virkon Aquatics soak. None of the postfiltration samples (n = 8 per group) were P. neurophilia positive, following the bleach soak, Wescodyne soak, and Wescodyne Plus spray. The presence of P. neurophilia, as determined by qPCR, was compared to postdisinfection among all six groups, with each method compared to the control. When comparing the qualitative data, the difference between the Virkon Aquatics soak and the control was statistically significant (p = 0.01; Fisher's exact test). In addition, the bleach soak, Wescodyne soak, and Wescodyne Plus spray were all statistically significant when compared to the control (p = 0.001; Fisher's exact test). The postdisinfection difference between the control and the RO soak was not different (p = 0.28; Fisher's exact test).
P. neurophilia mean gene copy numbers for the predisinfection and postdisinfection water samples are provided in Figure 4. To determine if quantitative postdisinfection water samples could be compared to one another, the variability in predisinfection samples was evaluated. The mean predisinfection gene copy number present among all groups was significantly different (p = 0.001; Kruskal-Wallis test), preventing postdisinfection comparison among groups. The gene copy number differences between predisinfection and postdisinfection samples were compared for each individual group. The reduction in gene copy numbers was statistically significant in all groups (p = 0.05 for the control, p = 0.02 for the Virkon Aquatics soak, and p = 0.008 for the bleach soak, Wescodyne soak, and Wescodyne Plus spray; Wilcoxon signed-rank test).

P. neurophilia gene copy numbers (mean ± SEM) present in MSCs before and after disinfection using one of six protocols as determined by quantitative polymerase chain reaction. Reduction in gene copy number was statistically significant (*p = 0.008, #p = 0.02, **p = 0.05, Wilcoxon signed-rank test) for each group. SEM, standard error of the mean. Color images available online at www.liebertpub.com/zeb
AIM 3: P. neurophilia transmission following MSC disinfection
All fish in both group DMSCx1 and DMSCx3 were negative for P. neurophilia as assessed microscopically at baseline. Adult fish were sacrificed 10 WPE, as opposed to 15 WPE in Aim 1, due to increased mortality seen between 8 and 10 WPE. At 10 WPE, only group DMSCx3 had P. neurophilia-positive fish (100% infected). All the positive fish had evidence of both intraneural and extraneural infection with P. neurophilia observed in the kidney, ovary, intestine, and within a blood vessel. When comparing group DMSCx3 to DMSCx1, the differences seen were statistically significant (Fisher's exact test; p < 0.0001). All embryos tested from both groups were qPCR negative at baseline. At 15 WPE, there was no evidence of infection in any of the fish tested.
Discussion
This study demonstrated that spawning AB zebrafish three times in MSCs carries an increased risk of P. neurophilia transmission to uninfected fish. In contrast, spawning AB fish in BTs, once or thrice, did not result in an increased risk of P. neurophilia transmission. Therefore, we evaluated the effectiveness of six disinfection protocols in only the MSCs. All MSCs became contaminated with P. neurophilia when infected fish were bred. Only bleach soak, Wescodyne soak, and Wescodyne Plus spray were able to eradicate P. neurophilia spores in all trials from the MSCs as tested by qPCR of postdisinfection water samples. However, disinfection of MSCs with Wescodyne Plus did not mitigate the risk of P. neurophilia transmission when spawning infected AB zebrafish with uninfected fish.
Aim 1 was designed to simulate a real-life setting where the P. neurophilia status of each fish is usually unknown. In the MSC, the uninfected fish had direct exposure to infected fish and water. Unlike commercially available BTs and MSCs, which contain slotted inserts or egg collector trays that effectively prevent consumption of embryos and feces, the red glass marbles present at the bottom of the custom-made MSC were ineffective in preventing exposure to feces and embryos as fish were able to move between marbles.3,4 P. neurophilia is not commonly detected in the feces and embryos of infected fish; thus, the transmission of this organism in the custom-made MSC should not have been greatly impacted by allowing the naive fish to have direct exposure to those materials. 24 Even if some fish did not spawn within the custom-made MSC, they were most likely exposed to spores that may have been released and consumed during urination and/or spawning.9,17 Given the differences between the design and structure of a custom-made and commercially available MSCs, however, future studies should determine if this increased risk still remains when using the latter.
BTs are typically used for spawning pairs or small groups of fish. 25 In Aim 1, although 40%–60% of tanks contained at least one infected and one uninfected fish, none of the BT groups showed evidence of P. neurophilia transmission. One possible reason for this observation is that not every infected fish releases spores into the water during spawning, especially if only intraneural infections are present. In a previous study, Sanders et al. demonstrated that only 40.7% of BTs housing breeding pairs had evidence of P. neurophilia spores in the water by qPCR, although each pair contained at least one P. neurophilia-infected fish and embryos were collected from all tanks. 17 In this study, all fish in the infected colony showed evidence of intraneural infection, while only 38.6%–62% displayed P. neurophilia spores in extraneural tissues, such as the ovary and testes, which could contribute to antemortem transmission. Ovarian infections may be associated with high numbers of P. neurophilia spores because developing ova are considered an immune-privileged site. 26 Consequently, it is probable that a limited number of P. neurophilia fish (with heavy extraneural infections) could contribute disproportionately to the spore burden in the MSCs. These heavy shedders have the potential to transmit disease to more fish in an MSC than when spawned in small group sizes in a BT. In addition, the potential for having larger numbers of infected fish and hence a potentially higher number of P. neurophilia spores is greater when using an MSC, allowing greater numbers of naive fish to become exposed and infected. It has been demonstrated that 300 or 500 spores/mL are needed to infect 5 DPF larval AB zebrafish; infection has not been observed with exposure to lower spore concentrations.9,17 It is therefore conceivable that a minimum number of spores and/or repeated exposures are needed to establish P. neurophilia infection in adult fish. Several of these factors may have contributed to the lack of P. neurophilia transmission between fish in the BTs.
Six disinfection protocols were evaluated to determine which are effective at eliminating P. neurophilia spores from the inner surfaces of the MSCs. Only the MSCs were evaluated in this aim since there was an increased risk of P. neurophilia transmission during consecutive spawning events when they were used. The 10-min contact time used was based on manufacturers' recommendations.22,23 It may be valuable to determine if shorter exposure times can yield similar results. Some facilities air dry MSCs after use to save time and money; however, this study highlighted that in almost every trial, spores were still detectable in the water using this method. Infectious microsporidial spores have a thick, chitinous endospore, making them highly resistant to environmental stress and lysis. 2 It is unknown how long P. neurophilia spores remain viable on surfaces, but viability can be maintained for extended periods in aquatic environments, such as within water droplets. 15 Cleaning MSCs with RO water alone removed spores in half of the trials, which can likely be attributed to mechanical rinsing and removal of the organisms from the tank.
The remaining four disinfection protocols utilized disinfectants commonly employed in zebrafish facilities (personal communications). Virkon Aquatics is a concentrated disinfectant, containing potassium peroxymonosulfate and sodium chloride, which is advertised to be effective against various aquatic bacterial, viral, and fungal agents.21,27,28 In this study, Virkon Aquatics at a 1.0% concentration was relatively effective in eliminating P. neurophilia spores; however P. neurophilia nucleic acid was detected postdisinfection in a single trial. The failure in the single trial may have resulted from incomplete mixing as Virkon Aquatics is sold as a powder. Moreover, the presence of organic debris at the bottom of the MSC may have prevented adequate penetration of the disinfectant, or may have neutralized one or both of the active ingredients.
Sodium hypochlorite (commonly known as bleach) is effective against common bacteria, viruses, parasites, and fungi. 28 P. neurophilia spores are highly resistant to chlorine, requiring >100 ppm to achieve >95% spore death. 29 In addition, buffering chlorine solutions to a pH of 7.0 increases the germicidal properties and activity of chlorine due to increased presence of hypochlorous acid (HOCI).28,30 The bleach soaks at 90–120 ppm buffered to pH of 6.8–7.4 were highly effective in eliminating P. neurophilia spores. Buffers and meters should be utilized to ensure that proper chlorine concentration and pH are achieved before use. In addition, given that bleach can be highly toxic to aquatic animal species, users should be cautious when working with this chemical to prevent accidental contamination of nearby water systems and tanks. 28
Wescodyne® is a broad-spectrum iodophor-based disinfectant that contains iodine as its active ingredient. 22 Although iodophors are generally used to disinfect nonporous environmental surfaces, they have also been utilized in aquaculture for disinfecting embryos.22,28,31 Wescodyne Plus is a concentrated solution of 175 ppm iodine that is formulated for easy mixing and includes a spray attachment. 23 While both 75 ppm Wescodyne and Wescodyne Plus were effective at eliminating P. neurophilia spores from the inner surfaces of MSCs, the spray feature of the latter allows users to save both time and water.
By virtue of its effectiveness in eliminating the P. neurophilia spores and with the goal of water conservation, Wescodyne Plus was selected for use in Aim 3. Aim 3 was conducted to determine if the increased risk of P. neurophilia transmission associated with repeated use of MSCs could be mitigated by disinfecting the chambers between uses. If residual spores from previous spawning events contributed to the increased infection rates after three spawning events, we would have expected a lower transmission rate with adequate disinfection between uses. In this aim, both infected and uninfected fish were allowed to spawn in an MSC either once or thrice, following use of 175 ppm Wescodyne Plus. Despite successful disinfection of the chamber as shown in Aim 2, P. neurophilia transmission to uninfected adult zebrafish persisted, with transmission rates identical to those observed in Aim 1 (100% infection after three spawning events with or without disinfection). This suggests that transmission associated with the use of MSCs occurs primarily through direct contact with infected fish by exposure to their urine and/or reproductive materials. Therefore, spawning of infected fish in MSCs should be avoided to minimize transmission of P. neurophilia within colonies.
Feces, urine, skin, and water can remain on the inner surfaces of MSCs after spawning. These materials can act as fomites for the transmission of various infectious organisms, if the MSC is inadequately disinfected. For example, the eggs of the zebrafish nematode, P. tomentosa, can be transmitted to other fish through the presence of contaminated feces. 32 P. neurophilia spores can remain within water droplets and adhere to the inner surfaces of MSCs after spawning. The spores of another microsporidian, P. hyphessobryconis, may be released and transmitted from the skin and urinary tract of infected fish. 33 Last, Mycobacterium spp. may be shed from skin lesions and feces, and can remain infective to other fish populations through the formation of biofilms. 34 Therefore, disinfection of spawning devices between uses is advisable to decrease the transmission of infectious organisms.
For Aims 1 and 3, none of the offspring showed microscopic evidence of P. neurophilia when reared to adults. During each spawning event, the number of embryos acquired from the groups varied, but typically ranged from 500 to 5000 embryos (data not shown). The relatively small sample size (n = 240) of the total number of embryos generated may have limited our ability to detect vertical transmission within these populations. The results from our study are consistent with previous publications that demonstrated that the risk of vertical transmission is low (∼0%–1%) in breeding populations, although not impossible, and therefore should remain a consideration throughout facilities.17,24
To determine whether transmission of P. neurophilia to uninfected fish occurred during spawning, rather than from tank-to-tank transmission on a recirculating rack, postfiltration sentinels were placed and evaluated for P. neurophilia status. None of the sentinels showed microscopic evidence of P. neurophilia infection, despite the presence of numerous P. neurophilia-positive fish in other tanks on the rack. Although the efficacy of UV sterilization against P. neurophilia has not been reported, effective doses for other microsporidia vary from 6 to 283,500 mW/s/cm2 depending on microsporidian species. 7 These results support previous recommendations to maintain effective UV sterilization on recirculating racks to minimize the transmission of microsporidiosis.7,11,35 In contrast, the prevalence of P. neurophilia within the infected colonies continued to increase throughout the study. For example, all the fish submitted from the infected colony at the conclusion of the study had evidence of P. neurophilia, despite the initial estimated prevalence of ∼84% at 15 weeks postexposure. Any uninfected fish that remained in tanks with the infected fish most likely acquired P. neurophilia during cohousing and spawning within the tank in which they were housed.
Although this study demonstrated an increased risk of P. neurophilia transmission to uninfected AB zebrafish when using MSCs thrice, we were unable to mitigate the risk using an effective disinfectant. Wescodyne Plus (with a 10-min contact time) successfully eliminates P. neurophilia spores from MSCs as evidenced by a lack of detection in all postdisinfection water samples through qPCR, while also saving time and money. However, 100% of uninfected fish still acquired the infection when spawning in MSCs thrice, despite disinfection between uses. This is most likely attributed to the fact that infected fish were present in the breeding population. We were unable to evaluate the significance of residual spores in P. neurophilia transmission to naive fish in this study because the uninfected fish had continual direct contact with infected fish. However, the prevalence of P. neurophilia is low to absent in feces and embryos of infected fish; thus, direct contact with infected fish themselves is likely to be a more common method for transmission in this setting. 24 Further studies are required to determine the true impact of environmental spores on P. neurophilia transmission by spawning only uninfected fish in a P. neurophilia-contaminated MSC.
P. neurophilia is one of the most prevalent infectious organisms present within research zebrafish populations today and has been shown to impact research in a multitude of ways, despite its common subclinical presentation. 10 P. neurophilia spores aggregate within brain and spinal cord structures and have been shown to impact startle responses, shoaling behavior, and anxiety responses in zebrafish.10,36,37 These infection-induced behavioral alterations can have negative ramifications for investigators who utilize zebrafish for behavioral and neuroscience studies. 36 It has also been demonstrated that P. neurophilia-infected zebrafish exposed to gamma irradiation experienced increased parasite loads and higher mortality than fish exposed to either P. neurophilia or gamma irradiation alone, highlighting the effect that P. neurophilia can have on immunosuppressed animals. 38
This study is the first report to show histological evidence of P. neurophilia in the testis of an infected zebrafish. Previous publications have been unable to identify P. neurophilia within the testicular parenchyma and only showed evidence of spores within the connective tissue adjacent to the testes by histology or in sperm samples submitted for PCR.2,7,13,17 The observation of P. neurophilia in testicular tissue suggests that infected sperm may contribute to horizontal and vertical transmission. This new finding may pose a special problem for cryopreservation, as zebrafish lines are cryopreserved as sperm. It is likely that P. neurophilia can survive the cryopreservation process, since other microsporidia can be maintained long periods in tissue culture by cryopreservation. 9
In conclusion, this study demonstrated that spawning fish multiple times in MSCs increased the risk of P. neurophilia transmission to uninfected zebrafish. In facilities enzootically infected with P. neurophilia, the cohousing of fish from different tanks in an MSC will likely increase the prevalence of P. neurophilia within the colony. Therefore, knowledge of the health status of fish utilized for breeding is important to decrease P. neurophilia transmission, especially when working with immunocompromised lines. Other methods for decreasing transmission of P. neurophilia include the use of P. neurophilia-free zebrafish for breeding stock, rapid removal of sick fish from holding tanks, and culling aged (>1 year) fish from breeding populations.7,11,39 We demonstrated that P. neurophilia can remain on the inner surfaces of MSCs after spawning and drainage; therefore, it is prudent to disinfect spawning apparatuses (MSCs and BTs) between use to decrease the transmission of P. neurophilia and other infectious organisms.
Footnotes
Acknowledgments
We thank Mark Francis, Aquaneering, Inc., for donating the MSCs and Dr. Michael Kent and his colleagues for the infected CNS tissue. We also express gratitude to Jacqueline Candelier, Elizabeth Conner, and Kenny Lüøñg from the Laboratory of Comparative Pathology for their assistance with necropsy, K. Alice Wood Fox from the Center for Applied Microbiology at SUNY-ESF for technical support with the qPCR, and Lee-Ronn Paluch for her review of the article. This research study was funded, in part, through the NIH/NCI Cancer Center Support Grant P30-CA008748 and NIH Research Education Grant (5R25OD010447-03). C.M.W. was supported by the Office of Research Infrastructure Programs of the NIH under a subcontract of the award number R24OD010998. The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIH.
Disclosure Statement
No competing financial interests exist.