
Abstract
Abstract
The zebrafish, Danio rerio, has become recognized as a valuable model for infectious diseases. Here we evaluated the susceptibility of zebrafish to be infected with the mammalian vesicular stomatitis virus (VSV). Both zebrafish cells and embryos were highly susceptible to VSV infection. Mortalities exceeded 80% in infected embryos and were preceded by the invasion of the central nervous system by VSV. Live imaging of the infection with GFP-VSV as well as virus titration from infected fish confirmed the viral replication. Immunohistochemical analysis of embryonic fish provided evidence of viral antigens as well as of the apoptosis marker caspase-3 in the brain, eye, liver, pronephros, and skeletal muscle. So far, this is the first report describing the susceptibility of zebrafish to the mammalian virus VSV.
Introduction
V
VSV most prominently produces a vesicular disease in domestic animals such as cattle, horses, or sheep. 10 The virus can spread quickly and even though VSV does not usually cause animals to die, it does cause significant economic losses to livestock producers. VSV transmission between hosts occurs via insect vectors 11 or contaminated surfaces. 12 Although natural infections in humans are rare, vesicular stomatitis is a zoonotic disease and there are reported cases of people infected through natural infection.13,14 The VSV serotypes involved with disease in livestock are New Jersey and Indiana subtypes 1, 2 and 3. The clinical disease in these animals is characterized by vesicles in the tongue, feet, prepuce, and teats (reviewed in Letchworth et al. 15 ). In addition, VSV can cause neurotoxicity in mice, nonhuman primates, and humans. 16 This VSV tropism driving the virus to the brain has been used by some researchers to evaluate brain-related phenomena. 17
VSV establishes noncytopathic, lifelong persistent infections and the virus can be transmitted transovarially from infected females to their offspring in insects. 18 In contrast, mammalian infections are typically acute and cytolytic, and cleared in a few days. 15
Recently, zebrafish Danio rerio has emerged as a new model for infectious agents and host/pathogen interactions. Although no natural infections of virus have been described so far in zebrafish,19,20 different studies have shown that zebrafish can be infected with fish-specific viruses,21–30 and some human viruses such as chikungunya, herpes simplex type I, influenza A, or hepatitis B and C can infect and cause disease in zebrafish.31–35 Zebrafish embryos present a combination of advantages, making it an attractive organism for biomedical research. It is small, cheap, easy to manipulate, and during early embryo and larval stages, individuals are transparent, being highly suited to in vivo imaging.
In the present study, we examined the ability of a recombinant VSV expressing the green fluorescent protein (VSV-GFP) to replicate and cause disease in embryonic zebrafish. Moreover, we also carried out live imaging of VSV-GFP infection in the entire group of animals and analyzed the evolution of infection. Infection of embryonic zebrafish with VSV led to ubiquitous virus replication and apoptosis induction, causing death within 48 h. To our knowledge, this is the first demonstration of the susceptibility of zebrafish to VSV infection.
Materials and Methods
Zebrafish maintenance
The protocols used in this study were performed in compliance with the EU animal experimentation regulation (EU, 2010) and were approved by the Bioethics Committee for Animal Experimentation CEEA-LU (Universidade de Santiago de Compostela, Spain). The zebrafish line used in the experiment was the wild-type AB line. Zebrafish embryos were obtained from breeding of wild-type adult zebrafish. Adult zebrafish were maintained in a controlled environment in a 14-h light/10-h dark cycle at 28°C. 36
Virus and cell cultures
Viral infections were performed with VSV of Indiana strain or a recombinant VSV derived from the Indiana strain expressing the GFP (VSV-GFP). 37 Viruses were grown and virus titers were determined by plaque assay on BSC40 cells. Temperature-inactivated virus was obtained by incubating VSV-GFP virus for 60 min at 60°C.
BSC40 cells were maintained in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS), 100 U mL−1 penicillin, 0.1 mg mL−1 streptomycin, and 2 mM L-glutamine at 37°C and 5% CO2. The zebrafish cell line ZF4 was maintained in DMEM supplemented with 10% FBS, 100 U mL−1 penicillin, 0.1 mg mL−1 streptomycin, and 2 mM L-glutamine at 28°C and 5% CO2.
Viral titration of the extracellular media of ZF4 cells was performed in triplicate on BSC40 cells by standard plaque assay.
Antibodies
Antibody against the G protein of VSV (anti-VSV-G) was a generous gift from Dr. Iván Ventoso (CBM, UAM). The anti-tubulin antibody (MCA78G) was purchased from AbD Serotec and anti-caspase-3 antibody was purchased from Promega.
VSV microinjection in zebrafish embryos
Dechorionated 48 hpf zebrafish embryos were microinjected into the yolk with 15–20 nL VSV-GFP (containing 600, 60, or 6 plaque-forming units [pfu]) in phosphate-buffered saline (PBS). Control groups were similarly injected with PBS. Forty-eight embryos were injected in each condition. Injections were carried out with a Narishige IM-31 microinjector, and borosilicate microneedles (Harvard Apparatus, MA). After microinjection, embryos were distributed in individual wells of 96-well plates, kept at 33°C, and 24 h after infection, embryos were anesthetized with 0.02% tricaine (MS-222; Sigma-Aldrich) and photographed using a fluorescent zoom microscope (Nikon AZ100 Multizoom microscope) and NIS-Elements BR Software (v.4.13.04; Nikon). Half of the embryos were then euthanized and kept frozen (−80°C) until processed and tested for virus by the Reed and Muench method. The remaining embryos were transferred back to the plate for an additional period of 24 h. Embryos were then processed as described above. Clinical signs of infections, including pericardial or yolk edema, tail deviations, or body malformations, were assessed. Experiments were carried out in triplicate.
In addition, 24 embryos were microinjected into the yolk with 600 pfu VSV-GFP per embryo, or PBS as a control, and at 24 h after injection they were processed for histopathological and immunohistochemical studies.
Recovery of infectious GFP-VSV from embryos
Embryos were harvested, resuspended in DMEM, and homogenized. Homogenates were centrifuged for 3 min at 12,000 g and the supernatants were titrated by the Reed and Muench 38 method on BSC40 cells.
Histopathology and immunohistochemistry
Whole embryos were fixed in 10% buffered formalin for 12 h, dehydrated through a graded ethanol series, washed in xylene, and embedded in paraffin wax. Serial sections of 3 μm thickness were dewaxed, rehydrated, stained with hematoxylin and eosin, and immunolabeled for VSV and active caspase-3. For immunohistochemistry, all incubations were carried out at room temperature in a humid chamber. Washes with 0.01 M PBS were performed three times, 5 min each. Antigen retrieval was performed heating the tissue sections under pressure in Target Retrieval Solution High pH (Dako), allowing them to cool at room temperature. The monoclonal mouse anti-VSV-G and the polyclonal rabbit anti-active caspase-3 (Promega) were incubated after quenching endogenous peroxidase activity with the peroxidase-blocking solution (Dako). After the primary antibodies, a secondary antibody conjugated with peroxidase, ImmPRESS VR Reagent anti-mouse IgG and ImmPRESS VR Reagent anti-rabbit IgG (Vector Laboratories), was applied and developed with Vector VIP (Vector Laboratories). Sections were counterstained with hematoxylin. Appropriate positive and negative control sections were included.
Statistical analyses
Student's t-test was performed for viral titration of zebrafish cells and embryos. The significance level chosen for the statistical analysis was p < 0.05. Survival data were plotted using the Kaplan–Meier estimator, and log-rank tests were performed to assess differences between groups. Statistical analyses were performed using SPSS software.
Results
Susceptibility of VSV to replicate in zebrafish cell line ZF4
To evaluate whether zebrafish cells were susceptible to VSV replication, ZF4 cells were infected at 28°C with VSV at a multiplicity of infection (MOI) of 0.05 pfu per cell. At different times after the infection, cellular extracts were collected and the synthesis of the VSV protein G was analyzed by Western blot. We detected expression of the VSV-G protein at 12 h after infection with the virus (Fig. 1A), indicating that these cells were susceptible to be infected by the virus. In accordance with this susceptibility, we also observed that VSV infection caused a classical cytopathic effect characterized by rounded cells and an almost complete destruction of the monolayer at 24 h after infection (Fig. 1B). To further demonstrate the susceptibility of ZF4 cells to VSV infection, we decided to evaluate the virus production in these cells at a comparable level. We infected ZF4 cells with VSV at an MOI of 0.05, and at different times after infection at 28°C, cultures were harvested, centrifuged at 2000 g for 10 min at 4°C, and the virus titers present in supernatants were quantified by plaque formation assay in BSC40 cells. As shown in Figure 1C, we observed an increase in VSV titer reaching a maximum level at 24 hours postinfection (hpi), demonstrating the active replication of the virus. Similarly, replication of the recombinant VSV-GFP in ZF4 cells was also observed (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/zeb).
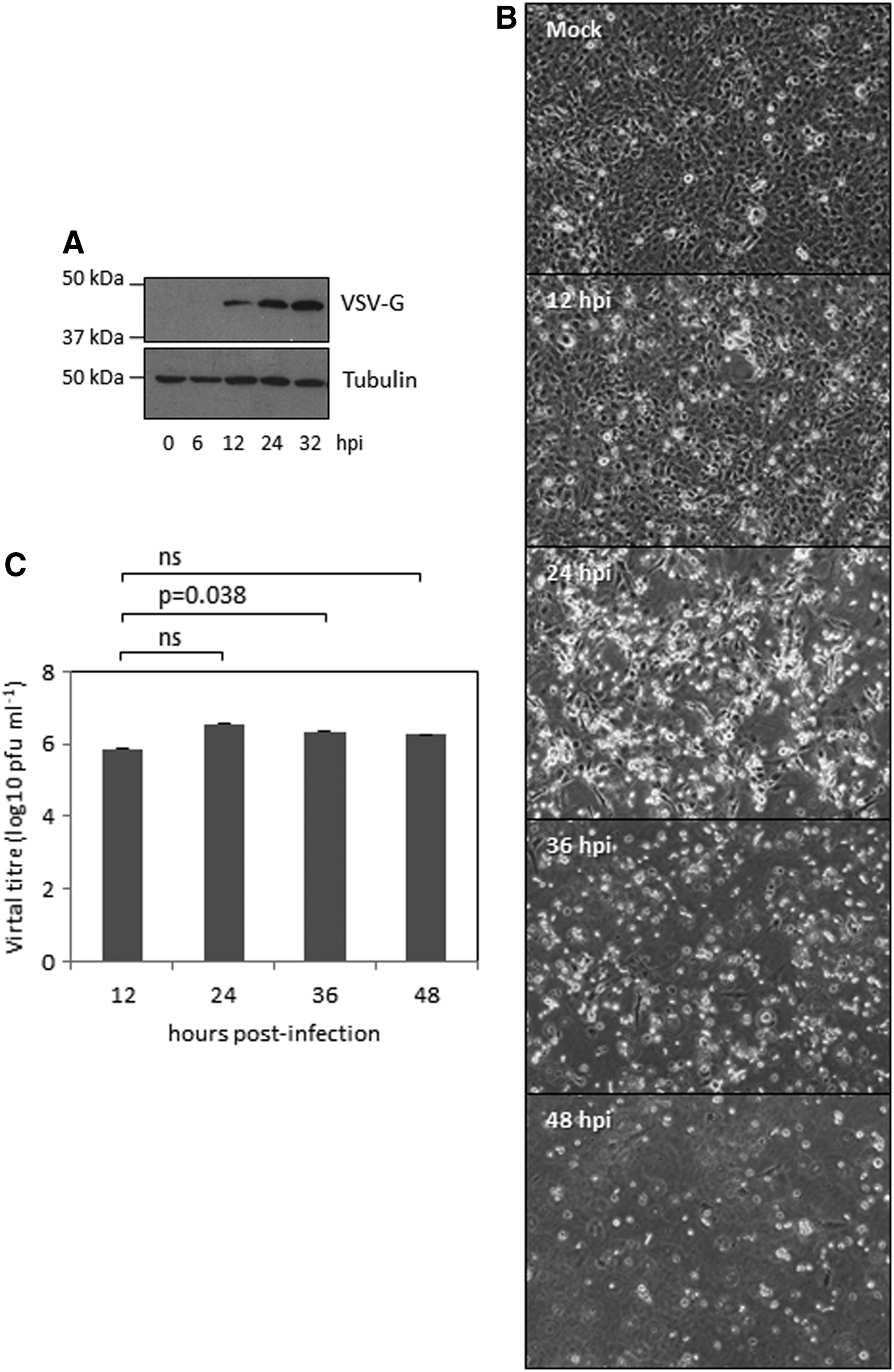
Susceptibility of the zebrafish cell line ZF4 to VSV replication.
Susceptibility of zebrafish embryos to VSV
We then wondered whether zebrafish were also sensitive to VSV infection. Zebrafish embryos were injected into the yolk sac with PBS or different doses (6, 60, or 600 pfu per fish) of VSV-GFP. Injection of PBS, conditioned media derived from BSC40 cells, or with 600 pfu of temperature-inactivated VSV-GFP did not result in zebrafish mortality; however, zebrafish embryos quickly died after injection with VSV-GFP (the LD50 for VSV-GFP was 2 pfu, Fig. 2A). Analysis of the results obtained in three different experiments revealed the existence of a positive relationship between the dose of GFP-VSV injected and the mortality observed. At 24 h after injection, mortality was around 5% among the embryos injected with 6 pfu, 14% among those injected with 60 pfu, and 18% among the embryos injected with 600 pfu GFP-VSV (Fig. 2A). Mortality clearly increased at 48 h after infection, when it reached 57%, 75%, and 87% of the embryos injected with 6, 60, and 600 pfu, respectively (Fig. 2A). In addition, injection of zebrafish embryos with 600 pfu of VSV revealed no significant differences in the resulting mortalities relative to those observed in the experiment carried out with VSV-GFP (Supplementary Fig. S2).
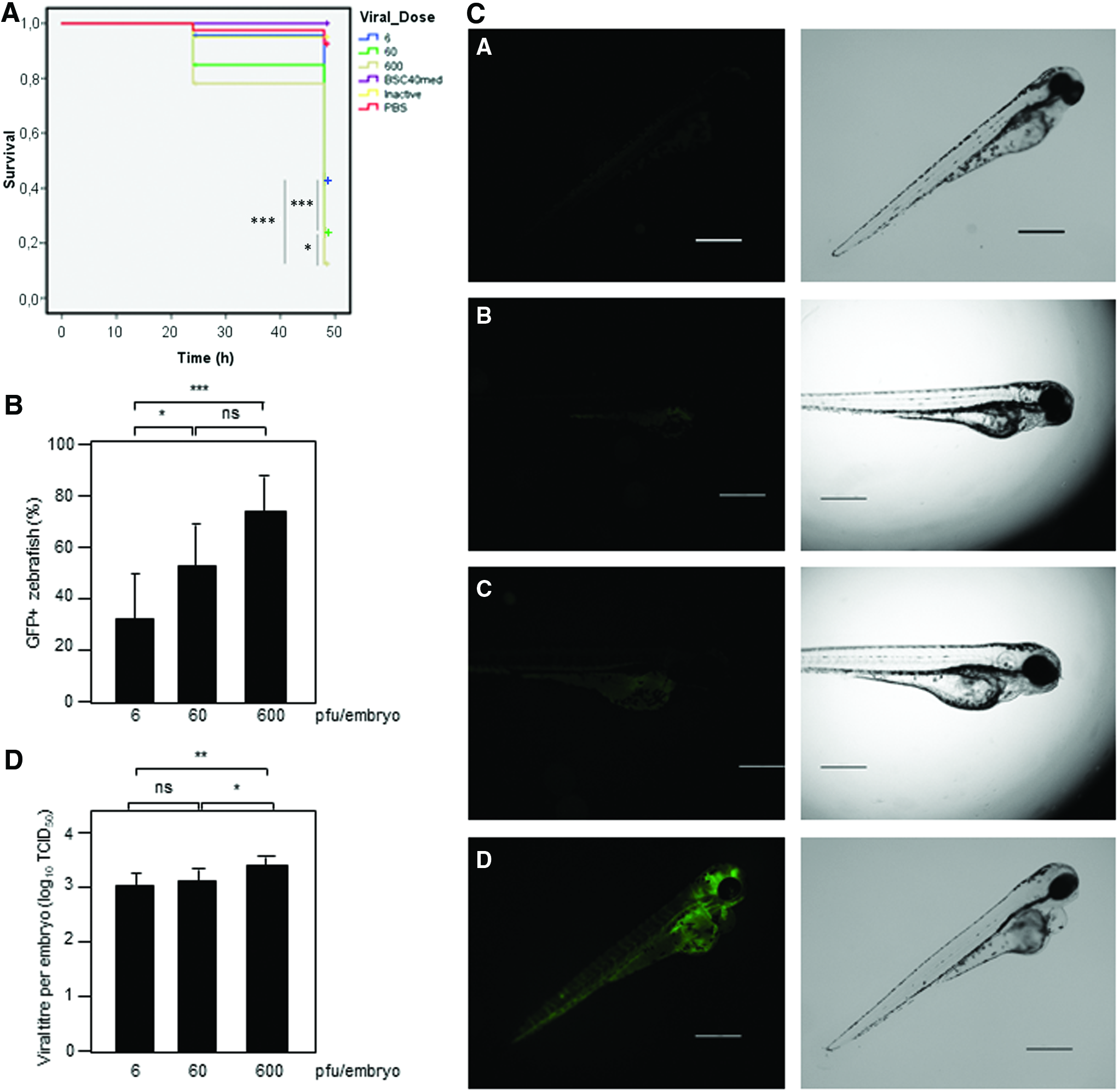
To follow the course of the infection, we evaluated the expression of GFP in the VSV-GFP-infected animals at different times after infection. We detected GFP expression in the infected embryos at as soon as 10 hpi, independently of the viral dose injected. Infectivity rates were directly related to viral burden (0.32 of the embryos infected with 6 pfu, 0.53 of those infected with 60 pfu, and 0.74 of the infected with 600 pfu, at 24 h after infection) (Fig. 2B). A delay in the detection of GFP expression in the VSV-GFP-injected animals but a similar mortality was detected when the animals were maintained at 28.5°C (Supplementary Fig. S3). Interestingly, we did not detect GFP-positive animals or zebrafish mortality after immersion of embryos in a solution containing 1.5 × 107 pfu VSV-GFP mL−1.
The GFP signal was initially detected in the yolk membrane, but rapidly spread in the fish body and mainly to the brain/central nervous system. Later on it was detected in the heart, vascular system, eye, and myosepta, reaching muscle fibers just before the embryos died. The time of development of the infection may vary between individuals. In Figure 2C, different stages of the infection process can be visualized in 24 hpi embryos.
Quantification of the viral burden from zebrafish embryos at 24 hpi revealed a clear increase in the titer of the virus relative to the initially injected virus (Fig. 2D). Moreover, the individual titers recovered from embryos injected with the highest dose were significantly higher than the virus recovered from those injected with lower titers (Fig. 2D). Altogether, these results indicate that VSV can replicate in zebrafish leading to the induction of death.
Histopathology and immunohistochemistry
To further probe the susceptibility of zebrafish embryos to VSV-GFP, we carried out immunohistochemistry analysis of fixed tissues from injected animals with an antibody recognizing the protein G of VSV (VSV-G). We analyzed 10 specimens infected with 600 pfu presenting different viral titers at 24 hpi. As shown in Figure 3A and B, the pattern of VSV-G was similar to the one observed for the GFP signal, indicating that GFP was a reliable indicator of the infection and confirming the susceptibility of zebrafish to VSV. If more definition was needed when using GFP fluorescence imaging, confocal microscopy could be carried out (Supplementary Fig. S4). In all the specimens, immunostaining for VSV-G was especially evident in the central nervous system (Fig. 3C). No immunopositivity against VSV-G was found in control specimens, coinciding with the absence of GFP signal in these zebrafish embryos (Figs. 3D–F).

Signaling of VSV-GFP in zebrafish embryos at 24 h after infection with 600 pfu.
The immunopositivity against VSV-G was distributed from the inoculation point to other tissues (Fig. 4A, B). An intense immunoreaction was observed near the injection point, as well as in the endothelium of surrounding vessels (Fig. 4B). In the areas near the otic vesicle, the reaction showed a reticular pattern occupying the layers beneath the lining epithelium and into the pharyngeal arches (Fig. 4C). The location of VSV-G antigen coincided with an increase in the number of apoptotic figures in the vicinity, detected with immunohistochemistry against active caspase-3 antigen (Fig. 4D), although not all the apoptotic processes can be due to viral infection since basal levels of apoptosis were detected in noninfected individuals (Fig. 4E). Similar findings were found in the endodermal tissue located ventrally to the notochord and dorsally to the yolk sac (Fig. 4F, G). Other tissues with an especially intense reaction were the liver and some tubule cells in the pronephros (Fig. 4H). In the case of the eye, the detection of the VSV-G antigen presented two different distributions: occasionally the periphery of the eye was immunopositive (Fig. 4I), whereas the majority of embryos displayed a reticular staining of VSV-G antigen inside the developing layers of the inner part of the eye (Fig. 5A). Similarly, active caspase-3-positive cells were present in this location (Fig. 5B). In the brain, the reticular pattern was present in the white matter and between the cells of the nervous system. The areas with more intense immunostaining of VSV-G coincided with the detection of higher numbers of cells morphologically undergoing apoptosis (Fig. 5C, D). Finally, the distribution of the VSV-G antigen followed the connective tissue of the myosepta, the endomysium, and the basement membrane of the yolk sac (Fig. 5E). In the periphery of yolk sac, a nonspecific immunoreaction with anti-caspase-3 antibody was observed (Fig. 5F).
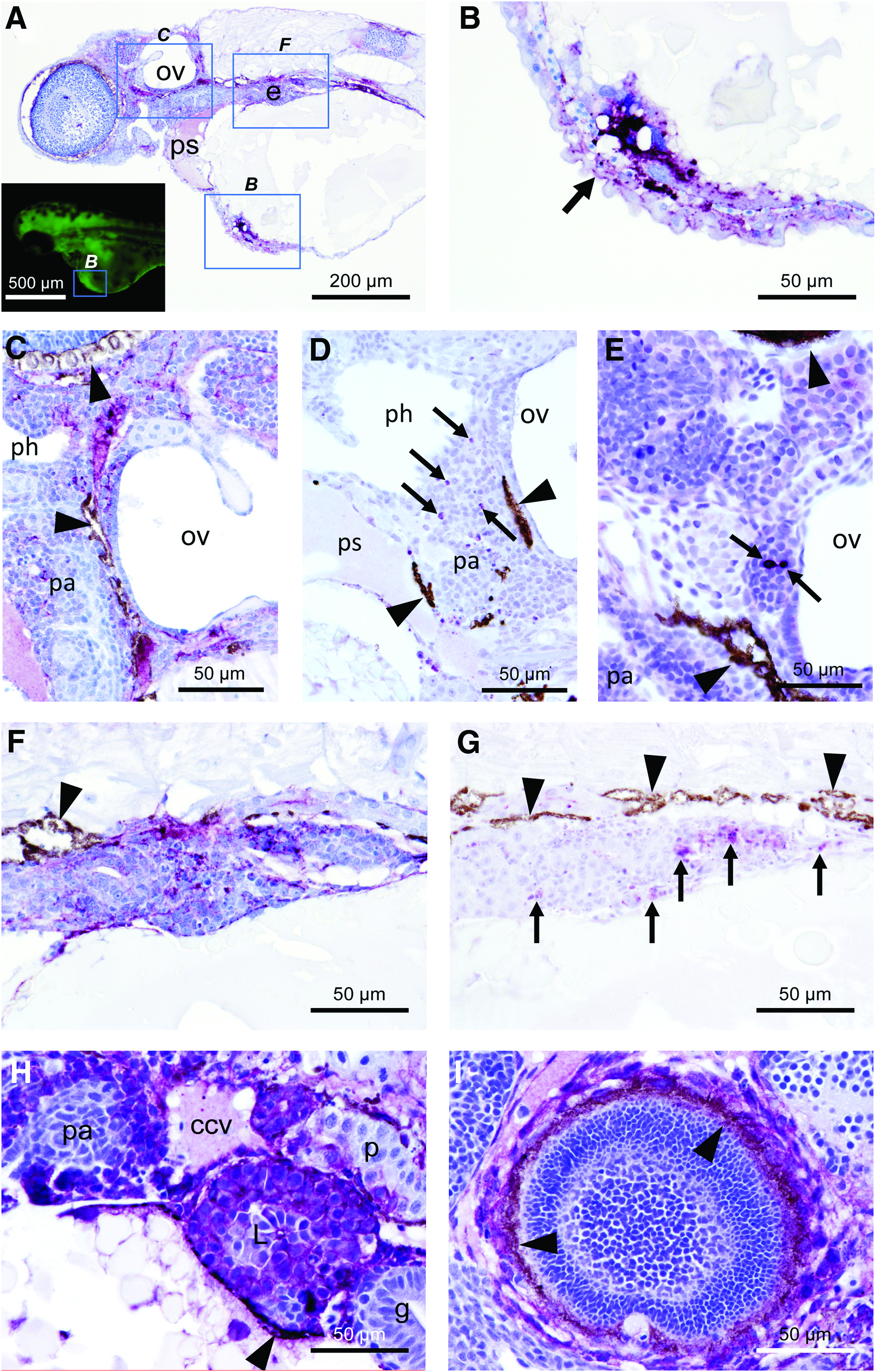
Immunohistochemistry of VSV-G protein
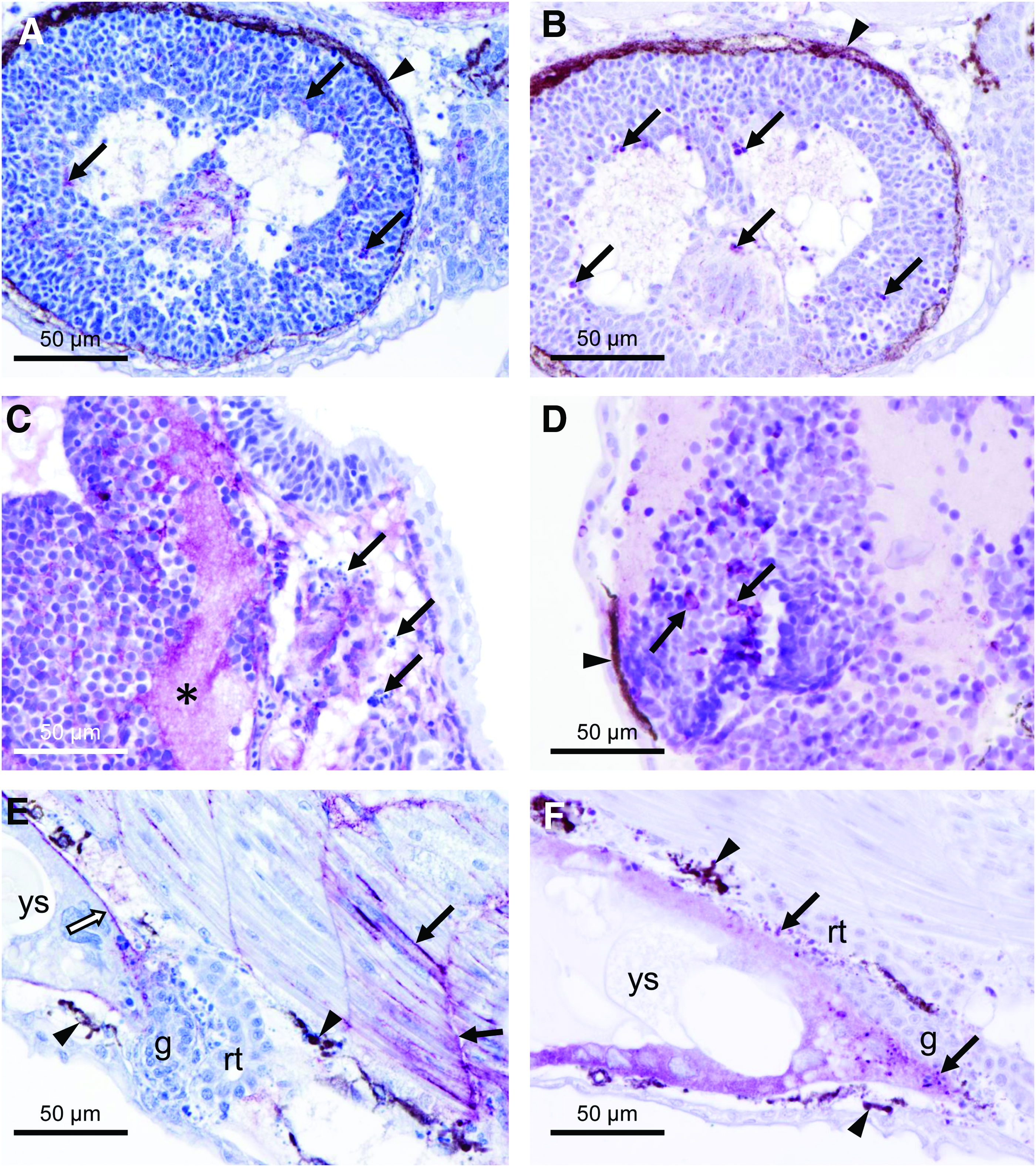
Immunohistochemistry of VSV-G protein
Discussion
Zebrafish has been revealed to be a powerful model system to study infectious diseases, including those caused by virus, and this model has been used to study multiple aspects of viral pathogenesis such as the screening for antiviral compounds, 33 studying viral infection-associated pathologies 32 or evaluating the oncolytic capability of viral agents to induce tumor regression. 39
In this study, we show the successful infection of zebrafish cells with VSV and VSV-GFP as well as the infection and pathogenesis in zebrafish embryos after VSV or VSV-GFP injection. The zebrafish cell line ZF4 was highly susceptible to VSV infection as demonstrated by expression of viral proteins, development of cytopathic effect, and production of infectious virus. Injection of VSV-GFP resulted in mortality of embryo zebrafish and spread from the site of injection to the central nervous system, in agreement with its neurotropic nature.5,40,41 Likely, reaching the central nervous system would be at some extent delayed in later developmental stages due to a more mature blood-brain barrier. Moreover, immunohistochemical analysis of zebrafish embryos using anti-caspase-3 antibody revealed a clear increase in caspase-3-positive cells in VSV-GFP-positive areas, indicative of apoptosis induction, the typical outcome of VSV infection in mammalian cells.42–45 We did not detect zebrafish infection or mortality after immersion of the embryos in a viral solution, suggesting that the virus cannot infect zebrafish larvae by oral or skin route.
The capability of VSV to infect zebrafish cells is in agreement with its pantropic infectivity and with the previously described use of retroviral vectors pseudotyped with the G glycoprotein from VSV for gene transfer in cells from fish.46–48 Moreover, the competence of VSV to replicate at zebrafish temperature (28.5°C) may be related with its ability to replicate in insect vectors.
VSV is a promising candidate as an oncolytic virus and as a vaccine vector,3,4 but strategies to limit its neurotoxicity as well as to enhance its oncolytic activity are still needed for the development of safe and effective gene therapy, vaccines, or oncolytic virus therapy. The results we show here establish the zebrafish embryo as a potential valuable animal model to study VSV oncotropism as well as the neurotropism of the virus.
Footnotes
Acknowledgments
We thank Dr. Ivan Ventoso who generously provided us with the anti-VSV-G antibody and María del Carmen Carreira and Sandra Maceiras for technical assistance. L.S., J.G.-V., M.I.Q., and A.P.L. acknowledge the financial support received from the Xunta de Galicia (Galicia, Spain) under the Grupos de Referencia Competitiva and con Potencial de Crecimiento Programme: Project GRC2014/010 (L.S. and J.G.-V.) and GPC2015/34 (M.I.Q. and A.P.L.). Funding at the laboratory of C.R. is provided by the Ministry of Economy and Competitiveness and FEDER (BFU-2014-58530), the Consellería de Cultura, Educación e Ordenación Universitaria (GI2119 and Centro singular de investigación de Galicia acreditación 2016‐2019, ED431G/05), and the European Regional Development Fund (ERDF). M.C. is a Miguel Servet investigator supported by an ISCIII and EU-FEDER grant (PI14/00554).
Authors' Contributions
Conceptualization: J.G.-V., M.C., C.R., L.S.; methodology: J.G.-V., M.B.-M., S.D.S.-A., A.P.L.; validation: J.G.-V., M.B.-M., A.P.L., M.I.Q., C.R.; formal analysis: J.G.-V., M.B.-M., A.P.L., M.I.Q.; investigation: J.G.-V., M.B.-M., A.P.L., S.D.S.-A.; resources: M.I.Q., M.C., C.R., L.S.; writing—original draft preparation: J.G.-V., M.B.-M., M.I.Q., C.R.; writing—review and editing: J.G.-V., M.B.-M., S.D.S.-A., A.P.L., M.I.Q., M.C., C.R., L.S.; visualization: J.G.-V., M.B.-M., A.P.L., C.R.; supervision: C.R., L.S.; project administration: C.R., L.S.; funding acquisition: M.I.Q., C.R., L.S.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.