
Abstract
Abstract
Fish cellular models are commonly used to study the toxic potential of environmentally relevant compounds. Several of these pollutants act on DNA and compromise its integrity. Little is known, however, about the DNA repair ability of these cellular models. Therefore, the aim of this study was to evaluate the DNA base excision repair (BER) of zebrafish Liver (ZF-L) cell line and primary hepatocytes. We performed kinetic studies of the DNA damage levels after exposure to hydrogen peroxide (H2O2, 20 μM for 10 min) using the Comet Assay. Ten minutes after H2O2 treatment, 16% and 50% of the initial damage, measured as comet tail length, were repaired in ZF-L cell line and primary hepatocytes, respectively. Primary hepatocytes repaired 50% of the damages twice as fast as ZF-L cell line and showed DNA damage levels similar to control 40 min after H2O2 treatment. The total recovery time for ZF-L model was of 180 min, which indicates the culture cells have a less efficient BER. In conclusion, both ZF-L cell line and primary hepatocytes exhibit BER activity; however, these cellular models have different repair capacity. In addition, we demonstrated that ZF-L cell line and primary hepatocytes are useful tools for ecotoxicological studies focusing on DNA single-strand breaks and BER.
Introduction
O
Among the different DNA repair systems, base excision repair (BER) stands out. Studies reveal that alterations and damage in the BER proteins as well as mutations in the genes of this mechanism of repair are found in several types of cancer.4–7 The BER acts on damages such as alkylation and oxidation of bases and single-strand breaks (SSB). In summary, in this type of repair, the altered nucleotide will be removed through the action of enzymes such as DNA glycosylases, which recognize the altered base and remove it. Removal of the base generates apurinic or apyrimidinic sites recognized by the enzymes AP endonucleases and phosphodiesterases, which remove the remaining sugar-phosphate backbone. In the sequence, a DNA polymerase adds the new nucleotide, which is bound to the strand by DNA ligase.8–10
The zebrafish (Danio rerio) is one of the most used models in the world because it has several advantages. This species has been considered useful for studying the DNA damage and repair capacity.11,12 It was considered that DNA repair was in general lower/slower in fish than in mammals. In fact, studies have shown that this activity depends on the DNA repair process analyzed and the developmental phase of the organism.1,13
Another advantage of zebrafish is the existence of in vitro experimental models for this species. Concerning the use of experimental models, in vitro biological models have gained great importance over the years.13,14 The zebrafish Liver (ZF-L) cell line was established from cells isolated from the liver of D. rerio in 1992 and has been used as a model in several toxicological studies and a promising alternative to the use of animals.15,16 Established cell cultures are considered to be very effective in some analysis due to their suitability to mechanistic studies. 14 There is a concern, however, whether cell cultures represent reliable models, since they can exhibit altered functions over time, for example, differences in morphology, development, and gene expression. 17 Despite challenges in the isolation and maintenance, the primary culture cells, such as the commonly used primary hepatocytes culture of D. rerio, are a good alternative. It is assumed that the similarity of the primary hepatocytes to the in vivo condition of the organism of origin allows to perform acute exposure studies.18,19
The increasing use of in vitro models and the importance of maintaining DNA integrity create another challenge, because little is known about DNA repair in fish cellular models used in ecotoxicological or clinical studies. Considering that a few studies have actually compared the activity of the DNA repair between cellular models, the aim of the study was to compare the capacity of DNA BER of the ZF-L cell line and primary hepatocytes through kinetic studies of the DNA damage level after H2O2 exposure.
Materials and Methods
Cell culture
ZF-L cell line
Zebrafish (D. rerio) hepatocytes (ZF-L cell line) were obtained from the American Type Culture Collection (ATCC® number CRL- 2643TM). The cells were maintained in cell culture flasks (Kasvi®) at 28°C and were grown in the LR medium (50% Leibovitz's L-15 [Gibco®; pH 7.4] medium and 40% RPMI 1640 [Gibco; pH 7.4] medium, both supplemented with sodium bicarbonate [0.2 g/L], L-glutamine [0.3 g/L], Hepes [25 mM], 10% fetal bovine serum [Sigma®] and 1% antibiotic and antimycotic [Sigma; penicillin {100 U/mL}, streptomycin {100 μg/mL}, and amphotericin B {0.25 μg/mL}]). The cells (passage number: 25–30) were then plated into 24-well plates (5 × 105 cells/well—for comet assay) or 96-well plates (5 × 104 cells/well—for MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] assay) and maintained for 48 h at 28°C to allow cell attachment.
Danio rerio primary hepatocytes
Adult wild-type zebrafish strains were obtained from Agroloja Red Fish, a supplier in Porto Alegre city (Rio Grande do Sul, Brazil). All experimental procedures were approved by the Ethics Committee on Animal Research of the Federal University of Rio Grande—FURG (CEUA-FURG P065/2015). Culture of adult D. rerio primary liver cells was adapted from a procedure published by Reschly et al. 20 Briefly, D. rerio was sacrificed with MS-222 (250 mg/L) and immersed in 0.5% bleach diluted in Ca2+- and Mg 2+-free phosphate-buffered saline (PBS [Ca2+/Mg2+free] 137 mM NaCl/2.7 mM KCl/10 mM Na2HPO4.7H2O/1.8 mM KH2PO4; pH 7.0) for 1 min. The livers were microdissected and immersed in 0.5% bleach for 2 min, and then rinsed thrice with the LR medium. The tissues were washed with a red blood cell lysing buffer (1 mL; 8.3 g/L ammonium chloride in 0.01 M Tris-HCl buffer) for 1 min, after washing (thrice) with PBS (Ca2+/Mg2+free; 1 mL).
Then, the collected tissue was placed in the LR medium containing 10% antibiotic and antimycotic for 20 min. The tissues were minced and spun at 500 g for 5 min at 4°C. The minced tissues were digested with 0.3% trypsin (in PBS-Ca2+/Mg2+free) for 5 min at room temperature in a 15 mL conical tube. The tube was slowly shaken several times to facilitate cell dissociation. The dissociated cells with trypsin were transferred to a tube containing the LR medium (with fetal bovine serum). This process with trypsin was repeated thrice. The total dissociated minceates were spun at 500 g for 5 min at 4°C. The supernatant was removed and cells were resuspended in the LR medium. Five D. rerio livers were pooled to initiate the culture of primary hepatocytes. The final viability is around 80%. The cells were then plated into 24-well plates (5 × 105 cells/well—for comet assay) or 96-well plates (5 × 104 cells/well—for MTT assay), maintained at 28°C, and treated the following day.
Cells treatment
Cells were washed with PBS (with Ca2+/Mg2+; pH 7.0 [1 mM CaCl2-2H2O/0.5 mM MgCl2-6H2O]) and incubated for 10 min in cold PBS (with Ca2+/Mg2+) with 20 μM of H2O2. H2O2 was used to generate mainly SSB and thus activate the BER system.21–23 Moreover, the alkaline comet assay method has extensively been used to detect SSB induced by H2O2 exposure in different biological models.24,25 Considering preliminary experiments, we identify the 20 μM as the ideal concentration for the analysis of DNA repair activity. We observe that low concentrations of H2O2 (1 μM of H2O2; data not shown) did not induce significant amount of DNA damage to allow the DNA repair activity. Moreover, higher concentrations of hydrogen peroxide (100 μM of H2O2, data not shown) induced a loss of cellular viability, which could interfere with result interpretation. After incubation with H2O2, cells were washed twice with PBS (with Ca2+/Mg2+) and maintained in the LR medium at 28°C for up to 360 min. The primary hepatocytes were centrifuged at 500 g for 2 min in each treatment process.
Cytotoxicity was measured by the MTT assay. Cells (n = 6) were washed with PBS (Ca2+/Mg2+ free) and incubated for 3 h at 28°C with MTT (0.5 mg/mL in LR medium). After the incubation period, the medium was removed and formazan crystals were dissolved in 200 μL of dimethyl sulfoxide (DMSO; Sigma) with gentle shaking. Absorbance values were determined in a multiwell plate reader at 490 nm (ELX 800 Universal Microplate Reader; Bio-TEK).
Comet assay and kinetic measurements
DNA damage was estimated using alkaline single cell electrophoresis (comet assay), performed as described by Singh et al. 26 and Steinert et al., 27 with some changes. Briefly, after trypsinization, a resuspended cell aliquot (20 μL) of each sample (5 × 105 cells/mL) was mixed with 100 μL of 1% low melting point agarose and added to slides that had been covered with a layer of 1.5% agarose (normal melting point). After solidification of the layer, cells on the slides were lysed with a lysis solution (2.5 M NaCl, 100 mM EDTA, 10 mM Tris, 1% Triton X-100, and 10% DMSO, pH 10) overnight at 4°C. Electrophoresis was then performed for 20 min at 1 V/cm and 280 mA. Finally, the slides were neutralized with 0.4 M Tris buffer (pH 7.5). The comet assay was conducted with four samples per group (n = 4) in several experimental times (0; 10; 20; 40; 60; 180; and 360 min). Right before analysis under fluorescence microscope (Olympus BX50), slides were stained with 30 μL of SyBR Safe (2:10,000; Molecular Probes). Hundred nucleoids were randomly selected and photographed on each slide with a CCD camera (Pro-Series; High Performance) coupled to the image acquisition software. The nucleoids were analyzed by Image J software and DNA damage was represented by tail length (μm). Considering that other comet assay parameters were analyzed (tail moment and% tail DNA—data not shown) and they were correlated to tail length (Pearson correlation coefficient 0.93), we opt to show only tail length in the results section.
Determination of median repair time (RT50: time, in minutes, to repair 50% of DNA damage) and 95% confidence interval were calculated using the trimmed Spearman–Karber method, version 1.5.
Statistical analysis
Data are expressed as the mean ± standard error and were analyzed by one-way analysis of variance and Newman-Keuls multiple range test. Significance level was set at p < 0.05 and the analysis assumptions (normality and variance homogeneity) were previously verified.
Results and Discussion
H2O2 cytotoxicity and DNA damage levels
While cell viability was not decreased, 20 μM H2O2 increased the levels of DNA damage in both ZF-L cell line and primary hepatocytes (Table 1). Initial DNA damage induced by exposure to H2O2 was 75.3 ± 12.3 in the ZF-L cell line and 55.0 ± 8.3 in primary hepatocytes (Table 1).
Data correspond to the mean ± SEM and analyzed by one-way analysis of variance.
Different letters indicate statistical differences (P < 0.05).
ZF-L, zebrafish Liver.
BER kinetics
BER occurs in both cell models evaluated (Fig. 1). Ten minutes after H2O2 treatment, 16% and 50% of the initial damage were repaired in the ZF-L cell line and primary hepatocytes, respectively. The ZF-L cell line and primary hepatocytes exhibited an efficient repair capacity with a total recovery within 180–40 min, respectively. The images analyzed in this study clearly demonstrate a reduction in tail length, indicating a reduction of damage over time and the method efficiency (Fig. 2).
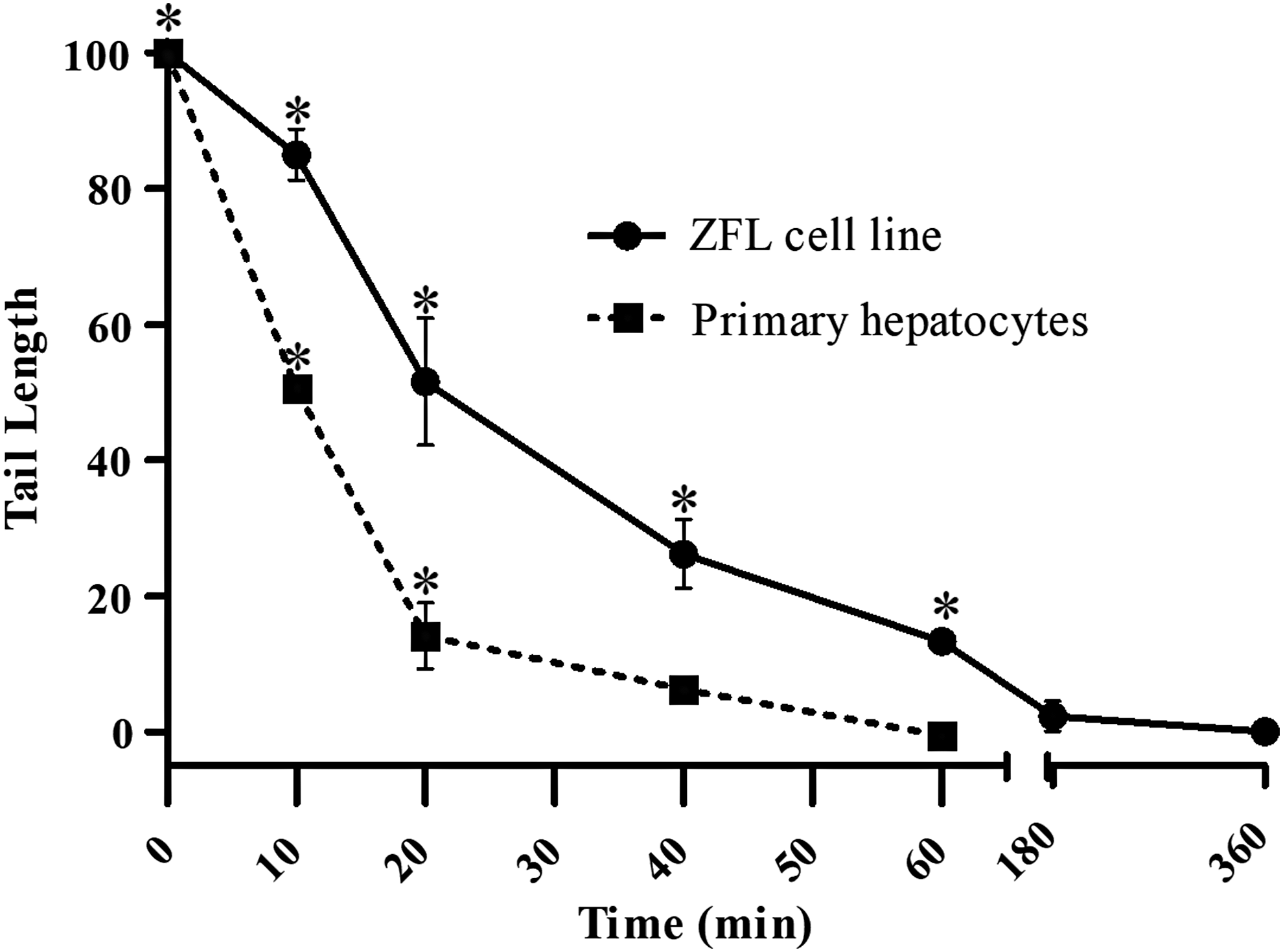
Repair kinetics after exposure to H2O2 (20 μM, 10 min), residual damage level detected by the alkaline comet assay in ZF-L cell line (●) and Primary culture (■). Results are mean ± SEM and *significant difference between exposed and control cells (p < 0.05) for each experimental time point. ZF-L, zebrafish Liver.

Representative images of comet assay in each experimental group. ZF-L cell line and primary hepatocytes 0; 10; 20; 40; 60; 180; and 360 min after treatment with H2O2 (20 μM for 10 min).
Our findings are in agreement with the literature on the capacity of cellular models to repair DNA oxidative damage and SSB. Other fish cellular models, for example RTL-W1 and RTG-W1 (rainbow trout cell lines derived from liver and gill epithelium), showed a complete recovery, with DNA damage levels similar to control within 60–120 min after exposure to 10 μm H2O2. 25 In mammalian cells, such as HeLa cells and human lymphocytes, the SSB repair occurred in 120 min after 100 μM H2O2 treatment. 28 In addition, HeLa and Caco-2 cells have the damages completely repaired within 1 h after a 5-min exposure to 30 and 20 μM H2O2, respectively. 29 All these data indicate that the BER kinetics, after H2O2 treatment, found in this study for ZF-L cell line and primary hepatocytes is similar to that observed in other fish and in human cell lines.
Comparing RT50 values, primary hepatocytes repaired 50% of damage twice as fast as the ZF-L cell line (Table 2). This indicates that the ZF-L cell line has the DNA repair mechanisms altered when compared with systems closer to in vivo. Similarly, alterations in DNA repair mechanisms have already been observed in a study comparing the expression and sensitivity of genes to exogenous ligands in the ZF-L cell line and primary hepatocytes. 30 Analysis of proteomic phenotypes revealed that the Hepa1-6 (Hepatoma) mice cell lines were deficient in mitochondria, leading to rearrangement of metabolic pathways when compared to mice primary hepatocytes. 31 Other studies also show that, serial passaging of cell lines over time may cause genotypic and phenotypic alteration. Moreover, a review about cell line quality summarized that, despite great scientific contributions from investigators using continuous cell lines as research tools, 18–36% of cell lines are misidentified or cross-contaminated. 17
The values were based on Comet assay (Tail Length) and determined using Trimmed Spearman-Karber method, version 1.5.
95% confidence interval.
Understanding the DNA repair in fish cellular models is crucial for the application of these models in ecotoxicological and clinical studies. So far, most studies have involved quantifying the expression of DNA repair genes. Few studies have actually compared the activity of DNA repair between models and over time. In this regard, the comet assay has been considered to be a simple and effective method to measure DNA damage and repair.1,32,33
In conclusion, both ZF-L cell line and primary hepatocytes represent a useful tool for ecotoxicological studies, in regard of the evaluation of DNA damage and repair. The BER system occurs in the two cell lines of the D. rerio liver. This is the first work to describe the DNA repair kinetics in established and primary cells of zebrafish. We observed differences in the time needed for BER between these two in vitro models, DNA repair being more efficient in primary hepatocytes. In addition, the different DNA repair time we found stresses the necessity of precautions other than using similar and identified passage numbers to ensure better and reproducible results when using cell lines. The molecular mechanisms underlying this difference in repair time between ZF-L cell line and primary hepatocytes remain to be determined.
Footnotes
Acknowledgments
The authors are grateful to Mariana L.M. Hoff for article suggestions. This work was supported by grants from the Brazilian Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES; fellows to SRC, RRV and MTR).
Disclosure Statement
No competing financial interests exist.