
Abstract
Abstract
Zebrafish larvae were used to further understand the liver toxicity of nux vomica. The histopathology, protein expression, and gene expression were assessed to confirm apoptosis in the liver, and then, profiles of the metabolites in zebrafish were investigated by untargeted metabolomic assessment to understand the potential toxicity mechanism of nux vomica. Histopathological observations showed that nux vomica caused damage to liver cells. Western blot results indicated increased expression of activated caspase3, and the result of real-time polymerase chain reaction showed a significant increase in the expression level of caspase-3, caspase-8, and caspase-9 genes (p < 0.05) compared with the control group. The liver injury from nux vomica was linked to the downregulation of amino acid (e.g., proline and alanine) and fatty acid (e.g., palmitoleic acid) metabolism and upregulation of some other fatty acid (e.g., arachidic acid) and purine (e.g., xanthine and uric acid) metabolism. The hepatotoxicity of nux vomica resulted from metabolic pathway disturbances, including small molecules involved in energy, purine, lipids, and amino acid metabolism.
Introduction
N
Although mammalian toxicity studies have been the gold standard for hepatotoxicity evaluation and the prediction of risk in humans, in vivo tests are relatively expensive and require large numbers of animals, which are subject to increased scrutiny from animal welfare organizations. 4 In vitro assays have been amenable to high-throughput screening, but unexpected difficulties in correlating known mechanisms of toxicity, and the lack of consensus about the approaches and study designs, have been the main obstacle in mirroring the metabolism of whole animals and in accurately modeling the multicellular processes.5,6
Due to the limitations mentioned above, researchers have shown increased interest in zebrafish-based assays to assess the safety of drugs included in Traditional Chinese Medicine (TCM). 7 The last few years have witnessed an increased number of studies describing the potential use of zebrafish in toxicity studies. The reasons for this trend include not only the advantages of high fertility, reasonable maintenance costs, and rapid extrauterine development 8 but also include the similarity of organs, tissues, and cell types between zebrafish and mammals, promoting its use as a desirable tool in investigating the different toxicological endpoints of the liver. The transparency of the zebrafish embryo 9 and transgenic technology has facilitated a clear time-lapse noninvasive observation of the organ phenotype in complex biological processes 10 and human disease processes.11,12 Furthermore, the visual scoring of phenotypic defects has been realized13,14with the characteristic of high throughout. During this interval, a more modest increase in the publications on zebrafish-based toxicology studies has been underway.15–17
It is worth mentioning that there were a number of early indicators of liver toxicity in zebrafish 18 (McGrath and Li, 2008). Several long-standing methods for liver toxicity screening, such as in inflammatory bowel disease, nonalcoholic fatty liver disease, and alcoholic liver disease, can be modeled in zebrafish.19–22 Zebrafish larvae before 5 days postfertilization (dpf) were considered to not suffer from real pain during toxicity exposure and thus not subject to the strict regulatory requirements and adult mammal principles. Systems toxicology approaches have become a powerful tool for clarifying pathogenesis, identifying biomarkers for diseases23,24 and developing predictive toxicity25,26 through comprehensive information on the biochemistry of pathological conditions, physiological processes, and quantification of a broad spectrum of all endogenous metabolites.27–31
In our previous study, the liver toxicity of nux vomica was demonstrated in the zebrafish model, 32 the results of which provided the initial toxicity phenotype and cell apoptosis in the liver using the acridine orange staining method. Therefore, to identify the underlying toxicity mechanism in this study, histological sectioning and hematoxylin/eosin staining were performed to confirm liver injury in zebrafish liver. Real-time polymerase chain reaction (qt-PCR) was used to assess the expression level of apoptotic regulating genes, including caspase3, caspase8, and caspase9, and Western blotting was used to assess the expression of activated caspase3 protein. Finally, metabonomic technology was used to investigate the effect of nux vomica on endogenous metabolites in zebrafish to provide insights into the herb's toxicity mechanisms. The results of this investigation may provide a scientific explanation for the hepatic toxicity mechanism of this herb and be of great significance in the prediction of liver toxicity in other TCMs in the future.
Materials and Methods
Zebrafish maintenance and embryo collection
Wild-type (AB strain) zebrafish were maintained at 28.5°C in a 14:10-h light/dark cycle in a small aquaculture facility (ESEN, Beijing, China) for aquatic organisms. The fish were fed brine shrimp three times daily. The zebrafish embryos were obtained from spawning adults with a sex ratio of 1:1 (5–8 months old) and raised at 28.5°C in the embryos' water (0.137 mol/L NaCl, 5.4 mmol/L KCl, 0.25 mmol/L Na2HPO4, 0.44 mmol/L K2HPO4, 1.3 mmol/L CaCl2, 1.0 mmol/L MgSO4, 4.2 mmol/L NaHCO3). Zebrafish embryos can receive nourishment from an attached yolk sac, and no feeding was required before 7 dpf. At 4 h postfertilization, the healthy zebrafish larvae were selected through a dissecting microscope, Zeiss Axio Zoom V16 stereo fluorescence microscope (Zeiss, Germany) for subsequent experiments.
Plant material and extraction
The nux vomica (code#160611) was supplied blinded by the Anguo Shengshan Pharmaceutical Co., Ltd. (Anguo, China). After being smashed into 25.4 mm powder, the nux vomica (50.0 g) was extracted with distilled water (w/v 1:10) by refluxing for 2 h. After the crude extract was filtered, the supernatant was collected for further procedures, and the residue was successively extracted with distilled water (w/v 1:10) by refluxing twice, once every hour. The water extract was mixed, filtered, and centrifuged (3000g) for 30 min (RJ-LDL-50G, Wuxi Ruijiang Analysis Instrument Co., Ltd., Wuxi, China) and the supernatant was concentrated under reduced pressure using a vacuum rotary evaporator (Beijing Oriental Science and Technology Development Co, Ltd., Beijing, China). After this, the concentrated solution was collected and dried under a vacuum at room temperature. The dried extract was kept in a desiccator until it was used.
Embryonic exposure experiments and sample collections
All the zebrafish used in this study were utilized in accordance with the Animal Management Rules of the Ministry of Science and Technology of the People's Republic of China for experimental care and use of animals and approved by the Animal Ethics Committee of Beijing University of TCM. To assess the liver toxicity of nux vomica, 4 dpf zebrafish were exposed to different concentrations of nux vomica for 24 h. The nux vomica extract was first dissolved in the embryos' water to prepare the stock solution (400 μg/mL) and then diluted to the required experimental concentration. According to the previous work, the concentration–response curves for nux vomica were Y = 0.74048X-39.1667 and the concentration for the LC10 was 76.523 μg/mL. Thus, the final exposure concentrations were below 70 μg/mL to avoid the appearance of death and to detect the major toxicity targets. For each concentration, 80 embryos were cultured in 12-well plates. Each well contained 4 mL of test medium and 20 zebrafish. Three replicates were conducted for each group. At the experimental exposure endpoint, the samples were prepared according to the relevant requirements for the follow-up experiments.
Histopathological analysis
According to the publication 33 (Amali et al., 2006), at the end of treatment, 20 whole zebrafish from different groups were fixed in 4% paraformaldehyde until the mounting in 1% agarose and processed sequentially in ethanol and xylene. The agarose block was gradually dehydrated with 50%, 70%, 85%, 95%, and 100% ethanol and 100% xylene. Then, the tissues were sectioned (4–5 μm) after being embedded in paraffin wax and mounted on slides. Finally, the sections, stained with eosin and hematoxylin, were air-dried and observed in an Olympus BX51 microscope. The images were captured at 400 × magnification using an Olympus DP71 camera and software. The experiment was repeated three times.
Western blotting analysis
The Western blotting analysis was according to the published research. 34 35 Whole zebrafish (n = 80 per concentration) were collected and homogenized in RIPA lysis buffer (Beijing TDY Biotech CO., Ltd. Beijing, WB0003) (w/v = 1:9) containing protease inhibitors (1 mM PMSF). The whole homogenization process had to be carried out in an ice/water bath. The homogenate was incubated in ice for 20 min and centrifuged (4°C, 13,000 rpm) for 20 min to get the supernatant. Protein quantification was determined by the BCA method (Beijing TDY Biotech Co., Ltd. Beijing, WB0028). The final concentration of each sample was 2.5 mg/mL and it was boiled for 5 min to realize protein denaturation. The samples and β-actin were normalized for equal loading (20 μg/lane) and electrophoresed on 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis gels. Immediately after electrophoresis, the gels were transferred to a nitrocellulose filter membrane. Blocking of membranes was performed in 3% bovine serum albumin-Tris buffered saline Tween (BSA-TBST) solution for 1 h at room temperature. The membrane was incubated in rabbit anti-caspase3 (ab13847, abcam) at a dilution of 1:500 with 3% BSA-TBST (4°C) overnight and washed in TBST five times before the next incubation, each time for 3 min. Secondary antibody incubation was performed for 40 min with goat anti-rabbit secondary antibodies IgG(H+L) HRP (Beijing TDY Biotech CO., Ltd. Beijing, S004) at 1:3000 dilution with 5% nonfat milk-TBST. After this, the membrane was washed in TBST six times, each time for 3 min. Finally, the protein bands were visualized using a ChemiDoc XRS (Bio-Rad, Marnes-la-Coquette, France) using the ECL Kit (Millipore, WBKLS0500). Protein levels were then determined by computer-assisted densitometric analysis (Densitometer, GS-800, Bio-Rad Quantity One). Protein expression density values from the different treated groups were normalized as changes relative to the expression levels of the control. Each test was performed in three experiments with different batches of zebrafish to ensure consistency of results.
Total RNA extraction and reverse transcription-polymerase chain reaction
The larvae were washed twice with the embryos' water and the total RNA of the 80 homogenized zebrafish larvae from the different groups was extracted using the TaKaRa Mini BEST Universal RNA Extraction Kit (Takara, Tokyo, Japan) according to the manufacturer's instructions. The total RNA content was determined by measuring the absorbance at 260 nm, and the quality was verified by measuring the 260/280 nm ratio using a spectrophotometer (NanoDrop 2000; Thermo Scientific). Equal amounts of total RNA samples from the different groups were reverse transcribed to cDNA using the FastKing RT Kit (Tiangen Biotech Co, Ltd., Beijing, China). The qt-PCR was carried out using the CFX96 Touch Systems (Bio-Rad) and TransStart Top Green qPCR Super Mix (TransGen Biotech, Beijing, China) in 96-well PCR plates (Applied Biosystems, Foster City, CA). All processes were according to the manufacturer's instructions provided by the different kits. The amplification cycling protocol was as follows: 95°C for 3 min, followed by 40 cycles of 60°C for 20 s, 60°C, 9°C for 15 s, 6°C for 10 s, and 9°C for 15 s. All data were statistically analyzed as fold change between the exposed groups and control. The experiment was conducted in triplicate. The caspase-3, caspase-8, and caspase-9 primers were obtained from Primer Premier 5.0 software, and the primer sequences are shown in Table 1.
The Sequences of the Primers
Design of the metabolomic analysis
Chemicals and reagents
High-purity methylamine hydrochloride, fatty acid methyl ester (C7–C30, FAME), anhydrous pyridine (99.5%), and anhydrous sodium sulfate were purchased from the Sigma-Aldrich Company (St. Louis, MO). Derivative reagent MSTFA (containing 1% TMCS), methanol (Optima liquid chromatograph mass spectrometer [LC-MS]), and normal hexane were purchased from Thermo Fisher (Fair Lawn, NJ). Dichloromethane (99.5%), chloroform (99%), and acetone (99.5%) were purchased from the China National Pharmaceutical Group Corporation (Beijing, China). The experiment used ultrapure water (18.2 M) prepared by Reference Ultrapure Water Systems (Billerica, MA) equipped with an exclusive filter head for liquid mass chromatography (LC-MS Pak).
Sample preparation
In this experiment, the collected samples were precisely weighed and a weight normalization method was used to correct the individual sample errors. Sample preparations were processed based on the published literature36,37and some improvements were made. Samples of the zebrafish to be estimated were weighed accurately at 25 mg under the protection of the salt/ice bath, following which, the precooling grinding beads of chromium oxide and 10 μL internal standard water solution were added and mixed slightly. The samples were homogenized in an 80 μL precooling 50% methanol (Thermo Fisher) aqueous solution using a fully automatic tissue homogenizer (Next Advance, Inc., Averill Park, NY). All homogenates were centrifuged for 20 min under conditions of 13,500 rpm and 4°C (Microfuge 22R, Beckman Coulter, Inc., Indianapolis, IN). The second homogenates were centrifuged again under the same conditions after removal of the upper layers and the addition of 200 μL of precooling chloroform/methanol (China National Pharmaceutical Group Corporation, Beijing, China) (3:1, v/v). All supernatants were mixed to obtain the whole supernatant. Then, 200 μL of supernatant in the automatic sample glass bottles (Agilent Technologies, Foster City, CA) was transferred to the vacuum centrifuge concentrator (Labconco, Kansas City, MO) to remove the chloroform, following which, the sample was transferred to the ultralow freeze dryer (Labconco, Kansas City, MO). When its temperature reached room temperature, high-purity nitrogen as the protectant was introduced and the samples were placed in the XploreMET system controlled by the automatic derivation platform (Gerstel GmbH & Co., Mulheim, Germany) to process silane derivation as shown in the following steps: addition of a 50 μL methoxypyridine solution (20 mg/mL) and incubation at 30°C for 2 h and addition of 50 μL MSTFA and incubation at 37.5°C for 1 h. In the derivation process, the derived samples were automatically entered by the injection robotic arm.
Instrumentation
In this experiment, the gas chromatography-time of flight mass spectrometry (GC-TOFMS) (LECO Corp., St. Joseph, MI) was used as the detecting platform for screening of nontargeted metabolomics. The optimized instrument parameters are shown in Table 2. The instrument system had to be optimized and maintained after the derivation and analysis of three batch samples.
Instrument Settings for the GC-TOFMS
GC, gas chromatography; GC-TOFMS, gas chromatography-time of flight mass spectrometry.
Quality control/assurance procedures
In this experiment, a series of standardized measurements were adopted to ensure the data quality of the fully automatic nontargeted metabolomic technology platform.
General requirements for handling precious samples
The defrosting process was carried out slowly under the ice/salt condition to avoid the effect of metabolic enzymes reactivated on the metabolites and concentrations of metabolites. Furthermore, the reagent used in the extraction process of the precipitation protein had to be precooled at −20°C in the refrigerators. Although the ice bath could keep the frozen samples at a low temperature (−10°C to −0°C), the whole sample preparation process needed to be completed as soon as possible. Dichloromethane (China National Pharmaceutical Group Corporation, Beijing, China) was used to remove crystallization water and moisture residing in the freeze-dried samples to avoid degradation or failure of the derived products. High-purity nitrogen (Parker Balston, Lancaster, NY) was added to the automatic sample bottle to remove any possible residual dichloromethane and ensure the oxygen-free state of the system. The auto-derivatization process was conducted by the automatic crown capper (Agilent Technologies, Foster City, CA). The sample injection had to be completed again within 24 h once the injection problem was found by the automatic software check. Preparation of the unqualified samples had to be conducted again within 36 h for the subsequent analysis tests.
Automated sample processing and extraction procedures
Automatic processing techniques, including the automated liquid workstation (epMotion M5073; Eppendorf, Hamburg, Germany) or repeat sampler (Eppendorf), were used to ensure repeatability, operability, and accuracy of test results. The high-throughput freeze-drying process could handle 1000 samples each time, which guaranteed stability of samples and eliminated errors caused by the preparation of various batches. The automated derivatization and injection processes were conducted by the automatic preparation equipment (Gerstel, Mulheim, Germany), which was controlled by the XploreMET systems. The injection was processed after 6 h of automated derivatization; thus, the waiting time for the sample was greatly shortened through automated derivatization and injection. A low-temperature sampling plate was used to avoid degradation of the sample in the sample injector and to enhance reproducibility of the derivative process, effectively avoiding systematic errors.
Quality controls
The Metabo-Profile XploreMET systems with strict requirements for quality control were adopted in the nontargeted metabolomic platform. Five different internal standards were required for quality control in the sample preparation and analysis processes, including a standard mixture, retention index, internal standard, mixture quality controls, and NIST standard serum SRM 1950 (Gaithersburg, MD). In addition, a reagent blank, system blank, deactivation of the glass liner of the injection port, and biological samples for system balance were also used to monitor for possible contamination in the analysis process and guarantee the reliability and stability of data quality. To eliminate any errors resulting from the sequence of the analytic process, the samples to be measured were all intersected and injected according to proportionality rather than group information.
Metabolite annotation
For the sake of accuracy of endogenous metabolites, high-purity chemical standards were purchased from Sigma-Aldrich, Cayman Chemical, Santa Cruz Biotechnology, and Advantis. Based on these standards, a standard database (JiaLib) of endogenous metabolites associated with the silylation derivatization was established for the GC-MS, which contained more than 1500 kinds of endogenous metabolites. The XploreMET system software and database were used to automatically compare the retention index and mass fragmentograph after deconvolution.
Data processing for the raw mass spectral data
The raw data produced by the GC-TOFMS were automatically output to the XploreMET software through ChromaTOF software (v4.51.6.0, LECO, CA), where the baseline correction, deconvolution, extraction, and alignment of the raw chromatographic peak signal, retention index correction, identification of metabolites, and data preprocessing (normalization and standardization) were carried out. In addition to accurate identification of a single standard, the metabolites with biologically useful functions in the upstream and downstream were also extracted from the Kyoto Encyclopedia of Genes and Genomes (KEGG) and the Human Metabolome Database (HMDB). Furthermore, biological significances were expressed by the ratio of products to reactants, which indirectly reflected the changes in metabolic enzymes in the upstream and downstream, and were a useful supplement to a single metabolite biomarker at the metabolic level.
Metabolic pathway enrichment analysis
For peaks with VIP >1 and p < 0.1, the metabolites were considered to be the potential differential metabolites to conduct the enrichment of the signaling pathways. A hypergeometric algorithm was used for the enrichment of the metabolic pathways of the KEGG metabolites, to strengthen the contribution of metabolites and find the key metabolic network.
Statistics
A variety of multidimensional statistical methods, including principal component analysis (PCA), partial least squares-discriminant analysis (PLS-DA), orthogonal partial least-squares discriminant analysis (OPLS-DA), random forest, and support vector machine, were used to explain the relationships between variables. In this experiment, a variety of single-dimensional statistical methods, including the Student's t-test, Mann–Whitney–Wilcoxon (U-test), and analysis of variance (ANOVA), were used in conjunction to screen the overlapping disparate metabolites, which were generally considered to be stable differential metabolites. *p < 0.05 and **p < 0.01 were considered significant.
Results and Discussion
Hepatic histopathology
Zebrafish livers in the acute nux vomica-treated group exhibited histopathological changes compared with control (Fig. 1D). The zebrafish exposed to nux vomica elicited obvious swelling of hepatic cells, obscure cell borders, and hepatocellular vacuolar degeneration in the 30 μg/mL group (Fig. 1A). Hepatic lesions became much more severe in the zebrafish exposed to 50 μg/mL nux vomica (Fig. 1B). The cellular structure was nearly collapsed in the livers of zebrafish receiving the 70 μg/mL nux vomica (Fig. 1C).
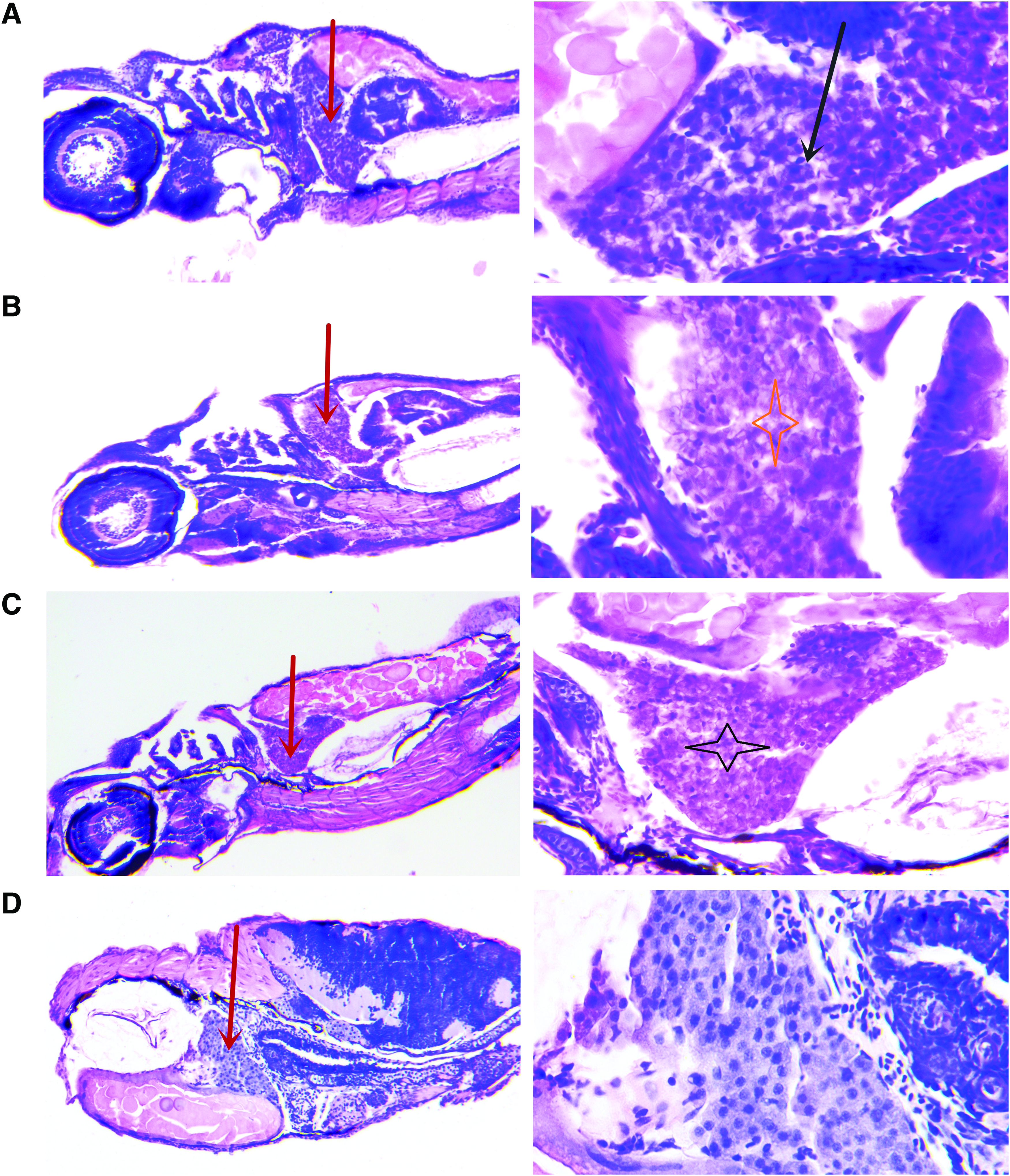
Histopathology of zebrafish liver after acute exposure to nux vomica for 24 h. Photomicrographs of liver sections (4–5 μm) stained with hematoxylin and eosin.
Nux vomica induced gene and protein expression associated with apoptosis in liver
In the previous experiment, apoptosis in the liver was observed. It has been demonstrated that the caspase family plays an important role in the regulation of apoptosis and this is varied among different positions in a zebrafish. First, the levels of activated caspase3 were analyzed by Western blot in zebrafish from the different groups compared with the control group. We found higher levels of activated caspase3 in the treated group than in the untreated group (Fig. 2) with a significant difference (p < 0.01). Therefore, we examined whether nux vomica induces apoptosis via the caspase pathway by the examination of the gene expression of caspase3, caspase8, and caspase9. The result showed that, compared with the control group, the gene expressions of caspase3, caspase8, and caspase-9 (Fig. 3) were significantly upregulated in a concentration-dependent manner with a significant difference.
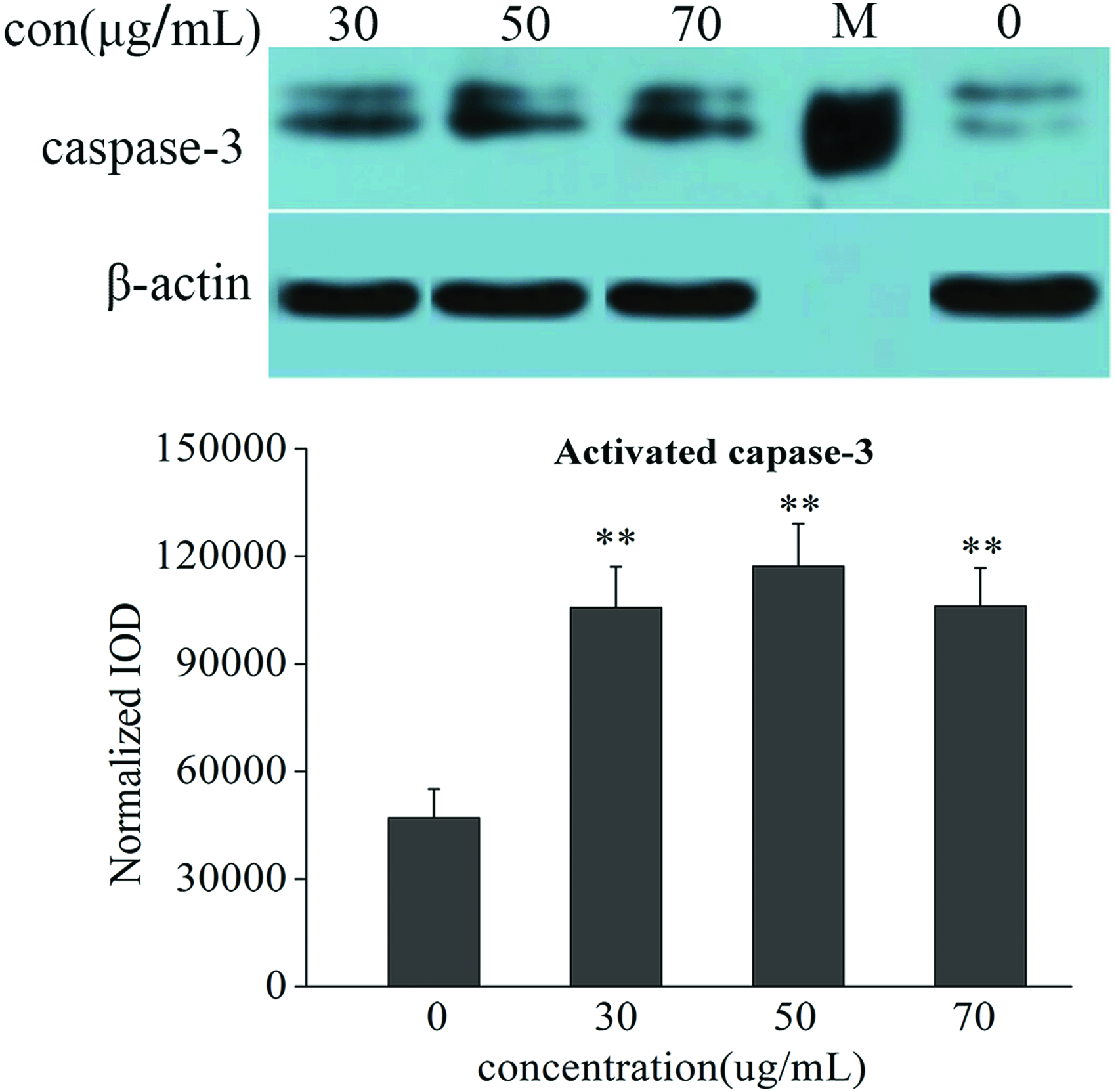
Protein expression of caspase3 was examined by Western blot in nux vomica. The treated group and control group zebrafish larvae n = 80. Values are averages ± SDs, **p < 0.01 versus control, by one-way ANOVA. Western blot of zebrafish treated with increasing nux vomica. Concentrations showed a dose-dependent increase of caspase3 in the treated groups, indicating real apoptosis. ANOVA, analysis of variance; con, concentration; SD, standard deviation. Color images available online at www.liebertpub.com/zeb
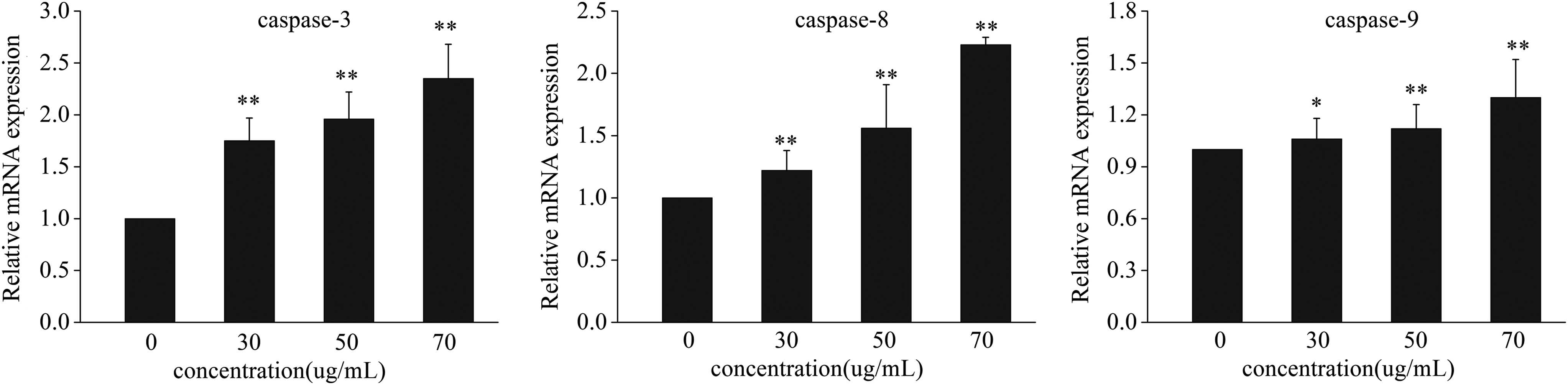
Expression profiles of the tested genes in the control group and the groups exposed to 0, 30, 50, and 70 μg/mL nux vomica for 24 h. Values that were significantly different from the control are indicated by asterisks (one-way ANOVA, *p < 0.05; **p < 0.01 vs. control). The values are the means of three determinations on each of the three replicate exposures, and are presented as the mean ± SD. The results of zebrafish treated with increasing nux vomica concentrations showed a dose-dependent increase of caspase-3, caspase-8, and caspase-9 expression levels compared with the control group.
Metabolite profile visualization
A control chart (Fig. 4) was used to monitor the error in the analysis process and find the outliers whose scores exceeded the ±3 standard deviation. The results showed that all samples met the requirements of sample quality control and no abnormal metabolic spectrum was found.
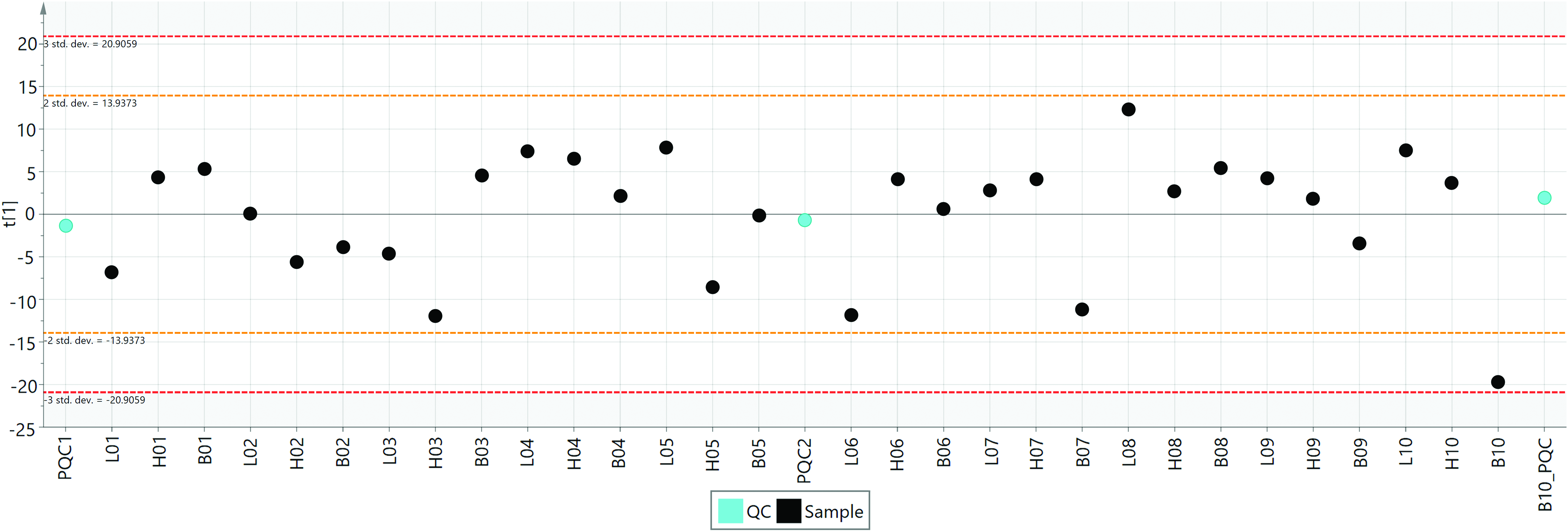
QC/QA control chart with the PC1 among the samples. Each dot represents global metabolic profiles from an individual. The black dots and blue dots were samples and mass control, respectively. The scores that exceeded ±3SD were generally considered to be at risk for outliers. QA, quality assurance; QC, quality control. Color images available online at www.liebertpub.com/zeb
The results of identification of the metabolites revealed that accurate qualitative analysis of 123 endogenous metabolites was achieved, including amino acids and derivatives, carboxylic acids and derivatives, fatty acids and conjugates, hydroxy acid and derivatives, monosaccharides, imidazopyridines, and others (Fig. 5). Based on the metabolic pathway and regulatory network from the KEGG and HMDB databases, 36 metabolite ratios (metabolic enzymes) were re-extracted, while 245 metabolites were still unknown.
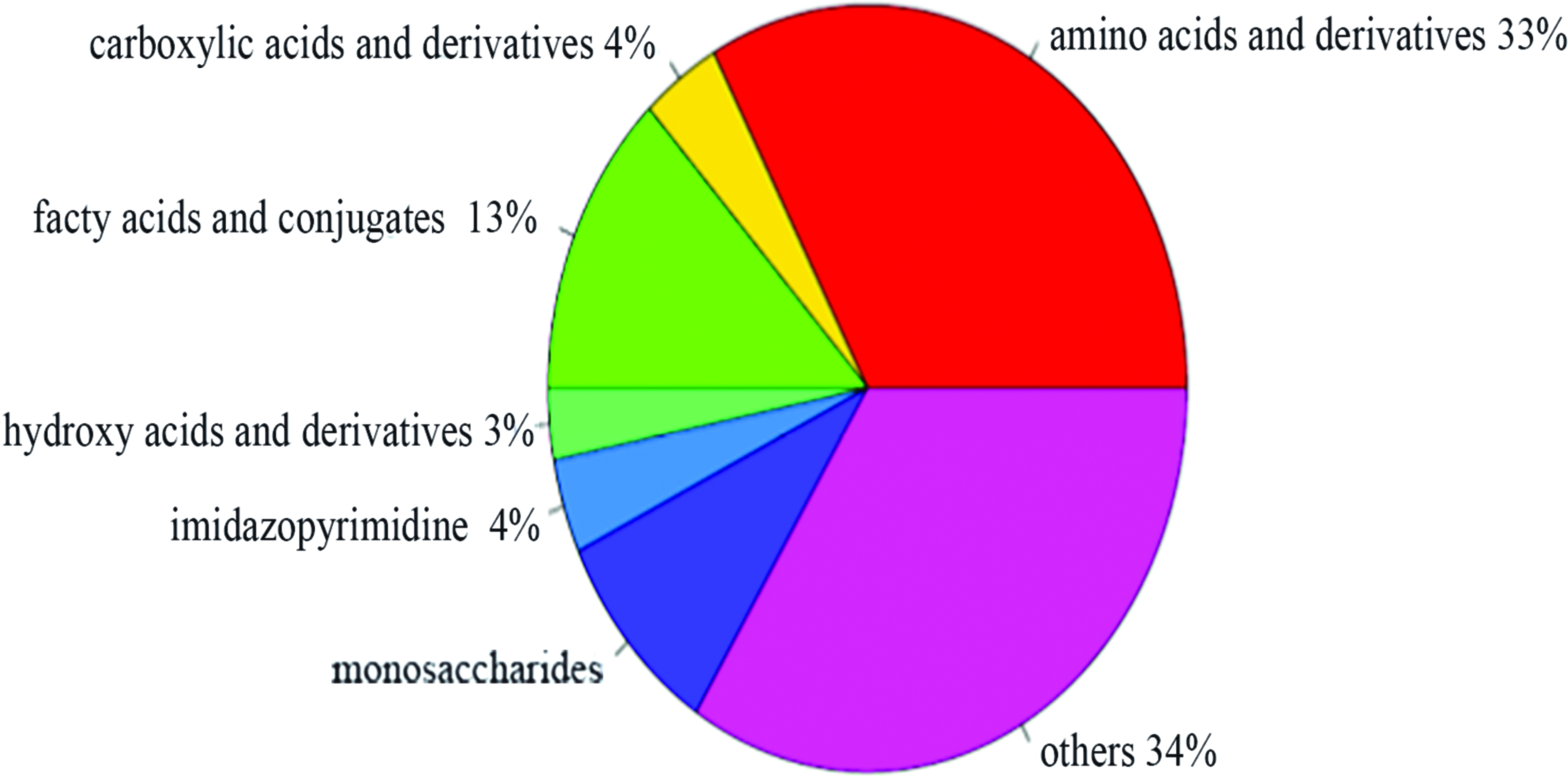
Classification of different metabolites in zebrafish exposed to nux vomica. Color images available online at www.liebertpub.com/zeb
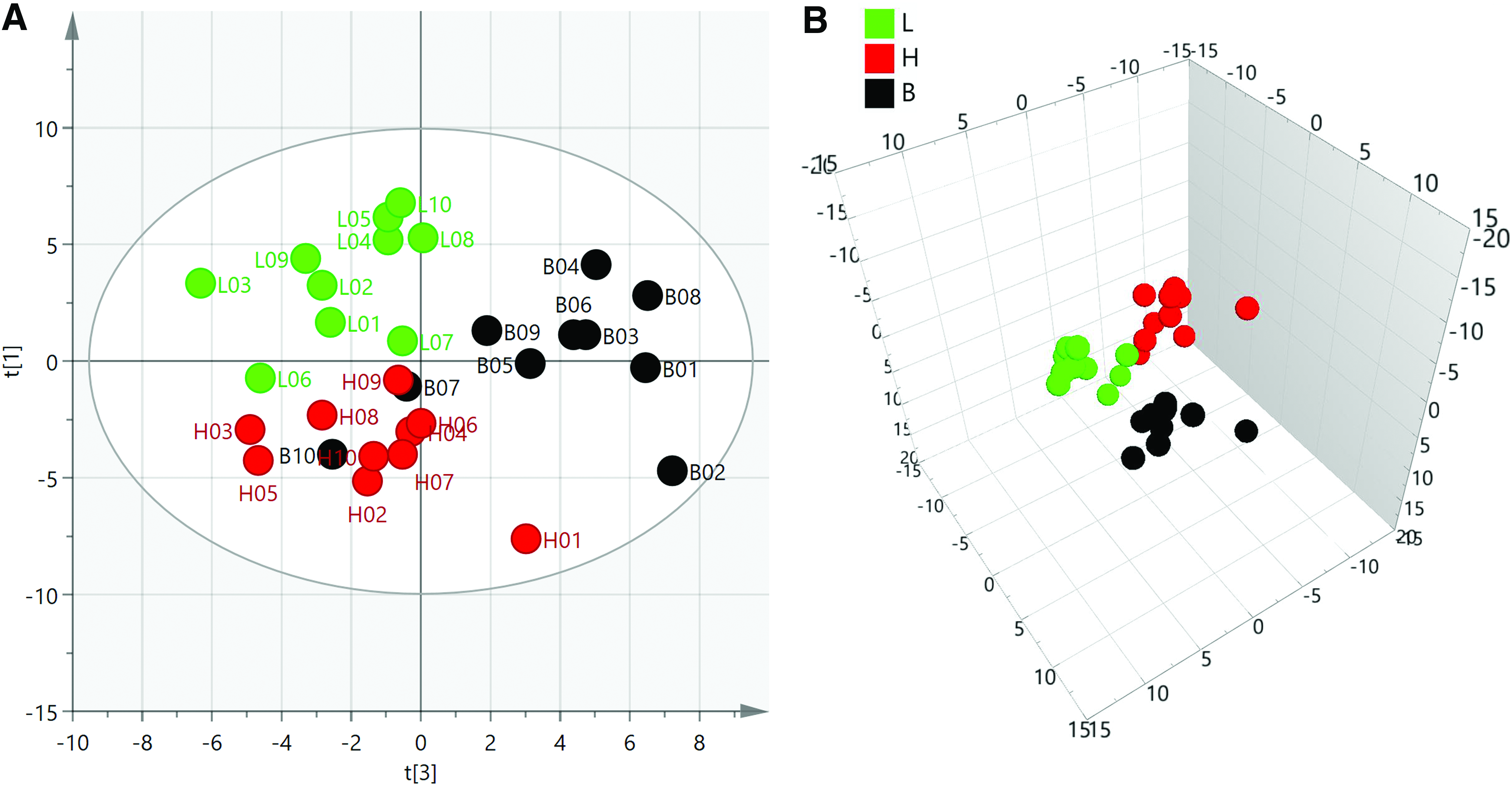
The results of the PCA are shown in Figure 6, from which some conclusions can be drawn: (1) All samples were distributed within the 95% confidence interval of Hotelling's T2, indicating that the biological and analytical results of the whole batch of samples were stable. (2) The separation trend between the control group (Class 1;B) and the high-concentration group (Class 3;H) was significantly more obvious than that between the control group and the low-dose group (Class 2; L). (3)The separation trend between the H group and the L group also existed. Some samples (L07) from the L group were sensitive to the treated condition.

The PCA score of all the samples. The PCA plot
The result of cluster analysis (Fig. 7A) and correlation analysis (Fig. 7B) showed a complex metabolic spectrum among the three groups of samples with a significant difference. All identified metabolites were also classified into their respective groups according to the correlation analysis.
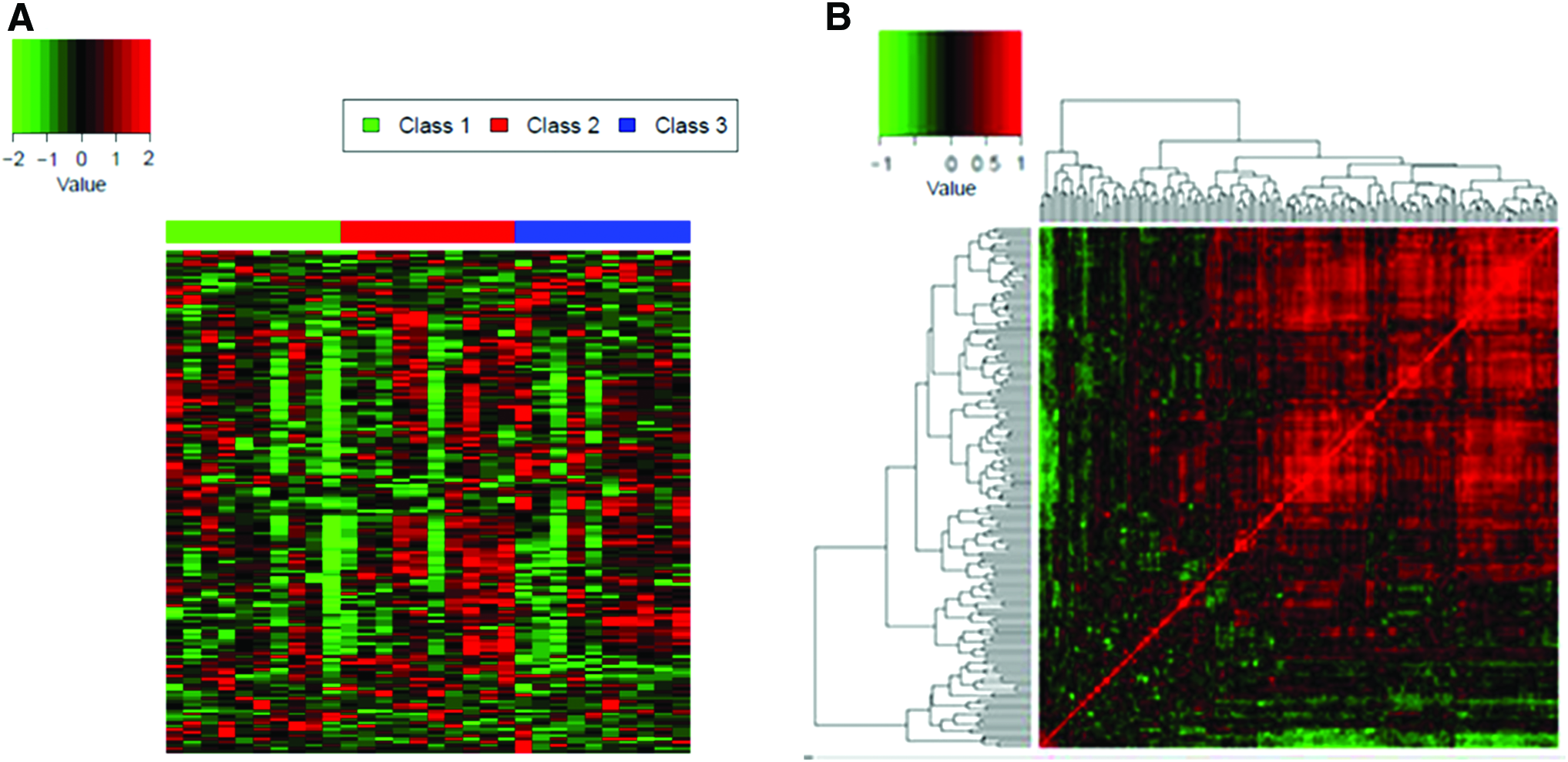
The z-score
Furthermore, the supervised PLS-DA method was used for clustering result visualization among the three groups given the complexity of the biological samples and the individual differences. The clustering results of the metabolic profiling displayed a better regularity among the three differently treated groups (Fig. 8).
Two-dimensional PLS-DA score plot
Pairwise comparisons
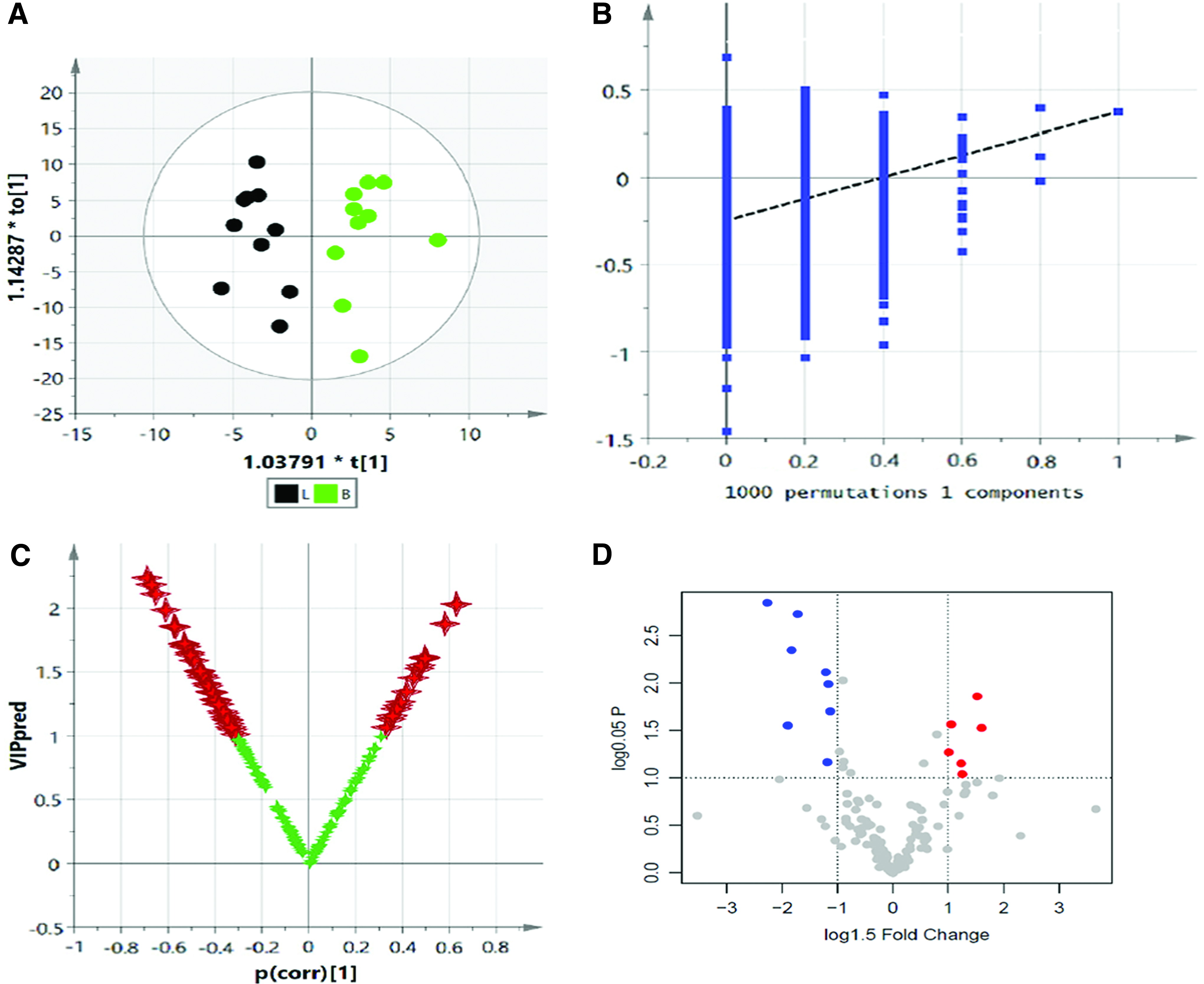
OPLS-DA was used to distinguish the difference in the metabolic spectrum between the B and H, and significant differences in the metabolic spectrum (CV-ANOVA, F = 4.36, p = 0.013) (Fig. 9A) were observed through the one predictive component and two orthogonal components (Fig. 9A, R2Y = 0.98, Q2Y = 0.67). Then, the reliability of this model was further identified by testing 100 random samples (Fig. 9B, Q2 = −0.39). OPLS-DA was also used to find the key metabolites and their metabolic regulation network associated with the differences in metabolic phenotype, and the result of the Vplot showed that there was a significant difference in 53 metabolites and their metabolic enzymes (Fig. 9C). The result of the normal distribution test showed that ∼50% of the metabolites were in a non-normal distribution in the sample set; thus, the one-dimensional Student's t-test or U-test was suitable for verification of the different variables. In this experiment, the Student's t-test was considered to be the criterion for screening and verification of the different variables (Fig. 9D). For the purposes of visual representation of the differences between the two groups, the boxplot of representative different metabolites in the ranking is shown in Figure 10.
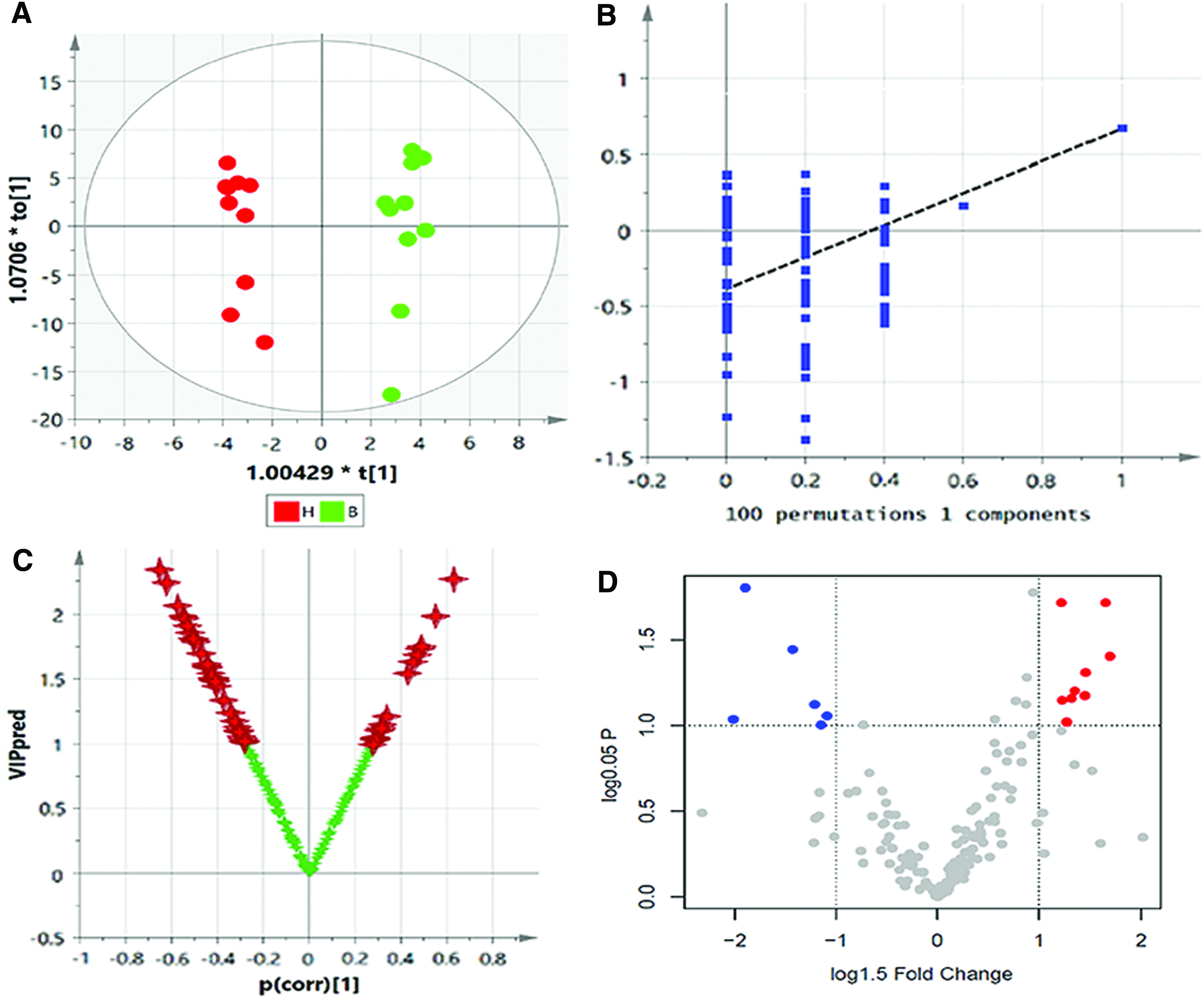
Top-ranked differential metabolites between B and H. Color images available online at www.liebertpub.com/zeb
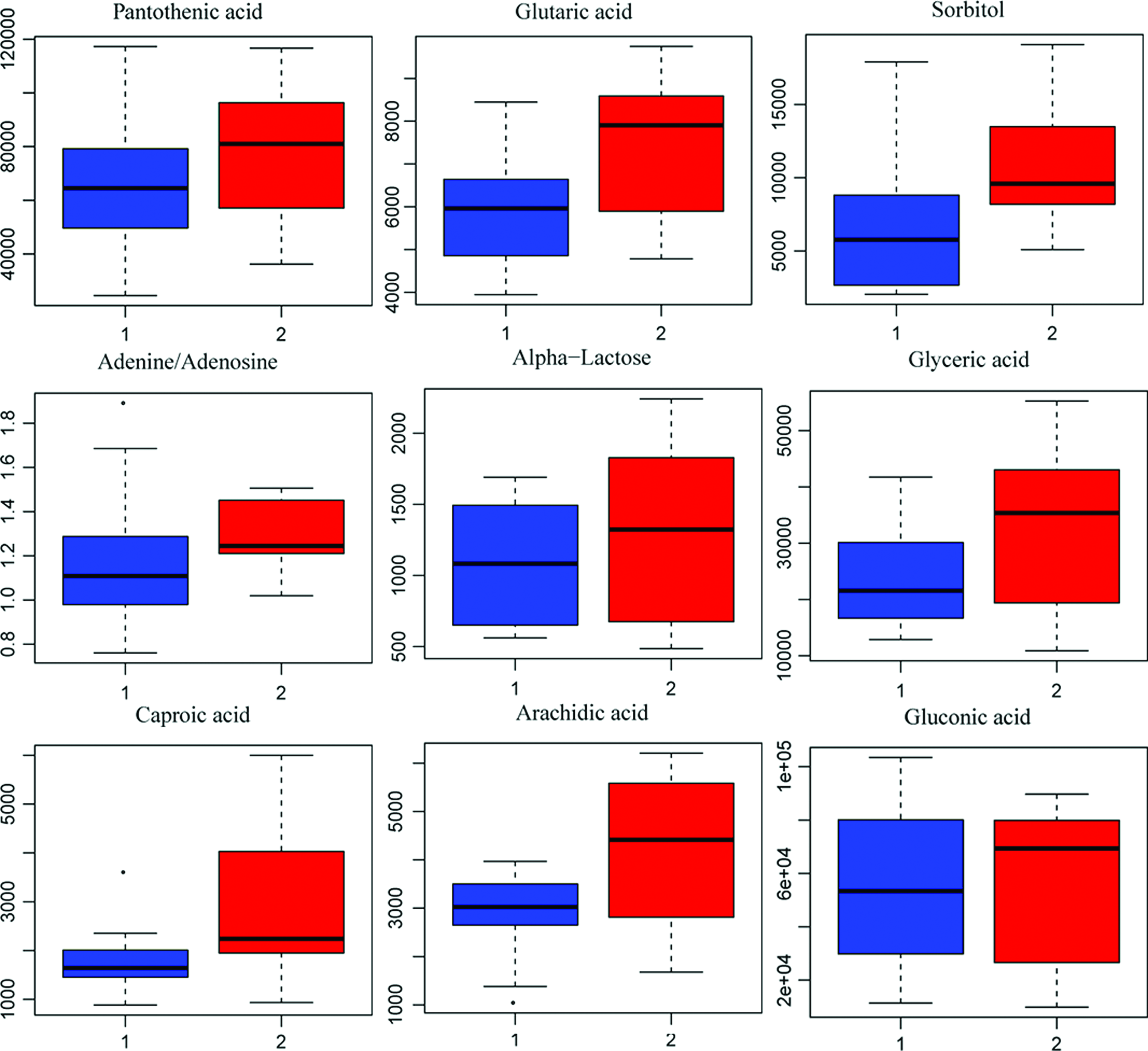
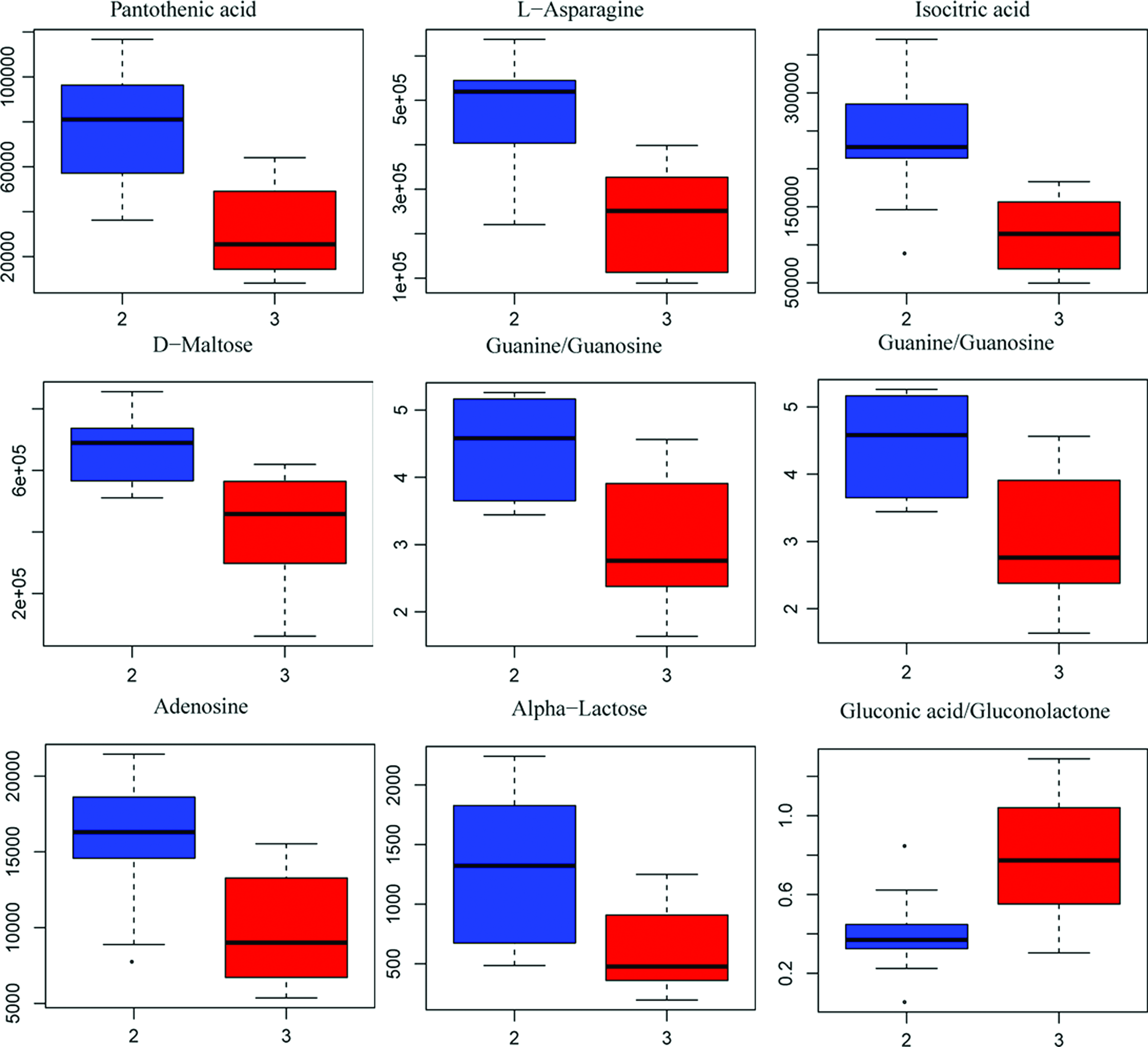
For the B and L groups, the differential trends in metabolite profiling were obviously observed through the one predictive component and one orthogonal component (Fig. 11A, R2Y = 0.84, Q2Y = 0.38), while the result of V-ANOVA showed inconspicuous differences (CV-ANOVA, F = 2.25, p = 0.112) (Fig. 11A). Similarly, the test of 100 random samples was processed and the credibility of the prediction model was verified (Q2 = −0.25, Fig. 11B). The result of the Vplot (Fig. 11C) showed 55 metabolites and their metabolic enzymes, which were associated with the differences in metabolic phenotype. Approximately 50% of the metabolites were in a non-normal distribution and the Student's t-test was considered to be the criterion for screening and verification of different variables (Fig. 11D). The boxplot of the representative different metabolites within the top is shown in Figure 12.
The visualization and Vplot analysis of the overall metabolite profile differences between B and L.
Top-ranked differential metabolites between B and L. Color images available online at www.liebertpub.com/zeb
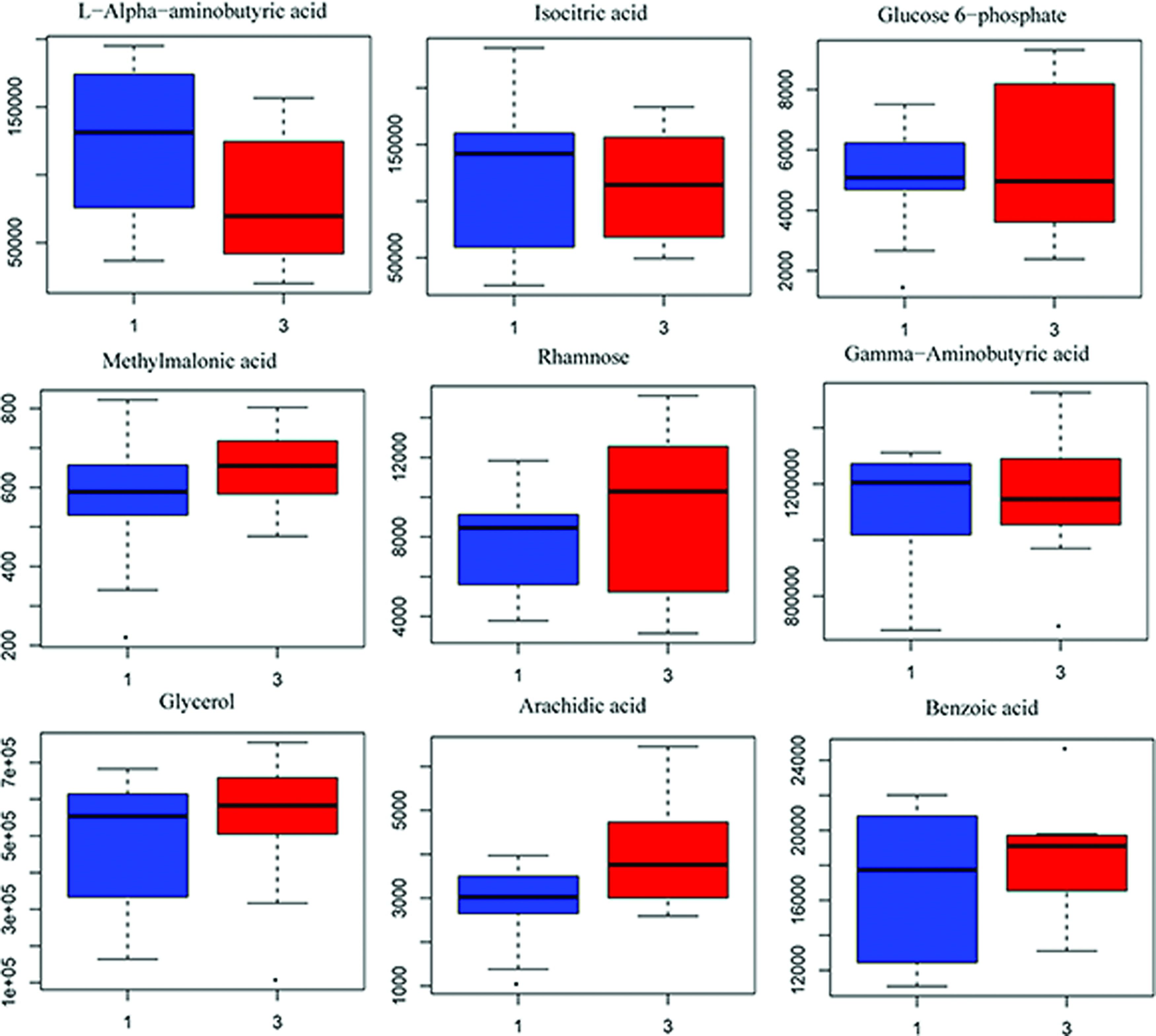
Similarly, the one predictive component and one orthogonal component were used to evaluate the different trends in metabolite profiling (Fig. 13A, R2Y = 0.85, Q2Y = 0.53), and there was a significant difference (CV-ANOVA, F = 4.15, p = 0.018) between the L and H groups. The test of 100 random samples was processed and the credibility of the prediction model was verified (Q2 = −0.25, Fig. 13B). The result of the Vplot (Fig. 13C) showed 54 metabolites and their metabolic enzymes with a significant difference. Approximately 50% of the metabolites were in a non-normal distribution and the Student's t-test was considered desirable as the standard for screening and verification of different variables (Fig. 13D). The boxplot of representative different metabolites within the top ranking is shown in Figure 14.

Top-ranked differential metabolites between L and H. Color images available online at www.liebertpub.com/zeb
Metabolic pathway enrichment analysis
All the differential metabolites among the groups and the metabolic effect with regard to the exposure concentrations and metabolites are summarized (Fig. 15). For example, the proline concentration was decreased 28.24% in the L group and 36% in the H group compared with the B group. The gluconic acid concentration in the L group was increased 8%, while it was increased up to 79% in the H group compared with the B group in the glucose metabolism pathway.

Differential metabolite trend analysis in L and H (vs. B). Color images available online at www.liebertpub.com/zeb
In this study, significantly changed metabolites were used to reveal the major perturbed pathways. The metabolic pathway enrichment analysis (MPEA) results were concluded and differential metabolic regulation networks among the three groups are shown in Figure 16A–C, respectively. As a result, some differential metabolic pathways were considered to be potential target pathways, including the TCA cycle, pyruvate metabolism, glycolysis or gluconeogenesis, and glycerophospholipid metabolism. Those pathways were highly related to amino acid metabolism, lipid metabolism, and energy metabolism.

Metabolic pathway enrichment analysis.
Discussion
Since those articles using a chemical screen with live zebrafish in a 96-well plate 38 and the earlier review of zebrafish toxicity assays were published, 39 zebrafish have received increased attention from researchers in investigating the pathogenesis of human diseases and drug screening. 40 Despite the differences existing in pharmacological effects between zebrafish and humans, there is a high degree of similarity in the gene sequences and protein function 41 maintaining similar elaborate response mechanisms to xenobiotics. Furthermore, great numbers of small molecules with conserved biological activities in zebrafish and humans have been found. Nevertheless, the toxicity evaluation endpoint was readily applied using nondestructive methods, including high-resolution microscopy, and achieved in a high-throughput application with advances in automation and software analysis.
In the previous study, the zebrafish larvae were used to assess the system toxicity of nux vomica through the overall phenotype of different organs, including the liver, and apoptosis in the liver was observed. In fact, apoptosis was an important process for performing self-elimination of superfluous or injured cells, determined by their own built-in programs, which were also involved with cell injury. In this experiment, we performed a systematic review of the toxic effects of nux vomica in zebrafish larvae by combining metabolomics, gene and protein expression, and histopathological approaches to increase our understanding of the mechanism of nux vomica-induced hepatotoxicity.
Histological examinations revealed that the most pronounced morphological alteration was observed, including obvious swelling of hepatic cells, degeneration of hepatocellular vacuoles, hepatic lesions, and destroyed cellular structure. Measurement of the caspase family gene and protein levels had been the most commonly used biomarkers for the assessment of effects of chemicals on apoptosis in the animal model, including zebrafish. Each of the genes associated with cell apoptosis was examined in this experiment. The result of Western blotting and quantitative real time polymerase chain reaction indicated that all genes were consistently upregulated in response to apoptosis resulting from the nux vomica. As Figure 4 shows, the expression levels of caspase3, caspase8, and caspase9 were increased in a concentration-dependent manner supporting this apoptosis model in zebrafish larvae. Besides, these observations were very similar to the previous results with acridine orange. What is worth mentioning is that the apoptosis and lipid accumulation mostly existed in the liver area in that concentration in this experiment, although the nux vomica could lead to the different organ toxicity. Thus, the great changes in gene and protein expression levels were the result of liver injury (data were not provided). The current results suggest that gene expression may be as sensitive as gross morphological endpoints (e.g., liver size, liver function) indicating that different endpoints should be used in the same experiment when evaluating the toxicity of chemicals.
In this study, we analyzed the effect of nux vomica exposure on the metabolomic changes as well as on the metabolomic pathway in zebrafish larvae to verify whether mechanistic information can contribute to the applicability of zebrafish in prescreening of TCM with hepatotoxic potential and to derive predictive markers for hepatotoxicity as the final goal. The result indicated that the pathways associated with apoptosis were affected, but the pathways were different under different conditions.
Global metabolic disorders in response to nux vomica exposure
The peaks in the metabolisms were input to the XploreMET software for further PCA and PLS-DA modeling. In the PCA plot, all quality control samples were clustered tightly among themselves and met the standard of quality control, suggesting good quality in sample processing and reproducibility of GC/MS. However, the spread of samples in both the control and treated groups was observed as indicating high biological variations. The PLS-DA results showed a decent separation of the metabolic profiling between the exposure groups and the control group. However, the result of Pearson's correlation analysis showed that the L group was more sensitive than the H group and different metabolic profiling was obviously observed. Subsequently, the differential metabolites were discovered and 123 differential metabolites were identified in total, including amino acids and derivatives, carboxylic acids and derivatives, fatty acids and conjugates, hydroxy acids and derivatives, imidazopyridines, monosaccharides, and others. Furthermore, the metabolites were individually analyzed to explore the significant differences between the treated groups and control.
Based on pairwise comparison analysis, the multiple tests and analyses of the metabolomics in the different groups showed pronounced effects of nux vomica exposure on purine metabolism, amino acids and amines, carboxylic acid metabolism, fatty acid and lipid metabolism, as well as in the sugars and sugar alcohol metabolism. However, most metabolisms showed a similar trend such as with proline and gluconic acid, regardless of some of the different metabolite trends, indicating that changes in most metabolics were dependent on concentration and that nux vomica exposure had specific toxic effects in zebrafish liver even at a low concentration.
Ingenuity pathway analysis revealed liver damage at the molecular level
To infer the possible biological impact associated with the metabolic changes, KEGG pathway analysis of metabolites (VIP >1, p < 0.1) was conducted for the metabolomic pathway analysis.
The experimental evidence, knowledge-based data mining of biological pathways, and biological interpretations together indicated that the toxicity of nux vomica involved amino acid metabolism, glycogen metabolism, and lipid metabolism. In molecular and cellular functions, amino acid metabolism was significantly changed among the different groups even with top significance in the MREA analysis.
In amino acid metabolism, significant reductions in alanine, proline, and asparagine were detected in response to nux vomica exposure. Decreases in glutamate and aspartate indicated that fewer carbon and nitrogen sources were supplied to the TCA cycle for the production of energy, amino acids, and pyrimidines. Alanine, involved in glucose production and nitrogen cycling, was decreased in the treated group, which failed to act as a reply to compensate for energy deficiency. In addition, the reduction in pantothenic acid was associated with the dearth of materials for the synthesis of coenzyme A, which was involved in lipid and protein metabolism. 42 In addition, the reduction of most individual amino acids might have resulted from the urgent need for increasing synthesis of complex in the electron transport chain, oxidation of fatty acids, and TCA in cytoplasm.
For carbohydrate metabolism, significant changes were clearly observed in response to nux vomica exposure. In the galactose, citrate cycle metabolism, the significant increase of galactose and isocitric acid was observed in the L group compared with the B group indicating that the low-concentration nux vomica treatments affect the energy metabolism pathways to supply more energy to the zebrafish. While in the H group, the remarkable decrease of galactose and isocitric acid was shown compared with the L group, suggesting that the toxicity of nux vomica had hampered the synthesis and utilization of energy both in the glycolysis and the TCA pathway. The decreases in TCA cycle activity revealed a reduction of carbon and nitrogen sources for the production of energy and biosynthetic precursors of amino acids, lipids, and pyrimidines. Furthermore, the increase of the glucose-6-phosphate and pyruvic acid in the treated group added more information for the anaerobic glycolysis metabolism, which was used to provide energy that agreed with the previous report.43,44 Abnormal mitochondrial function was unfavorable for many biosynthetic and bioenergetic processes and could lead to liver function disorders.
Interestingly, some annotation terms of lipid metabolism were also affected, including fatty acids, glycerol acid, and glycerol. In the treated groups, the levels of palmitoleic acid were significantly decreased, while those of arachidic acid (ARA) were increased. The increase in ARA indicated that an inflammatory response might exist because ARA is an inflammatory biomarker.45,46 Besides, the decreased palmitoleic acid in the treated groups suggested that the β-oxidation of fatty acid was induced to satisfy whole-body energy requirements. Combined with the ARA results, the result suggested that nux vomica could lead to the dysfunction of β-oxidation of lipids and liver damage. Based on KEGG pathway analyses, lipid metabolism pathways, such as the glycerolipid metabolism and glycerophospholipid metabolism pathway, were significantly affected.
Nucleoside metabolism was significantly disturbed in the zebrafish larvae. As we know, xanthine is an important biomarker of ischemia and oxidative injury. Xanthine resulted from the decrease of adenosine triphosphate (ATP) and ultimately converted to uric acid by dehydrogenase. Thus, the increase of xanthine and uric acid denoted that the supply of ATP was disrupted. Similarly, based on the KEGG pathway analysis, the purine metabolism pathway was significantly affected.
Although no specific biomarker was observed, the potential consequences of the internal effects of nux vomica exposure on zebrafish liver were identified. In addition, the whole zebrafish in this experiment was used to explore the potential toxicity mechanism and remarkable results have been obtained, however, the effect of nux vomica on other tissues and organs is not ignorable. Therefore, to reduce the effect from the other issues, the exposure concentration we chose was as low as possible and the result showed that the liver was the most sensitive target. Thus, the great change in endogenous metabolites induced by nux vomica reflected the liver response. Furthermore, this work was the first to predict molecular mechanisms, regulatory targets, and potential biomarkers of liver toxicology of nux vomica in the in vivo zebrafish model, providing a reference for hepatotoxicity studies in the future in other models.
In the future, studies will also need to be performed to investigate the effects of nux vomica on DNA damage, cell membranes, and mitochondria-related mechanisms to broaden our understanding of the toxicity mechanism of nux vomica and to specifically assess the correlation of these metabolomic markers with established markers in other models. It should be assessed in longitudinal settings whether the identified metabolites are predictive of new-onset incident fat liver disease or of change in liver fat content over time. Finally, functional studies are warranted to explore the molecular mechanisms linking the observed metabolites.
Footnotes
Acknowledgment
This work was supported by special funding from the Beijing Municipal Science and Technology Commission for innovation environments and platform construction (grant no. Z16111000500000).
Disclosure Statement
No competing financial interests exist.