
Abstract
Abstract
Astyanax is an abundant fish genus in South America. Some species of this group are characterized by the presence of B chromosomes and absence of morphologically differentiated sex chromosomes. In this study, we used quantitative real-time polymerase chain reaction to characterize mRNA expression of dmrt1 in Astyanax scabripinnis gonads. Maturing gonads of males with the B chromosome overexpressed dmrt1. Our findings suggest that B chromosomes may have an adaptive role in A. scabripinnis sex determination and maintenance.
Introduction
Astyanax Baird and Girad, 1854, is a genus of freshwater fish abundant in South America. To date, their estimated diversity is nearly 200 species 1 with a wide distribution along Neotropical continent. 2 Astyanax contains numerous cryptic species complexes, with Astyanax scabripinnis being among the most representative. 3 This species complex has a conspicuous karyotype characteristic, described in several allopatric populations: the occurrence of B chromosomes. 4 B chromosomes are supernumerary genetic elements found in all eukaryotic groups, which differ from standard complement chromosomes (A chromosomes), did not pair with any A chromosome in meiosis and exhibit irregular forms of inheritance. 5 Little is known about details of genomic composition and the origin of these genomic elements, but it is a consensus that in most cases, B chromosomes are considered nonfunctional and have no essential genes whose presence does not affect the carrier's phenotype.6–8 However, with the advancement of molecular techniques and comparative population studies, functional evidence for this chromosome has been described for various organisms in greater detail about their origin and evolution. 9
Sex ratio and B chromosome occurrence has been obtained for A. scabripinnis populations and seasonality distortions was observed, with distortion of the sex ratio in behalf of females and their significant association with the frequency of these B chromosomes, with a highly disproportionate number of males without these chromosomes.10–13 Interestingly, although chromosomal variability is wide, no morphologically distinct sex chromosomes have been detected in A. scabripinnis. 14 Nevertheless, males during the reproductive season clearly possess secondary sexual characters such as bony hooks on the anal fin. 15 Hermaphroditism was recently recognized as typical for A. scabripinnis, although the genetic factors involved in their primary sex determination and gonad differentiation remain unknown. 16
Primary sex determination in vertebrates involves both genetic and environmental mechanisms, with temperature-dependent sex determination being a well-known example of the latter. 17 This complexity means that we still have not fully resolved how sex is determined for the majority of fish species. For instance, morphologically differentiated sex chromosomes are known for only 10% of species with described karyotypes18,19 and for 6% of Neotropical species. 20 Even in these cases, sex-determining (SD) genes generally remain unknown, with a few exceptions, such as amhy in Odontesthes bonariensis. 21
Multiple genes have been implicated in the regulation of sex determination cascade. 22 Some of these are related to mutations in the Doublesex Male (DM) domain, such as dmy/dmrt1Y, specific to medaka (Oryzias latipes).23,24 First described in Drosophila melanogaster as doublesex 25 and in Caenorhabitis elegans as mab-3, 26 dmrt1 encodes a transcription factor with a highly conserved zinc-finger motif. The gene is masculinizing, responsible for male gonadal development in metazoans and, depending on species, acts either as an upstream or downstream regulator in the sex differentiation cascade.27,28
In vertebrates, dmrt1/DMRT1 (Doublesex Male abnormal-3-Related Transcription Factor-1) encodes a transcription factor with a highly conserved DNA-binding motif (i.e., DM domain). The gene is critical to primary sex determination and testicular differentiation for many species. 29 Expression of dmrt1 is dimorphic, but a transcription imbalance can occasionally cause male-to-female sex reversal, as seen in some amphibians, 30 chicken, 31 and fish. 32 For instance, in the half-smooth tongue sole (Cynoglossus semilaevis), dmrt1 likely appeared around 30 mya and in parallel to the differentiation of a ZZ-ZW chromosomal SD system. During C. semilaevis development, both sexes exhibit sex-specific differential dmrt1 expression that apparently occurs through a dosage compensation mechanism. 33
To clarify the role of dmrt1 in A. scabripinnis sex determination and determine potential links to the species' unique B chromosome, in this study, we characterize the gene's expression among individuals with or without B.
Materials and Methods
Sampling site and specimens
Fifty-one A. scabripinnis specimens (25 males, 25 females, and 1 hermaphrodite) were collected from the Lavrinha Stream (S 22°43′09.6″, W 45°25′38.5″) in Guaratinguetá, Brazil. The mature specimens were collected during reproductive (November to January) season and the maturing, during nonreproductive (April to June) season. Specimens were anesthetized with 0.1% benzocaine and sacrificed. All procedures followed ethical guidelines for animal experimentation detailed by the Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio-proc 15115) and Comissão de Ética em Experimentação Animal (COEP-UEPG).
Histology
Gonads were dissected and fixed in Bouin solution for 24 h at room temperature (25°C). Fixed tissues were dehydrated, embedded in paraffin, sectioned (3–5 μm thickness), then counterstained with Hematoxylin and Eosin for sex identification. Gonads were classified as either maturing or mature according to reference 34.
Whole chromosome painting and fluorescence in situ hybridization
To detect B chromosome presence through whole chromosome painting (WCP), mitotic chromosomes were obtained following a published protocol. 35 Additionally, a 5 mm3 fin sample was preserved in 70% ethyl alcohol, then fully dissolved in acetic acid. Subsequently, a 3:1 Carnoy's solution was added to the fin sample. A microdissected B-chromosome probe was then amplified through degenerate oligonucleotide-primed PCR following previous methods. 36 Subsequent WCP of interphase nuclei obtained from fins was also performed according to published procedure. 16 Hybridization occurred under high-stringency conditions (2.5 ng/μL probe, 50% formamide, 2 × SSC, 10% dextran sulfate), following a general protocol. 37 Signals were detected with the Anti-Digoxigenin-Rhodamine antibody (Roche, Mannheim, Germany). Materials were counterstained with ProLong™ Gold Antifade Mountant with DAPI (Thermo Fisher, Burlington, Canada). Preparations were analyzed using an epifluorescence microscope (Olympus BX41, New York) coupled to an image capture system (CCD Olympus DP 71® and DP-Controller v. 3.2.1.276).
RNA extraction and cDNA synthesis
After dissection, 3 mm3 gonad samples were immediately placed in RNAlater (Ambion, Vilnius, Lithuania) for 24 h and stored at −80°C to preserve nucleic acids. Total RNA was extracted through homogenization in TRIzol (Invitrogen, Burlington, Canada) following the manufacturer's protocol, then purified with recombinant DNAseI (Invitrogen), and quantified through spectrophotometry. Quality was verified through visualization of 28S and 18S bands in 1% agarose gels. Next, cDNA synthesis was performed with 1 μg of total RNA, oligo-dT12–18 primers (Invitrogen), and reverse transcriptase Super Script III (Invitrogen). The product was then diluted at 1:3 in nuclease-free water.
Real-time quantitative polymerase chain reaction
The experiment analyzed 44 individuals split into five experimental groups per gonad classification: female with B, female without B, male with B, male without B, and hermaphrodite. The maturing gonad category included nine, four, four, six, and one individual per group, respectively. The mature gonad category included four, three, six, seven, and 0 individuals per group. Each sample was run in triplicate to correct interassay variation.
Real-time quantitative polymerase chain reaction was performed in the thermocycler Stratagene Mx3005P (Agilent Technologies, Waldbronn, Germany). Reaction volume was 12.7 μL, including 9.9 μL SYBR Select Master Mix (Life), 2 μL of cDNA, and 0.4 μL of each primer (10 mM). Primer338,39 was used to design specific primers for dmrt1 from existing sequences 40 and for reference gene β-actin from genomic sequences of Astyanax mexicanus in Ensembl. 41 All sequences amplified with the primers were confirmed by sequencing. The primers cover different exons to avoid amplification with contaminant gDNA. Thermocycling conditions were: 95°C for 10 min, followed by 40 cycles of 95°C for 30 s and 60°C for 1 min, and then ending with a temperature increase from 60°C to 95°C. The final step created a melting curve and guaranteed unique amplicons. Dilution curves for calculating primer efficiency were generated through serial dilution (1:10) of cDNA.
Samples were excluded if they were without Ct values or had inconsistent Ct between replicates (Ct difference >2 cycles). Analysis and quantification using the standard curve method were carried out with the MxPro software (Agilent Technologies). Expression values for dmrt1 bY and dmrt1 a were normalized using β-actin as reference gene control. Mean ± SEM data were analyzed with one-way analysis of variance (ANOVA) followed by Tukey's test in GraphPad Prism 7 (GraphPad Software, California). Significance was set at p < 0.05.
Results
Histology identification
Histological analyses of gonadal cross-sections confirmed sex identification (Fig. 1a, b) and morphology according to reference 16. A hermaphrodite-like individual was also identified with maturing gonads (Fig. 1c, d).
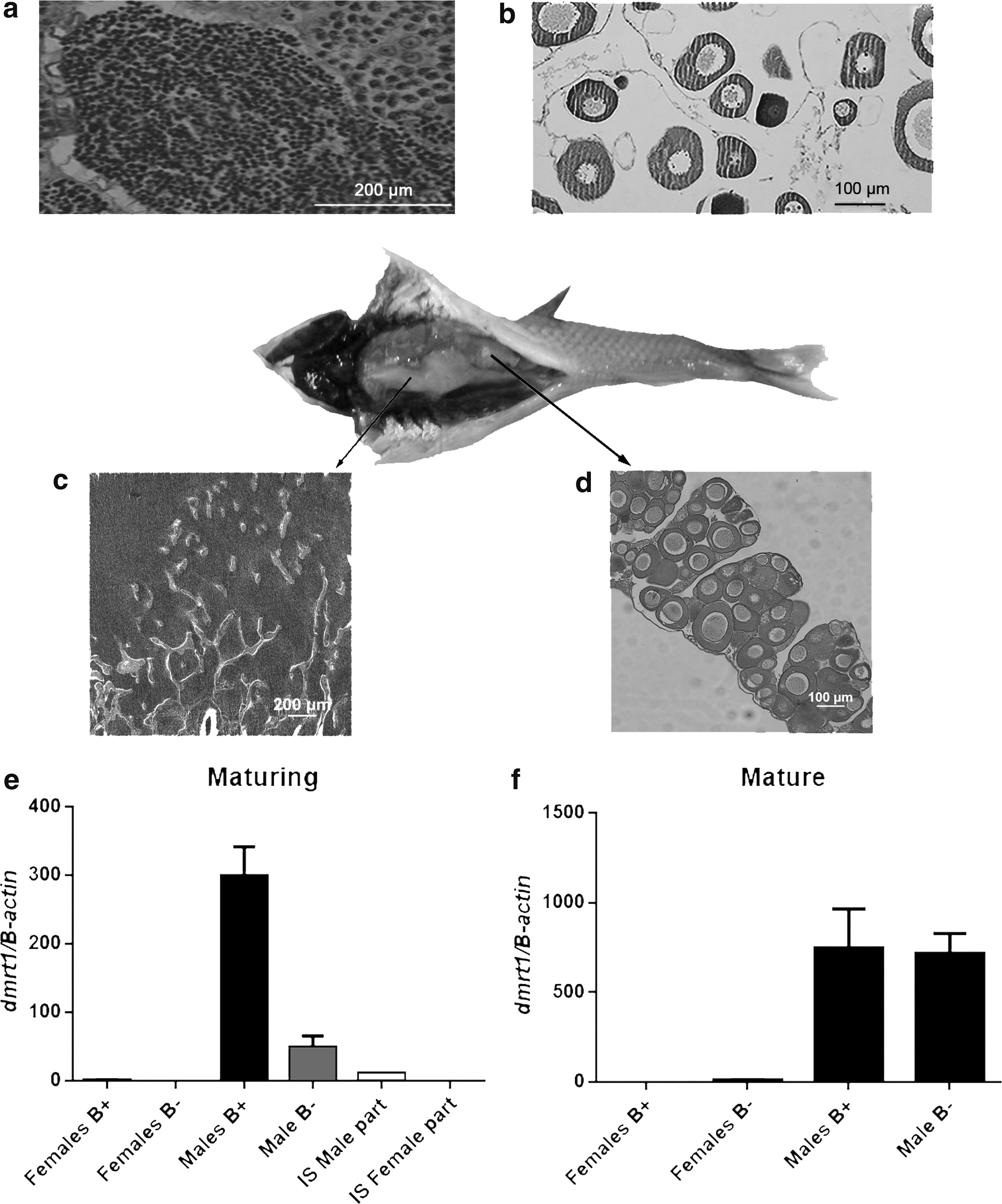
Histological sections of Astyanax scabripinnis gonads.
Detection of B chromosome through WCP and fluorescence in situ hybridization
The entire B chromosome was successfully visualized using WCP (Fig. 2). Signals in interphase nuclei were also detected in B+-bearing individuals. These results were used to verify B chromosome presence in all samples, as previously recommended. 16
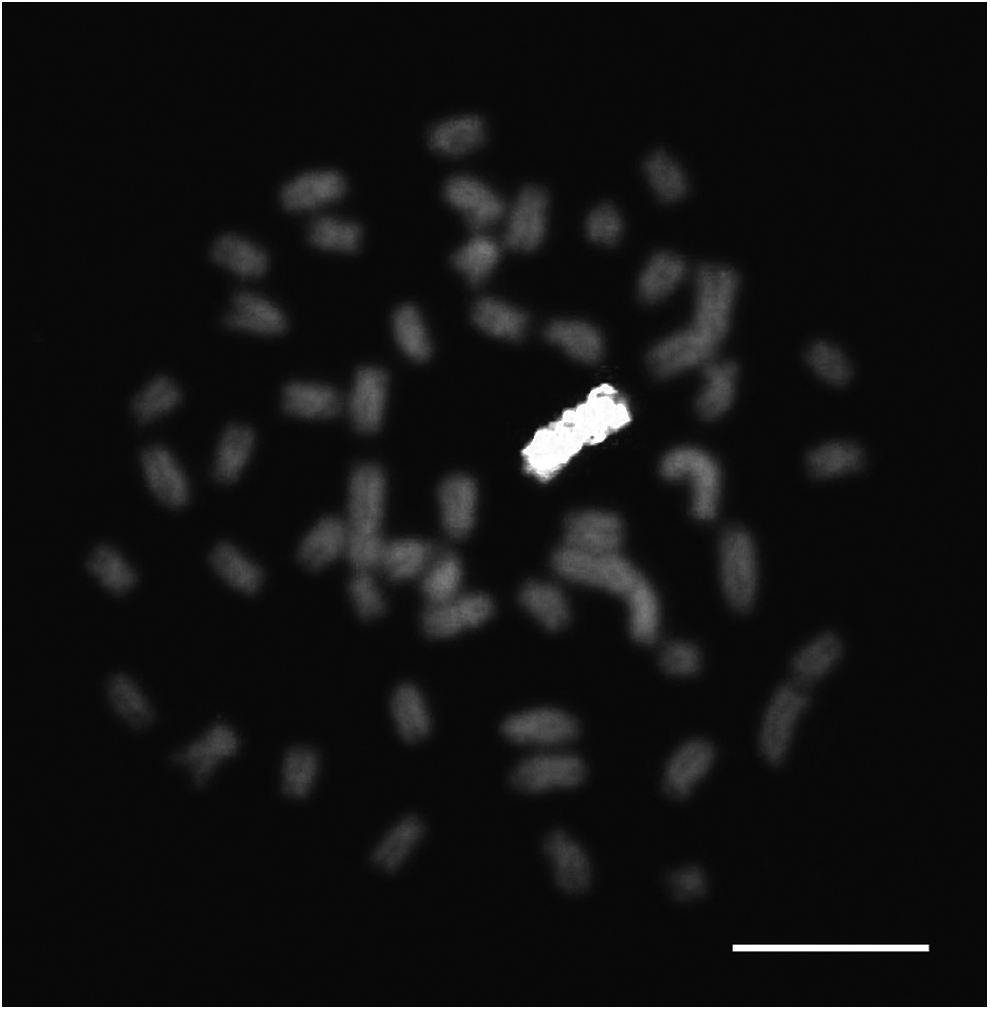
Whole chromosome painting with microdissected B-chromosome probe. Scale bar: 10 μm.
Sex, B chromosome, and gonad maturity influence dmrt1 expression
Both sex and B chromosome presence significantly affected dmrt1 mRNA expression. Additionally, gonad maturation stage influenced mRNA expression. Maturing male gonads with B chromosome exhibited significantly higher relative expression than others groups (Fig. 1e, f, and Table 1). The single hermaphrodite-like individual (with B chromosome) showed synchronized maturation of both female and male parts, apparently showing greater dmrt1 expression in the latter. The statistics showed no significant differences between the hermaphroditic parts, perhaps because of the low sample size (Fig. 1). Supplementary Tables S1 and S2 provide additional statistics.
Statistical Analysis (ANOVA with Tukey's Tests) of dmrt1 Expression in Female, Male, and Hermaphrodite Gonads of Astyanax scabripinnis With or Without B Chromosome
p < 0.0001, extremely significant.
p = 0.0001–0.001, extremely significant.
p = 0.001–0.01, very significant.
ANOVA, analysis of variance; B+, individual with B chromosome; B-, individual lacking B chromosome; IS, intersex; ns, not significant.
Discussion
This study is the first to observe that dmrt1 expression levels are highly associated with male sex, gonadal maturation stage, and B chromosome presence in A. scabripinnis. The dmrt1 gene is involved in sex determination and gonadal development in numerous vertebrates. 29 Evidence from fish (O. bonariensis 42 ; Halobatrachus didactylus 43 ) suggests that dmrt1 also affects spermatogenesis onset and maintenance. In teleosts, dmrt1 is expressed during male gonad differentiation.24,32,40,42,44,45 A functional copy specific to the Y chromosome (dmrt1Y) and one autosomal copy (dmrt1) has been reported in O. latipes (XX/XY sex system).23,24 In C. semilaevis, dmrt1 is localized to the Z but not in the W chromosome. Thus, genetic males (ZZ) have two copies of dmrt1, whereas genetic females (ZW) have only one.33,46 In fish, the chromosomal location of co-opted copies of dmrt1 is quite complex.
Male gonads of B+-bearing A. scabripinnis overexpressed dmrt1, suggesting a surprising adaptive role for the unique chromosome. Relatedly, B+-bearing A. scabripinnis females have a different reproductive activity peak, occurring during the summer (December), whereas B-lacking females, a reproductive peak extending from July to December. 16 Altogether, these observations imply a reproductive advantage for B+-bearing fish.
In zebrafish was demonstrated that adult testis had dmrt1 expression in nearly all stages of spermatogenesis, whereas majority of dmrt1 mutant develop as fertile females 47 Likewise, this condition is observed in mammals, which dmrt1 plays a crucial role in differentiating spermatogonia. The deletion of dmrt1 in spermatogonial stem cells has a progressive failure of spermatogenesis. These inferences show correlation between dmrt1 and longer period of spermatogenesis. 48
Thereby, B+-bearing males can have extended spermatogenesis that allows them to reproduce with similarly B+-bearing females outside the typical reproductive period for this species. 16 This increases the adaptive value and hence maintenance of the B chromosome in the population. A factor regulating dmrt1 expression may be present on the B chromosome of A. scabripinnis.
This inference led us to perform chromosome mapping analysis using dmrt1 gene sequence as probe in fluorescence in situ hybridization (data not show). We found signals in interstitial, equilocal regions of the B chromosome's short and long arms. This pattern is similar to the localization of As51, a repetitive DNA fragment 49 that is associated with transposon elements and microsatellites.50,51 However, we note that the likelihood of false localization on the B chromosome is high, because the used probe contains large intronic segments of dmrt1 (a mix of the cloned introns and exons) that increases signal detection during optical microscopy, resulting in the potential marking of other homologous regions.
We recently described hermaphroditism in A. scabripinnis, 16 a phenomenon apparently linked to fish populations inhabiting small, high-altitude rivers with unstable temperatures and seasonal flooding. This observation is in keeping with the idea that environmental disruption, along with biological factors, trigger hermaphroditism in fish. 52 Our current results indicate that to understand hermaphroditism, future studies should pay particular attention to dmrt1 expression during initial stages of A. scabripinnis ontogeny. Suggestively, dmrt1 regulation in A. scabripinnis influences both hermaphrodite occurrence and sex ratios; the latter tends to be unbalanced and has been linked to B chromosome presence. 12 Reflecting the importance of dmrt1 in testicular differentiation, a mutation in the gene caused ovaries to develop in XY O. latipes. 32 In contrast, dmrt1 mutation in C. semilaevis disrupted both spermatogenesis and ovarian development. 46 These data support our previous observations of A. scabripinnis hermaphrodites and confirms the importance of dmrt1 in the species' sex differentiation and maintenance.
Balanced sex ratios (1:1) is an adaptive condition 53 ensured by heterogametes in organisms with morphologically differentiated sex chromosomes. However, this condition has not been found for the majority of teleosts, including A. scabripinnis. In fact, such sex chromosomes occur in only around 6% of fish species.20,54 For most fish, sex determination and sex ratios are exclusively due to the expression of correlated genes following an unresolved, complex cascade that is heavily influenced by environmental factors. 22 A strong environmental influence is not unique to fish and has also been reported in other taxa, including reptiles, where egg incubation temperature determines sex. 55 In amphibians, environmental factors also have a huge impact on sex determination, and sex can be converted through steroid action, as seen in Rana rugosa. 56 Also among homoeothermic vertebrates, the epigenetic influence of temperature on sex determination is observed. Among the ZZ/ZW sex chromosome system of birds, dmrt1 overexpression in embryos induces the development of male gonads. 57 Currently, much remains to be understood regarding epigenetic effects affecting vertebrate sex determination and differentiation. Genes such as dmrt1 are strong candidates for deeper investigation into this topic.
To conclude, our findings support a possible adaptive role for B chromosome presence in A. scabripinnis. Furthermore, this study highlights the importance of dmrt1 on sex determination and maintenance.
Footnotes
Acknowledgments
The authors are grateful to the Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio). This work was supported by the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, process number 407187/2016-2), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES, scholarship to JPC, finance code 001), and the Fundação Araucária (Fundação Araucária de Apoio ao Desenvolvimento Científico e Tecnológico do Estado do Paraná, grant number 16174).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.