
Abstract
Abstract
Fixation and decalcification can alter protein structure in tissues, influencing the efficacy of primary antibodies routinely used in immunohistochemical (IHC) staining. Histologic examination of zebrafish requires both processes, making staining and analysis potentially challenging. Here, we investigated the effects of common fixation and decalcification protocols on IHC staining in zebrafish. We also identified zebrafish-reactive and -specific antibodies for use in research and diagnostics. For several of the antibodies, time spent in Dietrich's fixative containing 2% glacial acetic acid or 3.4% formaldehyde followed by decalcification with ethylenediaminetetraacetic acid (EDTA) significantly impacted IHC staining quality, particularly regarding staining intensity. Protocols utilizing shorter fixation times produced higher-quality stains. In addition, individual markers were variably affected by the type of fixative. Dietrich's fixative significantly reduced staining quality for the “neural” markers: glial fibrillar acidic protein, chromogranin A, S100. A negative time-dependent effect of fixation on staining quality was found for several antibodies: muscle actin (Dietrich's only), cytokeratin AE1/AE3, chromogranin, and S100. Neither decalcification protocol had a statistically significant negative time-dependent effect on staining quality. Based on our results, we suggest shorter fixation and decalcification protocols to best preserve IHC staining quality as well as recommend deliberate selection of the fixative used depending on the protein of interest.
Introduction
Demand for zebrafish-specific immunohistochemistry (IHC) is increasing as this animal model is utilized more often to study both experimentally induced and spontaneous neoplasms. Experimentally induced neoplasms in zebrafish are used to model many human cancer types. Spontaneous neoplasia is a common occurrence in some zebrafish facilities. 1 Initial classification of neoplasms in all species is based on histomorphologic characteristics as observed by light microscopic examination of stained slides.
IHC is the most widely used tool to further characterize the phenotype of neoplastic cells. Most validated antibodies that are commercially available for IHC are directed toward epitopes of human proteins. Antibodies directed at mammalian proteins are not optimized for use on zebrafish tissue sections. 2 Libraries of zebrafish-specific antibodies are now available, but few zebrafish-specific antibodies have been optimized or appropriately validated for use in paraffin sections. 3
Tissue fixation and decalcification are necessary steps when preparing whole zebrafish for histological examination. Both processes can negatively affect protein preservation and subsequent detection by IHC. 4 The effect of fixation and decalcification on IHC staining has been systematically analyzed in a few specific settings, such as human bone marrow samples and select human cancers. 5 Very limited data are available for zebrafish. One study analyzed IHC performance in the context of whole mounts; another in the context of embedding in plastic resin.6,7 IHC staining of zebrafish for cytokeratin is impacted by fixative or length of decalcification (Löhr, personal observation while working on Paquette et al. 2015). 8
Specific guidelines have been established for optimal processing of some human tissues for IHC. 5 Similar guidelines are needed to render IHC results from experimental and diagnostic examinations of zebrafish reliable and reproducible.
With that, the goals of this study were twofold: (1) to examine the effect of fixation and decalcification on IHC staining in zebrafish to help formulate guidelines for fixation and processing of zebrafish for IHC applications in research and diagnostics, and (2) to validate a panel of antibodies—both zebrafish-specific and interspecies-specific—for use on routine-processed zebrafish samples.
Materials and Methods
Zebrafish husbandry and handling
Zebrafish were maintained in a Biosafety Level 2 facility in flow-through tanks according to an Animal Care and Use Protocol approved by the IACUC at Oregon State University. Zebrafish of both sexes were raised and maintained at 25–27°C with sufficient aeration and a 14:10 h light–dark photoperiod. Zebrafish were euthanized once they reached ∼1.5 cm body length (∼4–6 weeks of age) and the coelomic cavity opened along the ventral midline.
Fixation and tissue processing
Whole zebrafish were fixed either in Dietrich's fixative containing 2% glacial acetic acid for 48 h (D48), 1 week (D1w), and 4 weeks (D4w) and held in 70% ethanol until processing or in 10% neutral-buffered formalin (ThermoFisher Scientific, Waltham, MA) for 48 h or 2 weeks followed by decalcification in CalExII (formic acid; ThermoFisher Scientific) for 48 h (N48C48 and N2wC48) or 1 week (N48C1w and N2wC1w) and held in 70% ethanol until processing. Each of the seven experimental groups comprised three fish (total 21).
Fixed and decalcified zebrafish were processed as one batch and embedded with each block containing fish from three to four different experimental groups to account for variability in staining between slides. Before embedding, each group of fish was inked to help identify the groups as small samples can be jostled during processing as follows: D48, none; D1w, yellow; D4w, blue; N48C48, orange; N48C1w, green; N2wC48, black; N2wC1w, red. To improve the probability of capturing all of the desired tissues, one longitudinal and three cross-sections of fish from each group were placed in cassettes.
Immunohistochemistry
Sections taken from the seven paraffin blocks were stained for a panel of zebrafish-reactive and -specific antibodies: cytokeratins (pan cytokeratin markers wide-spectrum cytokeratin [WSS] [Dako Z0622; 1:500] and AE1/AE3 [Dako M3515; 1:100]), glial fibrillar acidic protein [GFAP] [Genetex GTX27260; 1:2000], S100 [Dako Z0311; 1:1000], chromogranin A [Dako A0430; 1:1000], muscle actin [Dako M0635; 1:100], vimentin [Dako M7020; 1:500], and otoferlin [DSHB HCS-1; 1:20]. IHC was performed according to standard operating procedures of the Oregon Veterinary Diagnostic Laboratory. First, 4–5-μm-thick sections on charged slides (Tanner Scientific, Sarasota, FL) were baked at 60°C for 1 h and then rehydrated.
Depending on manufacturer's protocols, slides were pretreated with either high-temperature antigen-retrieval (chromogranin A, GFAP, otoferlin, S100) in a microwave pressure cooker (Viking Tender cooker) using Dako Target retrieval solution (s1699) for 10 min or were enzymatically digested with Proteinase K (Dako S3020) for 5 min (CK [WSS]; AE1/AE3, Vimentin). No pretreatment was performed for muscle actin. Using a Dako Autostainer, primary antibodies were diluted in Dako antibody diluent (s3022) and applied to the slides for 30 min at room temperature. Slides were then immersed in chromagen Nova Red (Vector Laboratories, SK-4800) for 5 min, rinsed with distilled water, followed by counterstaining with hematoxylin (Dako s3302), which was diluted 1:3 in distilled water for 5 min, rinsed with distilled water, run down to xylene, and coverslipped.
Sections of normal dog tissues served as positive and negative method controls for all the aforementioned antibodies, save for HCS-1, which contained internal positive controls (i.e., lateral line). Sections of zebrafish reacted with none-immune serum served as negative controls for each sample.
Considering the subjectivity in the evaluation of IHC results, the slides were read blindly and independently by two ACVP-certified pathologists (Löhr [CVL], Spagnoli [STS]) and one senior anatomic pathology resident (Meritet [DMM]). Each observer assigned a grade to each individual fish on a slide, identified by their ink color. Grade was determined as follows: 0 = no staining; 1 = weak and patchy staining in appropriate tissues; 2 = weak and diffuse staining in appropriate tissues; 3 = moderate staining in appropriate tissues; and 4 = strong staining in appropriate tissues.
The level of intensity was compared to the positive control (assigned a grade of “4”) for all antibodies, save for cytokeratin AE1/AE3 for which the positive control was weaker than some of the test slides. For cytokeratin AE1/AE3, the level of intensity was comparatively based on the strongest staining zebrafish (assigned a grade of “4”). After individual scoring, graders reviewed slides together to generate a consensus score.
Staining patterns were noted by each pathologist. Staining pattern was defined as tissues with immunopositive cells and, if relevant, location within tissue (i.e., perivascular); subcellular location was not specified. Nonspecific staining, that is, immunopositivity in tissues not known to harbor the antigen of interest, was recorded as tissue type, cell type, and intensity.
Statistics
To more objectively assess the relationship between the time spent in fixative and the staining intensity, regression values were calculated using Microsoft Excel 2016®. For each antibody, the average staining intensity scores (dependent variable) were plotted on the y-axis against their corresponding time spent in fixative (measured in days, independent variable) along the x-axis. Using Excel, an R2 value was calculated. For the purposes of this study, we considered an R2 value >0.95 to indicate a strong relationship (specifically, 95% of the variance in staining intensity can be explained by time spent in fixative). However, R2 values do not indicate the direction of relationship, and so this has been demonstrated with correlation coefficients. A correlation coefficient approaching or equaling −1 or 1 indicates a strong inverse or direct relationship, respectively.
To investigate whether or not the duration of decalcification negatively influences IHC staining for a particular antibody, a Kruskal–Wallis test, followed by Dunn's multiple comparison test, was performed using GraphPad Prism 7® software to detect whether there is a statistically different staining intensity between groups. Significance was set at p < 0.05.
Interobserver agreement was calculated by dividing the number of events that all three pathologists agreed on by the total number of events.
Average IHC intensity scores were compared to consensus scores for each antibody using a Wilcoxon signed-rank test with a p-value <0.05 indicating significance (GraphPad Prism 7).
Results
Immunohistochemistry
Each pathologist (DMM, CVL, STS) blindly evaluated and semi-quantitatively scored longitudinal and cross-sectioned (inked) zebrafish (Fig. 1A–F). Although several sections—both longitudinal and cross-sections—were provided for each group, not every tissue was captured on each slide (e.g., spinal cord was missing from several longitudinal sections). The average IHC intensity scores and consensus scores are graphically depicted in Figure 2A–F. Muscle actin was not reviewed as a group given the relatively consistent scoring done individually. For consensus statement, only the cross-section blocks were utilized unless there was a limited amount of material necessitating evaluation of the longitudinal blocks as well. General trends and staining patterns noted individually were also demonstrated by consensus.
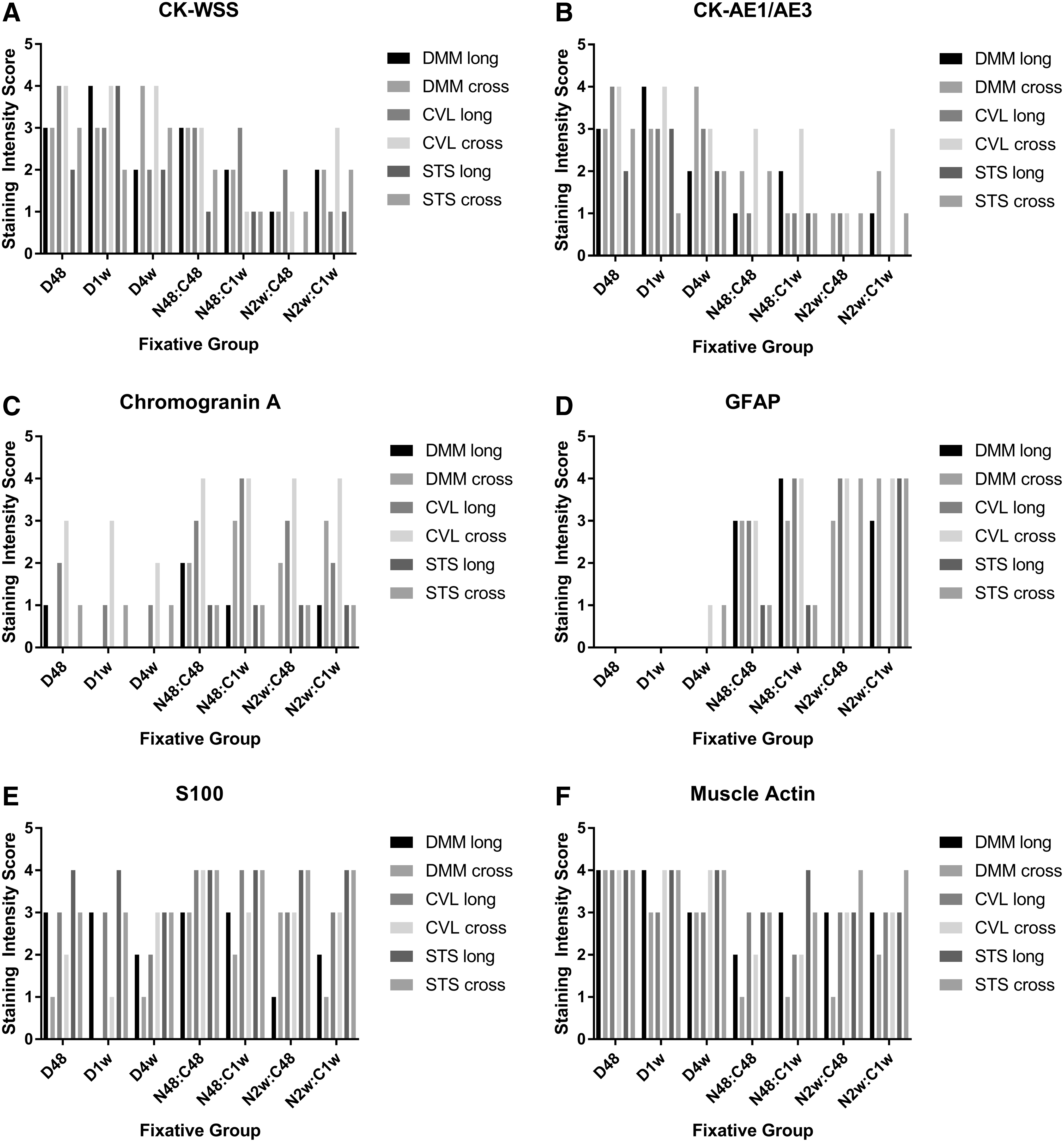
IHC staining scores by observer. Graphical representation of staining intensity for each group of zebrafish as determined by each pathologist (Meritet [DMM], Löhr [CVL], Spagnoli [STS]).
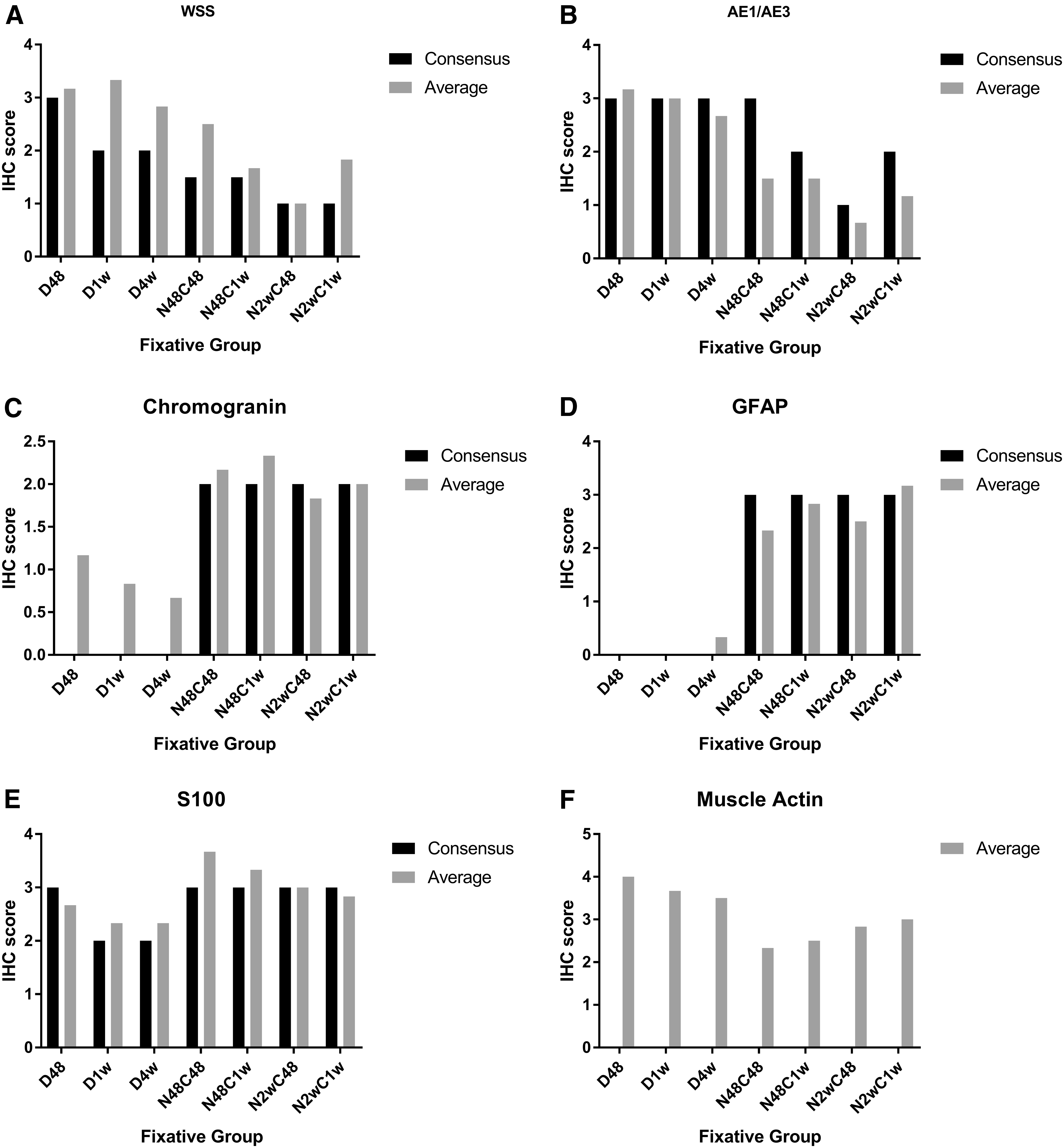
Average and consensus IHC scores
A compilation of staining patterns and intensity for each of the examined antigens follows.
For cytokeratin WSS (CK-WSS), moderate to strong staining was present in the skin of all zebrafish fixed in Dietrich's solution (Fig. 3A–C), with a slight decrease in staining intensity at the 4-week time point, whereas those fixed in neutral buffered formalin (NBF):CalEx exhibited weak to moderate staining (Fig. 3D–G). For both fixatives, weak immunopositivity—often described as a blush—was present in the liver and lens epithelium as well as in the pharyngeal plate; the gill and renal epithelium were uniformly immunonegative. With regard to the intestinal epithelium, the apical aspects stained strongly, while there was no staining of the basolateral membranes. In addition, the physaliferous cells of the notochord remnant exhibited moderate staining as did the gut contents.
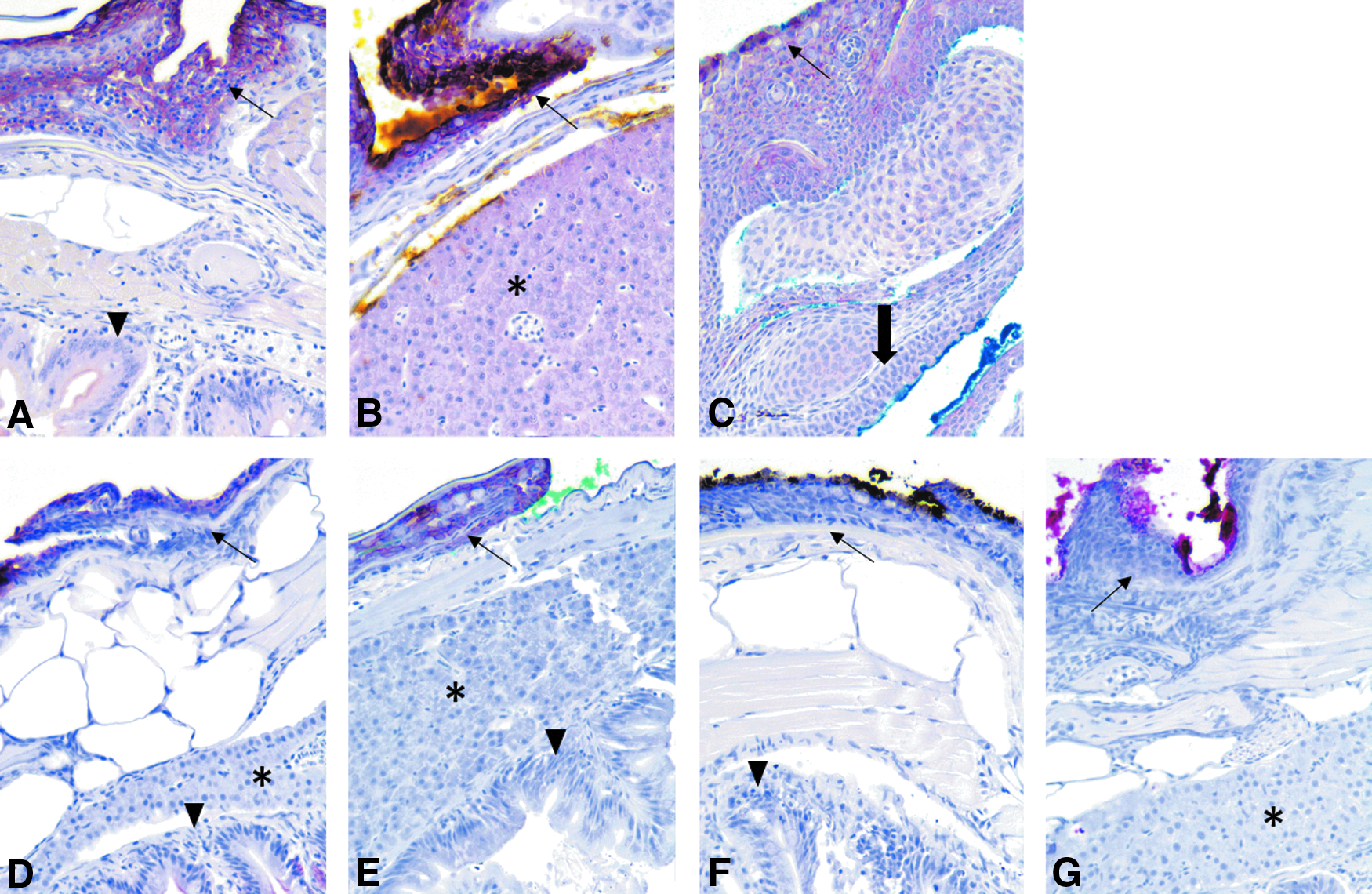
IHC images for CK-WSS. Immunohistochemistry for WSS. Dietrich groups shown in chronological order [
Staining for cytokeratin AE1/AE3 (CK-AE1/AE3) showed similar staining intensity with regard to the fixation groups as CK-WSS (Fig. 4A–G). In addition, tissue staining patterns were similar to those described for CK-WSS.
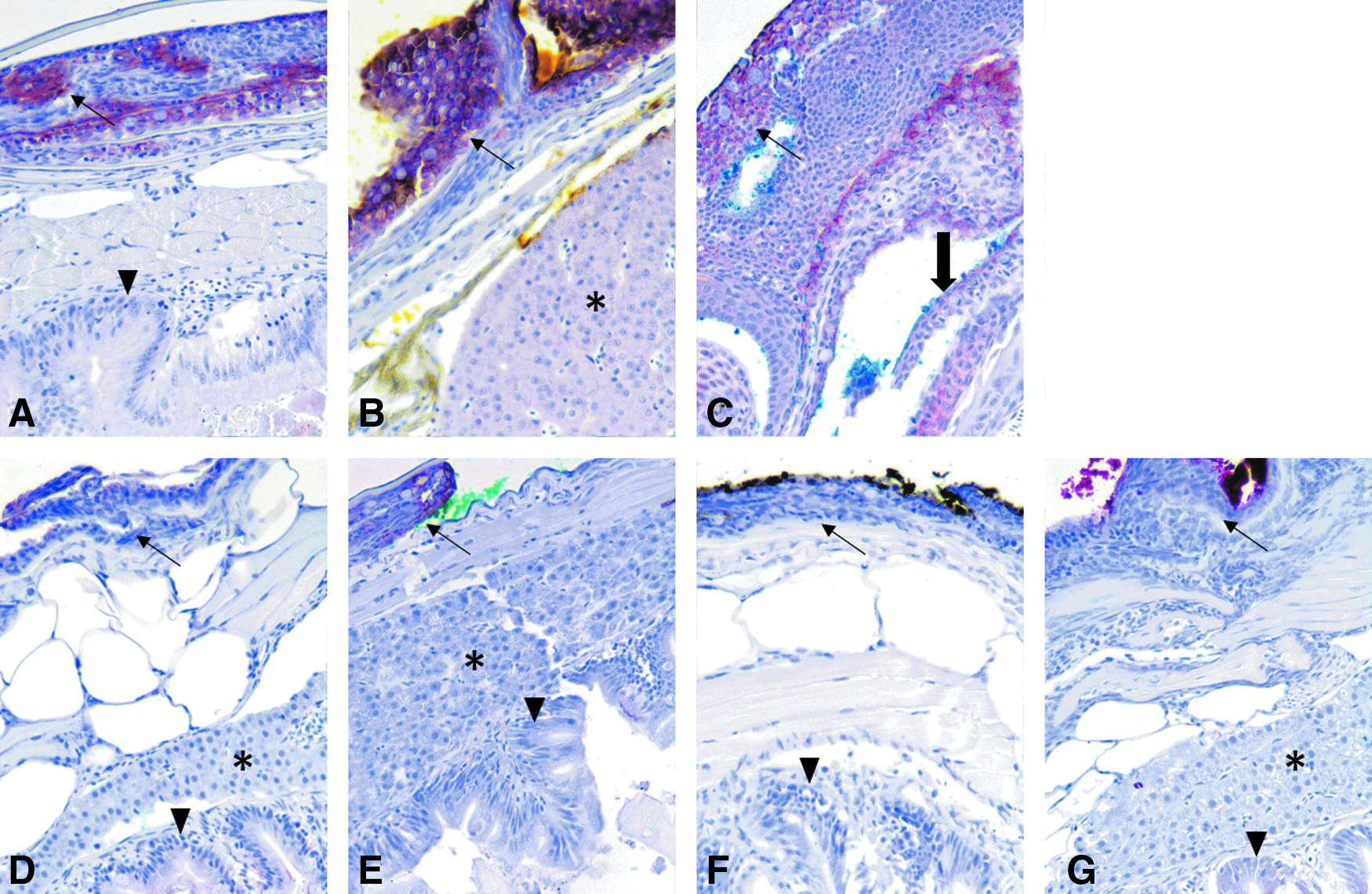
IHC images for CK-AE1/AE3. Immunohistochemistry for cytokeratin (AE1/AE3). Dietrich and NBF:CalExII groups shown in the same order as in Figure 3
Unlike with cytokeratins, the NBF:CalEx protocols resulted in superior staining intensity for chromogranin A (Fig. 5D–G) than did those fixed with Dietrich's (Fig. 5A–C). Interestingly, the different time points did not greatly alter staining intensities. Regarding the NBF:CalEx groups, staining for chromogranin A highlighted the neuronal cell bodies in the central nervous system and peripheral ganglia as well as endocrine cells within the pituitary gland and thyroid follicular cells. There was diffuse blush in non-neural tissues, including liver, skeletal/striated muscle, gastrointestinal (GI) contents, retina, otolith, cartilage, various epithelia (e.g., skin, GI, kidney), and pancreas. Contrary to the NBF:CalEx groups, the Dietrich groups collectively showed faint staining of peripheral axons. Although these groups exhibited less non-specific staining, the targeted tissues, including peripheral ganglia and endocrine organs, were immunonegative.

IHC images for chromogranin A. Immunohistochemistry for chromogranin A. Dietrich and NBF:CalExII groups shown in the same order as in Figure 3
Similar to chromogranin A, the NBF:CalEx groups, regardless of duration, had moderate to intense staining with the GFAP antibody (Fig. 6D–G), while the Dietrich groups showed little to no staining (Fig. 6A–C). In those exhibiting staining, the antibody to GFAP diffusely labeled spinal cord axons with occasional brain axons; the latter staining typically occurred within paraventricular areas. There was blush staining of the GI epithelium and gut contents.
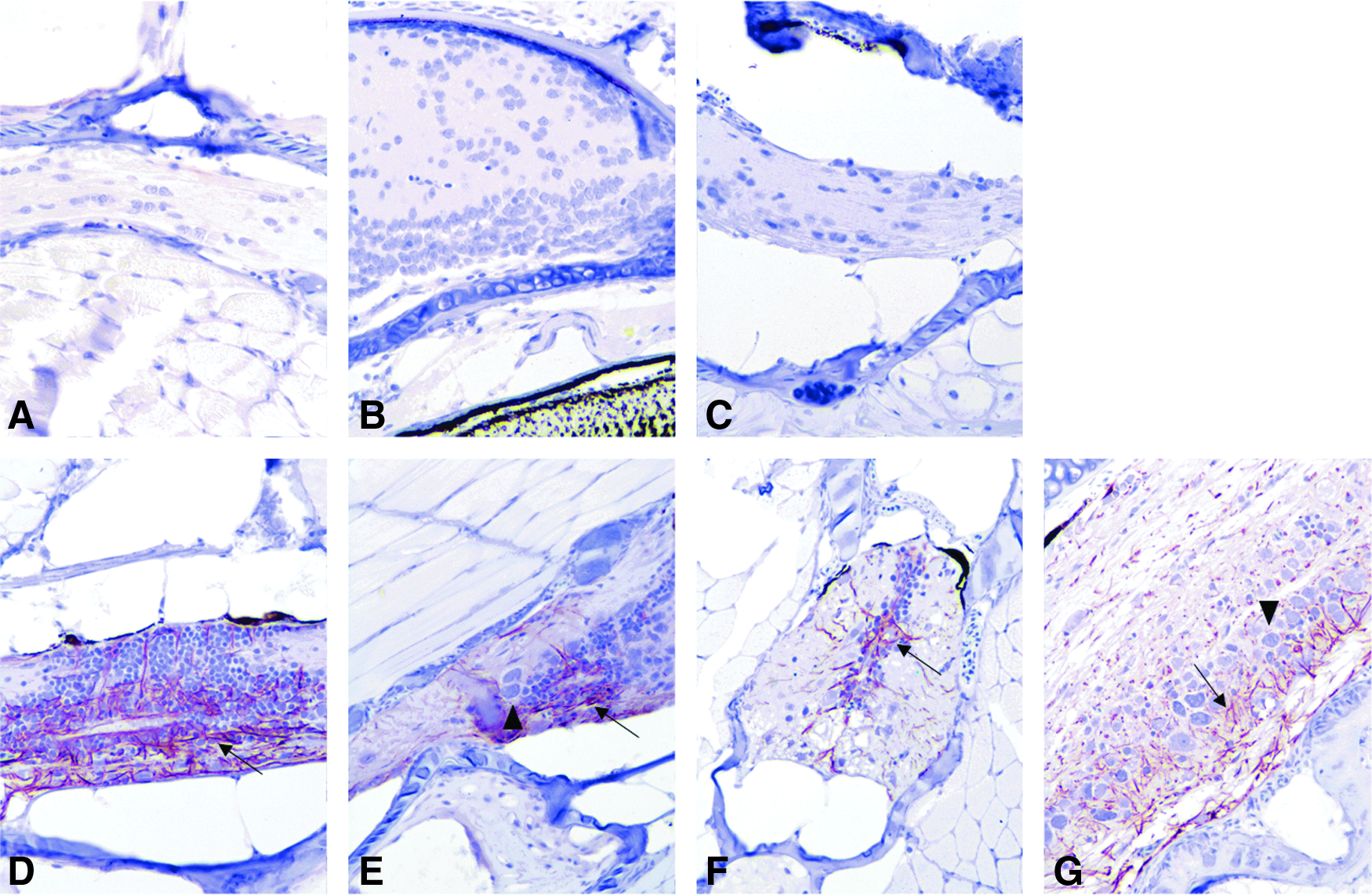
IHC images for GFAP. Immunohistochemistry for GFAP. Dietrich and NBF:CalExII groups shown in the same order as in Figure 3
Similar to chromogranin A and GFAP, S-100 staining intensity was most consistent and moderate to strong using the NBF:CalEx protocols (Fig. 7D–G), particularly with the 48-h fixative and decalcification (N48C48) group, compared with the staining noted in the Dietrich groups (Fig. 7A–C). There was diffuse, sometime fibrillar, staining of the neuropil in the brain with strong staining of glial cells and ependyma. In addition, neuronal cell bodies in the spinal ganglia occasionally exhibited moderate staining, though staining was absent in myenteric ganglia. There was weak to moderate staining of the olfactory epithelium as well as moderate staining of the ciliated cells in the ear (hair cells). There was diffuse blush staining of the skin with moderate to intense staining of cutaneous mucous cells. Rare blush staining of skeletal muscle was present.
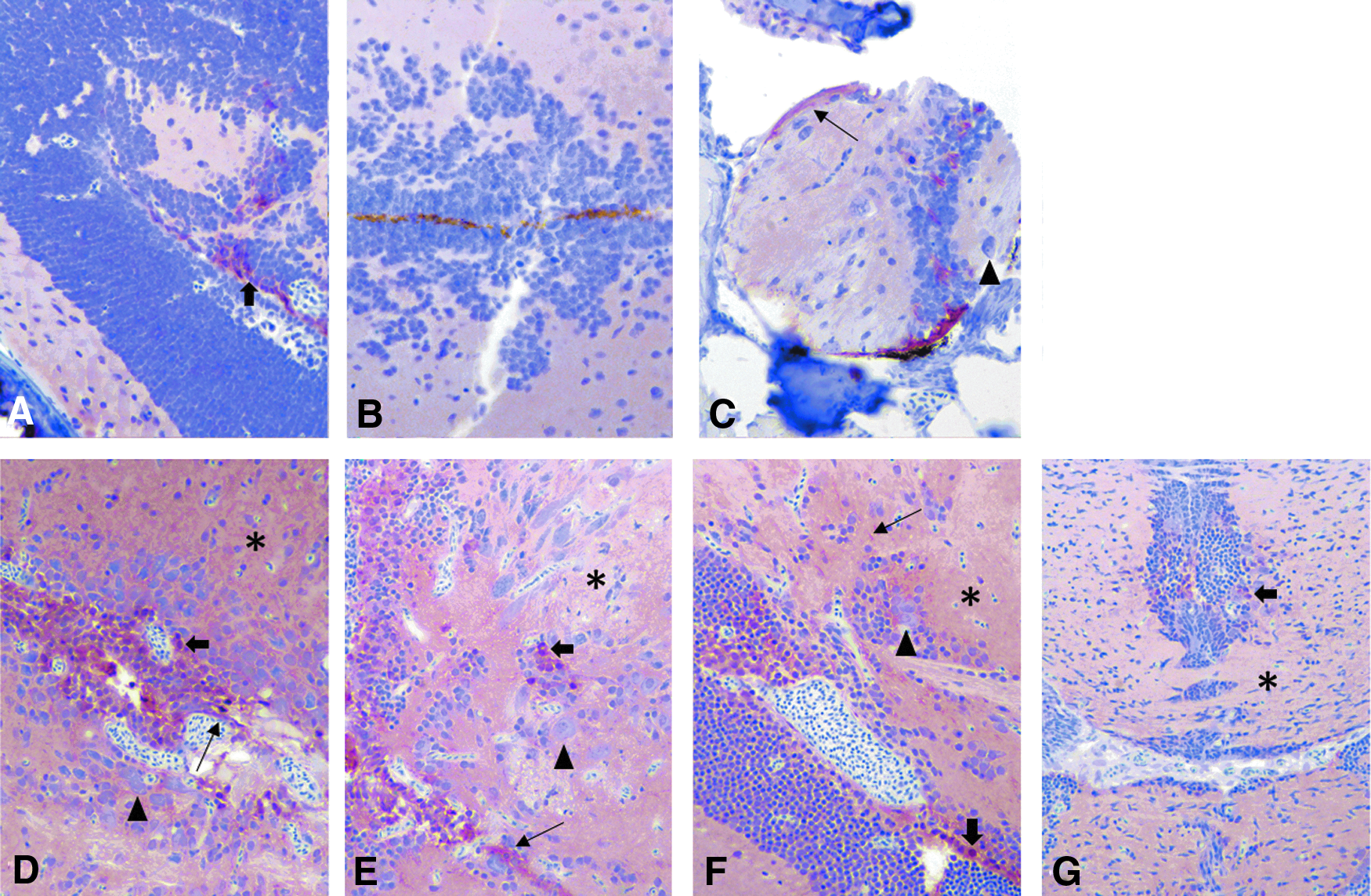
IHC images for S100
While there was moderate to strong staining for muscle actin in all groups, there was a modest increase in staining intensity for the 48-h Dietrich group (D48). Moreover, the Dietrich groups exhibited consistently moderate to strong staining, while the NBF:CalEx groups occasionally resulted in weak staining. Staining for muscle actin was strong in skeletal muscle and variable in smooth muscle in the gastrointestinal tract and within arteries. There was blush nonspecific staining of the liver and gut contents. Schreck cells within the skin stained weak to moderate.
Due to processing artefacts, the lateral line and labyrinth were frequently missing from the cross-sections, preventing adequate analysis of staining quality of the antibody raised against otoferlin between groups. However, when hair cells were present in sections, they were stained intensely irrespective of fixative and decalcification protocols.
Statistics
Regression analysis and correlation coefficients found strong inverse relationships between the time spent in fixative and staining intensity score for the following fixative–antibody combinations: Dietrich–CK-AE1/AE3, Dietrich–chromogranin A, Dietrich–muscle actin, and NBF:CalEx-S100 (Table 1, bold numbers). A strong direct, or positive, time-dependent relationship was found for NBF:CalEx–muscle actin (Table 1, superscript “a”). The remaining fixative–antibody combinations had an inverse, weak to moderate time-dependent relationship, save for GFAP that had a positive weak to moderate time-dependent relationship using both fixatives.
Comparison of Staining Intensity Scores for Each Antibody Across Fixative and Decalcification Protocols
Regression values (first row per antibody) and correlation coefficients (second row per antibody) to determine the relationship between time spent in fixative and average staining intensity scores for each antibody tested as well as statistical comparison of staining intensity scores between groups for each antibody tested. Bold denotes a strong relationship. Italicized groups indicate a negative mean rank difference.
Only significant values reported.
Denotes an upward sloping trend line. All others in bold are downward sloping.
NBF, neutral buffered formalin; CK, cytokeratin; GFAP, glial fibrillary acidic protein; CK-WSS, cytokeratin wide-spectrum cytokeratin.
Regarding the effect of decalcification duration, staining for CK-WSS showed the only significant difference in staining intensity using the same type of fixative/decalcification, which was between the N48:C48 and N2w:C48 groups (p = 0.0274). Given that the samples in these groups spent the same amount of time in ethylenediaminetetraacetic acid (EDTA), this difference was dependent on time in fixation and further supporting results presented above.
However, when comparing the different fixative/decalcification protocols, several groups exhibited statistically significant variations in staining intensity for some of the antibodies (Table 1). Pertaining to these groups, only for GFAP fixation in 3.4% formaldehyde followed by decalcification with EDTA yielded significantly superior staining quality compared with Dietrich's solution as indicated by a negative mean rank difference (Table 1, italicized numbers). For the other antibodies, shorter Dietrich's solution protocols were superior compared with longer periods spent in 3.4% formaldehyde followed by decalcification with EDTA. Based on these findings, shorter fixation periods overall yielded higher staining intensity scores, and Dietrich's solution negatively impacted staining intensity for GFAP compared with 3.4% formaldehyde followed by EDTA.
Observers agreed on 69 out of 294 observations (interobserver agreement = 23.47%). Interobserver disagreement primarily occurred with intensity of staining. Weak to negative staining of epithelial parenchymal organs with cytokeratin markers resulted in a score of 0 to 1 despite intense cutaneous epithelial staining. In rare instances, not all organs were contained within the examined sections for a given antibody, and some observers recorded this as 0 (no staining). This may have artificially inflated disagreement about staining intensity among observers. Despite the low interobserver agreement, the only antibody for which the average IHC intensity score significantly differed from the consensus score was CK-WSS (p = 0.0313).
Discussion
Overall, the time spent in fixative/decalcification solutions did not have a substantial impact on IHC staining intensity, though statistical analyses found that shorter Dietrich's protocols significantly improved staining quality for both cytokeratins and muscle actin compared with longer NBF:CalEx protocols. The opposite was true for GFAP in that both short and longer NBF:CalEx protocols resulted in better staining quality than both short and longer Dietrich's protocols. Furthermore, regression analyses revealed a strong inverse relationship between time spent in fixative and staining intensity for several antibodies (i.e., strong staining associated with shorter amount of time spent in fixative). Interestingly, in one instance—muscle actin in NBF:CalEx—strong staining was associated with longer times spent in fixative. It is likely that poor interobserver agreement impacted the results.
As previously mentioned, there is a considerable degree of subjectivity when evaluating staining quality of IHC, thus allowing for greater interobserver variation. For instance, some observers grade a cytokeratin marker as strongly staining if the cutaneous epithelium is highlighted even though there is weak to negative staining in visceral epithelial organs (e.g., liver, kidney, gut epithelium), while others assign a low score. Pathologists addressed and resolved these discrepancies in this study by arriving at consensus scores. Moreover, the incomplete sections and missing tissues may have influenced scores, thus contributing to interobserver variation.
In addition, the categorical evaluation criteria may have been too vague, leaving much open to interpretation from some observers compared to others. Most IHC studies involve binary data (present/absent), while this study entailed ordinal data, which inherently assumes some measure of ambiguity when assigning events to a certain category. In fact, with semi-quantitative scoring systems, more precise language involving percentage of staining cells and exact tissue distribution can enhance interobserver repeatability. 9
However, despite the low interobserver agreement, there was no statistically significant difference between the consensus scores and average IHC scores for most of the antibodies evaluated. It is suspected that the significant difference between consensus and average IHC scores for CK-WSS reflected the vague grading criteria. Specifically, CK-WSS variably stained different types of epithelia; so, while the skin stained strongly, other epithelia such as the gill and intestinal epithelium were immunonegative, raising the questions as to what exactly defines “appropriate tissues” and how an overall score is determined.
An interesting and unexpected finding is that the superior fixative/decalcification protocol varied among the individual IHC markers. The “neural” markers—chromogranin A, GFAP, and S-100—collectively showed stronger staining with NBF:CalEx protocols, while cytokeratins and muscle actin worked better with Dietrich's protocols. For both 10% NBF and Dietrich's solution, the primary fixative agent was formaldehyde, which utilizes a protein cross-linking mechanism. 10
Interestingly, all three “neural” markers underwent heat-induced antigen retrieval (HIAR) before IHC staining, whereas sections stained for cytokeratins were digested with proteinase K. HIAR has been found to unmask many antigens altered by formaldehyde-based fixatives, specifically NBF. 10 The weak to absent IHC staining for “neural” markers used in Dietrich-fixed groups may be the result of some persistent epitope masking caused by another component of this solution. For example, the ethanol in Dietrich's solution causes protein denaturation via dehydration reactions, resulting in protein coagulation and tissue shrinkage. 11
Furthermore, alcohol-based fixatives, including Dietrich's solution, are often combined with acetic acid to reduce the amount of tissue shrinkage, which also introduces hydrogen bonds in the tissue as part of the fixation process. Alternatively, formaldehyde creates cross-links between certain molecules, namely lysine residues, to fix tissues and has been shown to preserve secondary and tertiary protein structures.12,13 Therefore, it is reasonable to think that the differing methods of fixation can result in different patterns of epitope masking and subsequent differences in IHC staining.
The exact mechanism underlying the staining characteristics for different fixatives is uncertain and beyond the scope of this article. Based on these results, however, it may behoove researchers using IHC to investigate neural components in formalin-fixed, paraffin-embedded (FFPE) zebrafish to fix with 10% NBF followed by CalExII, instead of the standard Dietrich's solution. Moreover, prior studies have shown that longer fixation in formalin reduces IHC staining. 14
Assessing the effect of different decalcification periods for the NBF groups, the longer decalcification protocol (7 days in CalExII) resulted in significantly increased average staining intensity scores for the N2w group, which is contrary to the expected effect. Similar to the lack of consistent trends with time spent in fixation, this may be related to interobserver variation. Additional studies implementing more objective scoring criteria may help achieve greater interobserver agreement. Despite our results, other publications and prior anecdotal observations suggest that longer decalcification protocols adversely affect IHC staining. 4
With regard to tissue staining patterns, we cannot explain the lack of gill and renal epithelium staining with cytokeratins. Initially, a stain penetration issue was suggested, given the blush staining of other internal epithelia (e.g., liver, lens), but this would not account for the moderate staining of physaliferous cells in the notochord. In a previous publication by Paquette et al., the same CK-WSS and CK-AE1/AE3 stained both renal and gill epithelium. 8 The fish used in that study were fixed in Dietrich's solution, though the duration of fixation is uncertain as these were archival cases.
It is possible that our fixation times exceeded that used in the prior study and may have altered antigen accessibility. Alternatively, repeat freeze/thaw cycles can compromise the integrity of antibodies and thus antibody binding and staining intensity. The cytokeratin immunopositivity of physaliferous cells in the notochord remnant is not an unexpected finding as these cells have been shown to express cytokeratin in both human fetal notochords and chordoma neoplasms in zebrafish.15,16
The immunopositivity of chromogranin A in neural tissues, while unexpected, is not unprecedented. The previously mentioned Paquette et al. publication (2015) also remarked on a similar staining pattern in zebrafish with the same antibody derived from a rabbit host. In this study, a Western blot analysis found that chromogranin A rabbit antibody has a slightly greater molecular weight (∼44 kDa) than that of zebrafish as anticipated (∼41 kDa) based on sequence data. 8
However, chromogranin A is actually part of a larger family of chromogranins found within secretory granules in endocrine, neuroendocrine, and neural tissues. 17 Moreover, chromogranin A is expressed in the brain during zebrafish embryogenesis, possibly playing a role in neuronal development and, in rodents, is particularly localized in dendrites of certain areas of the central nervous system as well as within neuronal cell bodies of GABAergic neurons.18,19
Traditionally, S100 is considered a marker of cells stemming from the neural crest (i.e., schwann cells, melanocytes, glial cells). However, this class of calcium-binding proteins can be normally expressed by other non-neural tissues. In zebrafish, S100 protein expression has been demonstrated in keratinocytes, neuromasts, hair cells, ionocytes of gills, and olfactory epithelium either via mRNA detection through reverse-transcriptase polymerase chain reaction (RT-PCR) or in whole-mount situ hybridization. 20 This explains the non-neural S100 staining observed in this study.
The periventricular S100 staining likely represents radial glial cells, the primary neural progenitor cell in the zebrafish central nervous system, while the relative paucity of intra-neuropil glial cells can be explained by the lack of stellate astrocytes in zebrafish. 21 Radial glia also play a role in blood–brain barrier development and function, which would explain their presence near vascular/filtering structures (i.e., meninges and ventricles) observed in this study. 22
This unique distribution of glial cells in the zebrafish brain also explains the staining patterns of GFAP noted in this study. The aforementioned radial glia also express GFAP, accounting for the paraventricular staining in the brain as well as the distribution pattern within the spinal cord. Moreover, the lack of stellate astrocytes in the brain also aids in explaining the lack of GFAP staining in the neuropil.
We were unable to get the vimentin antibody to work in zebrafish FFPE tissue. We suspect that this was due to the low homology of human vimentin, used as the antigen in the generation of this antibody, and zebrafish vimentin (72% identical amino acids) based on peptide BLAST search using the Dako antibody protein ID P48616 and the zebrafish vimentin reference sequence F1QAM8.
The primary objective of this study was to assess the effects of types of fixative and duration of fixation on IHC staining in FFPE zebrafish. Some antibodies exhibited time-dependent variation in staining character. Such differences were not identified for other antibodies, potentially due to interobserver variation. Interestingly, we found that for “neural” markers, Dietrich's fixative solution greatly reduced IHC staining in our samples. While we cannot decisively recommend a fixation protocol based on our findings, we do recommend NBF followed by CalExII in neural IHC studies on FFPE zebrafish using the antibodies examined in this study. Given that an inverse relationship was found between time in fixative and staining intensity for some antibodies, a shorter fixation duration may be prudent to obtain the most accurate results when dealing with IHC.
A secondary objective was to create a panel of IHC antibodies to use in zebrafish for both research and diagnostic use. Here, we successfully determined the efficacy of seven antibodies to use for IHC in FFPE zebrafish samples. In addition, we described and expounded their staining patterns in FFPE of a species increasingly used in research.
Footnotes
Acknowledgment
This study was funded by an internal development grant from the Department of Biomedical Sciences at Oregon State University Carlson College of Veterinary Medicine.
Disclosure Statement
No competing financial interests exist.