
Abstract
Abstract
In this study, zebrafish follicles containing stage III oocytes were isolated and incubated in vitro to allow oocyte maturation. A minicell containing the intact oocyte spindle was excised after proteolytic digestion of the micropyle area of the follicle. The minicell was subjected to hypotonic treatment in deionized water for 10 min, and then spread on glass slide for karyotyping. Dyads at metaphase II and sister chromatids at anaphase II are clearly shown. Also, cytochalasin B treatment inhibited chromosomal segregation. This simple and inexpensive chromosome preparation protocol could be readily applicable to other fish species.
Karyotyping oocyte can detect changes in number of chromosomes, structure, and chromosome-attached proteins, which may be associated with infertility, developmental delay, and abnormalities of sexual differentiation and development. Zebrafish has already been an extensively studied vertebrate model organism in developmental biology, reproductive biology, and stem cells. No protocol for karyotyping zebrafish oocytes has yet been developed. The large volume of the gamete rich in yolk materials makes it difficult to apply chromosome spread methods developed in the much smaller mammalian oocytes.
In this study, zebrafish follicles at the size of 0.65 to 0.7 mm in diameter were isolated and cultured in vitro in oocyte maturation medium containing 90% Leibovitz L-15 medium (L-15) supplemented with 5 mg/mL bovine serum albumin (BSA) (V900933; Sigma), 1 μg/mL 17α,20β-dihydroxy-4-pregnen-3-one (DHP), pH 9.0, in a 35-mm culture dish at 28°C for 3 h.
1
The follicle-enclosed oocytes underwent germinal vesicle breakdown, visualized after ooplasmic clearing resulted from proteolytic cleavage of vitellogenin (Supplementary Fig. S1). At 4.5 h after the initiation of oocyte maturation, a glass needle with an inner diameter of 50 μm, filled with the crystallized protease, was placed over the micropyle region of the follicle. The setup was transferred into 90% Leibovitz L-15 medium (L-15) containing 5 mg/mL BSA and 5 μg/mL cytochalasin B (CCB, A606580; Sangon Biotech) and incubated at 25°C for 5 min. Positive pressure was applied to the needle using a microinjector. Three to five minutes later, the micropyle was digested, leaving a small hole with a diameter of 60 μm. The ooplasmic membrane under the micropyle started to extrude through the hole and formed a minicell including the spindle. The minicell was excised using a mouth-operated pipette, followed immediately by transferring into a 40 μL water droplet containing 1 mg/mL BSA (hypotonic solution) (Supplementary Fig. S2). Ten minutes later, the minicell was carefully transferred onto a glass slide prewet with fixative, 1% paraformaldehyde (PFA) (V900894; Sigma) in water, pH 9.2, containing 0.15% Triton X-100 and 3 mM
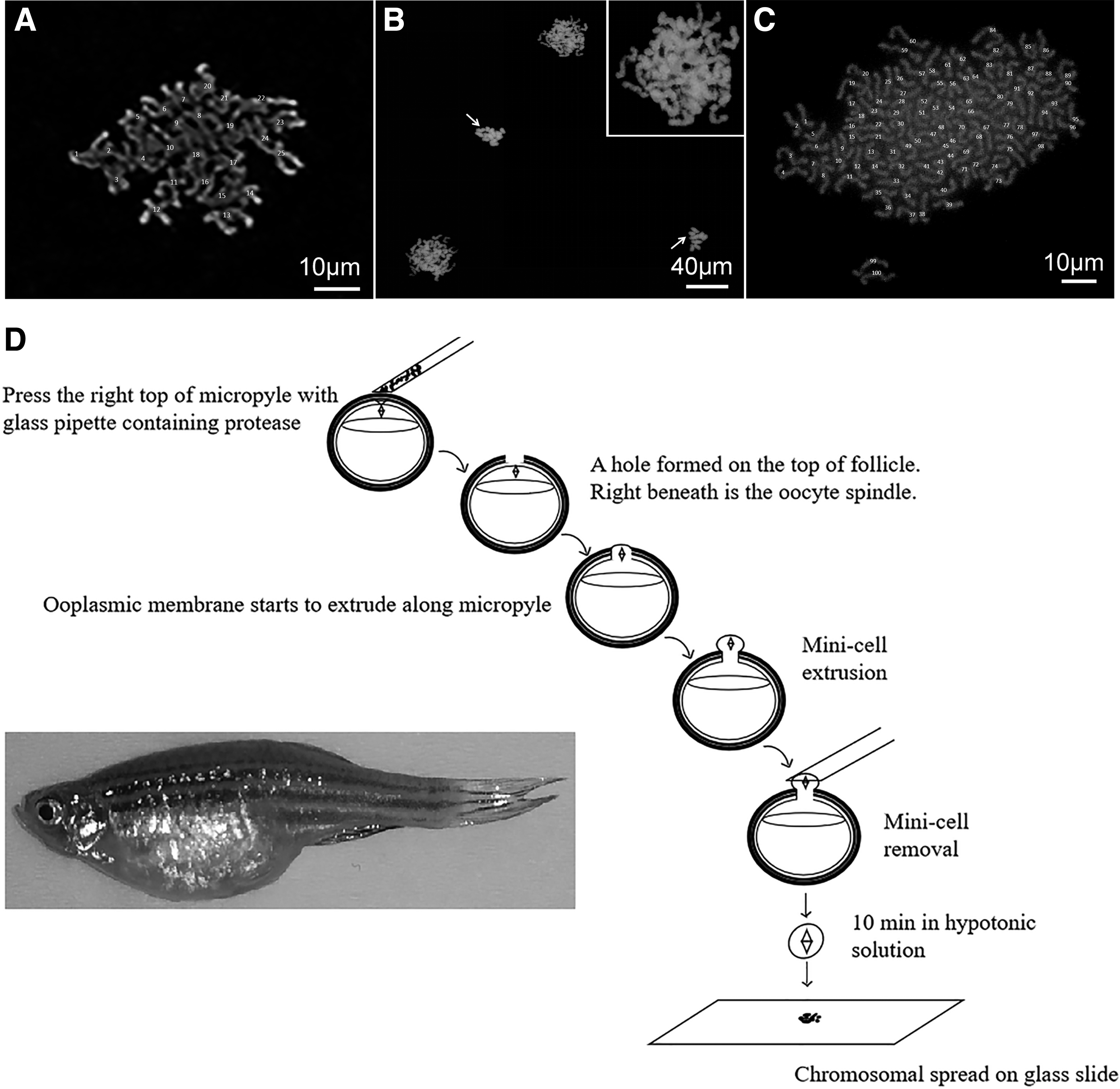
Karyotyping zebrafish oocytes at metaphase II and anaphase II.
To our knowledge, this is the first report of a method for karyotyping oocytes in zebrafish, and indeed in teleost. Two critical steps in this method are (1) we loaded crystallized protease in the microinjector needle to ensure highly localized protease activity in the area of micropyle and (2) we excised a minicell containing the spindle apparatus from the host oocyte. The minicell approach to karyotype large oocytes was first developed by the authors in Xenopus oocytes. 2 This method could be used for other fish oocyte karyotyping. In addition to karyotyping, the samples can be further used to analyze chromosome-associated proteins, as we have done in our previous study. 2
Footnotes
Authors' Contributions
H.S. conceived and designed the study. G.H., R.L., J.X., T.D., and H.S. conducted the experiments. H.S. and R.L. analyzed the data, and wrote the article with assistance from G.H. All authors read and approved the final article.
Acknowledgment
This study was supported by Guangdong province's Yangfan Program 2016 to H.S.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.