
Abstract
The zebrafish is a widely used animal model in biomedical research. Despite this, pathogens continue to be common in laboratory zebrafish. It is important to determine and describe the pathophysiology of cryptic infections on zebrafish to elucidate the impacts on experimental endpoints. Body condition is a basic measurement used experimentally and in health monitoring of animals. We exposed three wild-type zebrafish strains: AB, WIK, and 5D to Pseudoloma neurophilia. After 8 weeks postexposure, we individually imaged and processed fish for histology. Morphometric analysis was performed on images and an index of body condition was calculated based on the ratio of length/width from the dorsal aspect. Histological sections from each fish were examined to establish sex, severity of infection, and tissue distribution. We observed a significant decrease in body condition in female fish that was not observed in males. In addition, we observed a negative correlation between the total gonadal area of P. neurophilia exposed females and unexposed controls. These results illustrate the sex-specific impacts of a common chronic pathogen on zebrafish health and a commonly used experimental endpoint, further supporting the establishment of rigorous health monitoring programs in laboratory zebrafish colonies that include screening for chronic infectious diseases.
Introduction
The use of zebrafish as a model vertebrate has grown dramatically over the last 20 years. Originally, in a model for developmental genetics, the use of zebrafish in research has expanded to include several diverse fields such as infectious disease, immunology, cancer, behavior, and neuroscience. Cryptic infections of laboratory zebrafish continue to be a potential confounder in experiments. These infections can significantly hamper the reproducibility and translational potential of zebrafish research, particularly as experimental designs become more sophisticated and allow for increasingly sensitive measurements of physiological, behavioral, and genetic endpoint measurements.
The importance of chronic infectious diseases in research animals has been recognized since the early days of the laboratory mouse (see Review 1 ). However, as the zebrafish is a less mature research model, this recognition has come rather recently. 2 In fact, many researchers continue to obtain fish for use in research from local pet stores and wholesale suppliers, in which little to no biosecurity exists and multiple fish species cohabitate.
There are several well-described chronic pathogens of zebrafish. The most prevalent ones in zebrafish facilities include several Mycobacterium species, Pseudoloma neurophilia, and the nematode Pseudocapillaria tomentosa. 3 The microsporidian parasite, P. neurophilia, as the name suggests, infects primarily neural tissue.4,5 It has been demonstrated to infect a broad range of tropical fish species 6 and is one of the most common chronic infections in laboratory zebrafish. The parasite has been found in about half of the zebrafish research facilities based on surveys 3 and the database from the diagnostic service of the Zebrafish International Resource Center (http://zebrafish.org/health/). In general, P. neurophilia infections appear to be relatively benign. However, it has been shown to result in reduced fecundity, 7 increased mortality, especially in immune suppressed animals, 8 and has a measurable impact on the endpoints studied in behavioral research.9,10
While many fish infected with P. neurophilia appear normal, it has been associated with emaciation since it was first described from fish in research laboratories. 4 Body condition scoring (BCS) is a simple, visual method for health assessment of animals. Methods for BCS are generally semiquantitative and used to assess the welfare of animals used in research settings to determine morbidity. Clark et al. 11 provide landmark-based methods for BCS, which includes examination of fish from either side or from above. It is important to link these changes with more detailed clinical and pathologic changes in zebrafish.
Several behavioral assessment tools for zebrafish depend on visualization and morphometric analysis of adult fish. Zebrafish vary greatly in size between males and females, and it is possible that there is a differential effect of the infection on body size and condition factor based on sex. In this study, we evaluated the effects of infection on BCS by quantitative assessment using an imaging system with morphometric analysis of zebrafish from the dorsal aspect, recording strain, and sex of individual fish. We experimentally exposed three commonly used wild-type strains of zebrafish to P. neurophilia and assessed BCS of individual fish using morphometric image analysis. Histological endpoints of P. neurophilia infection were scored as described by Spagnoli et al., 12 and total ovarian area was measured.
Materials and Methods
Fish
Three wild-type strains of zebrafish (5D, AB, and WIK) maintained at the Champalimaud Center of the Unknown (CCU) Fish Platform were used in the study. Fish were maintained on a 14 h light-10 h dark cycle throughout the study and housed at a density of 10 fish/L. For P. neurophilia exposures, fish were transferred from the pathogen-free main room to the “high risk” quarantine, as described by Martins et al. 13 Controls were held in the main room. Water quality parameters in the two rooms within the facility were very similar. 13 For example, temperatures (28°C) were essentially identical, water exchange was 5% for the main facility and 10% for quarantine, and conductivity was 1290 versus 1693, respectively.
Experimental fish were fed twice daily with Gemma 500 diet (Skretting®, Spain). Fish were sexually mature and ranged in age 5–10 months at the beginning of the experiment, depending on fish line. Date of fertilization of each egg population was as follows: WIK, June 1, 2016; AB, October 20, 2016; and 5D, June 15, 2016. Regarding background infections, fish in the pathogen-free main room are screened multiple times a year for the presence of DNA from several bacterial and parasitic species, including P. neurophilia and P. tomentosa, by polymerase chain reaction by IDEXX (https://www.idexxbioanalytics.com/zebrafish-health-monitoring). All live animal studies were conducted at the Champalimaud Centre and all zebrafish researchers and platform staff were accredited as animal experimentation users according to the FELASA guidelines.
Exposure
After 1 month of acclimation to the quarantine room, each line was randomly exposed using carcasses of known infected fish provided by J.L.S. and M.L.K. from their flow-through facility located at Oregon State University. Approximately 30 fish were euthanized by overdose of MS-222 and sent on ice to the CCU Fish Platform. The viscera were removed and muscle, with spinal cord, and ovaries from females were minced and divided equally among the three tanks. Hence, fish were exposed by ingesting infected host tissue and possibly free spores. Water flow was reduced for 24 h and then resumed.
Examination
After ∼8 weeks postexposure to the parasite, both exposed and control unexposed fish from each tank were examined. Images with a resolution of 980 × 980 pixels were obtained from individual fish by placing them in a circular arena (diameter 45 cm, water depth 2.5 cm) used for behavior studies with a camera (Basler A622f, Zeiss 16 mm-objective) placed above ∼1.2 m from the water surface. 14 To quantify body condition, morphometric analysis was performed by measuring the length and width of each fish using a custom MATLAB script. Length (standard length) was determined from the tip of the snout to the base of the tail (i.e., end of the caudal peduncle) and the sides of each fish were marked in frames of the respective reference video, in which the animal showed a straight body posture. The script then calculated the length and width as the distance (in pixels) between the marked points. An index describing the overall thickness of each fish was calculated by dividing total length by body width.
After imaging, individual fish were euthanized by inducing rapid hypothermia in ice water,15,16 and then preserved individually in Dietrich's preservative. Fish were transferred to 50% ethanol after 72 h and sent to Oregon State University, where they were decalcified with Cal-Ex II for 24 h and processed for histology at the Oregon Veterinary Diagnostic Laboratory. They were then sectioned longitudinally by dividing the fish in half along the midline, both halves were processed and two slides were prepared, separated by 25 μm to optimize visualization of the spinal cord. This resulted in four separate sections/fish. All slides were stained with hematoxylin and eosin, and selected slides with inflammatory lesions were stained with Luna and acid fast 17 stains to visualize either spores of P. neurophilia or acid fast bacteria (i.e., mycobacteria).
Presence and severity of P. neurophilia and associated lesions were scored as follows: total number of aggregates of spores in the spinal cord, hind brain, and nerve roots, presence and severity of meninxitis as described by Spagnoli et al., 12 and presence of multifocal, chronic myositis previously associated with P. neurophilia infections. The total area of ovary was measured from all four sections at 25 × magnification using a SPOT RT® color camera with image capture (Diagnostic Instruments).
All statistical analyses were performed using GraphPad Prism (version 8.0.2). The mean length/width within each tank was compared between exposed and unexposed fish separated by sex using unpaired t-tests with Welch's correction for unequal variances. The relationship between ovary area and length/width was assessed by calculating the Spearman nonparametric correlation coefficient.
There were undetected infections within each tank of exposed fish. Hence, we compared the infection status in females with our two important fitness endpoints; length/width ratio and gonad area. Fish were identified as infected if they exhibited parasites or lesions that are essentially pathognomonic for the infection (e.g., myositis, meninxitis, etc.). All the data, except gonad area for the AB strain, were normally distributed, and hence, data were analyzed using a standard t-test. For the AB strains, gonad areas between infection detection were compared using a Mann–Whitney–Wilcoxon test. Significance was set at p < 0.05.
A total of four exposed fish were removed from the analysis as they were observed by histology to have lesions associated with other diseases: two females WIK, one male 5D, and one female 5D with ectopic testis.
Results
The overall prevalence of P. neurophilia observed in the exposed fish by histology was 45% (Table 1 and Figs. 1 and 2). Fish were randomly assigned to treatment groups; however, there was a clear female bias observed among all groups. This female bias has been previously documented in the fish population at this facility. 3 No parasites were observed in control fish. Typical of the infection, aggregates of spores were observed along the spinal cord, particularly in the myelinated fibers and nerve roots. Enumeration of parasite aggregates showed a range from 0 to as high as 36, with an overall mean of 6/fish.

Pseudoloma neurophilia infections in experimentally exposed zebrafish. Hematoxylin and eosin.
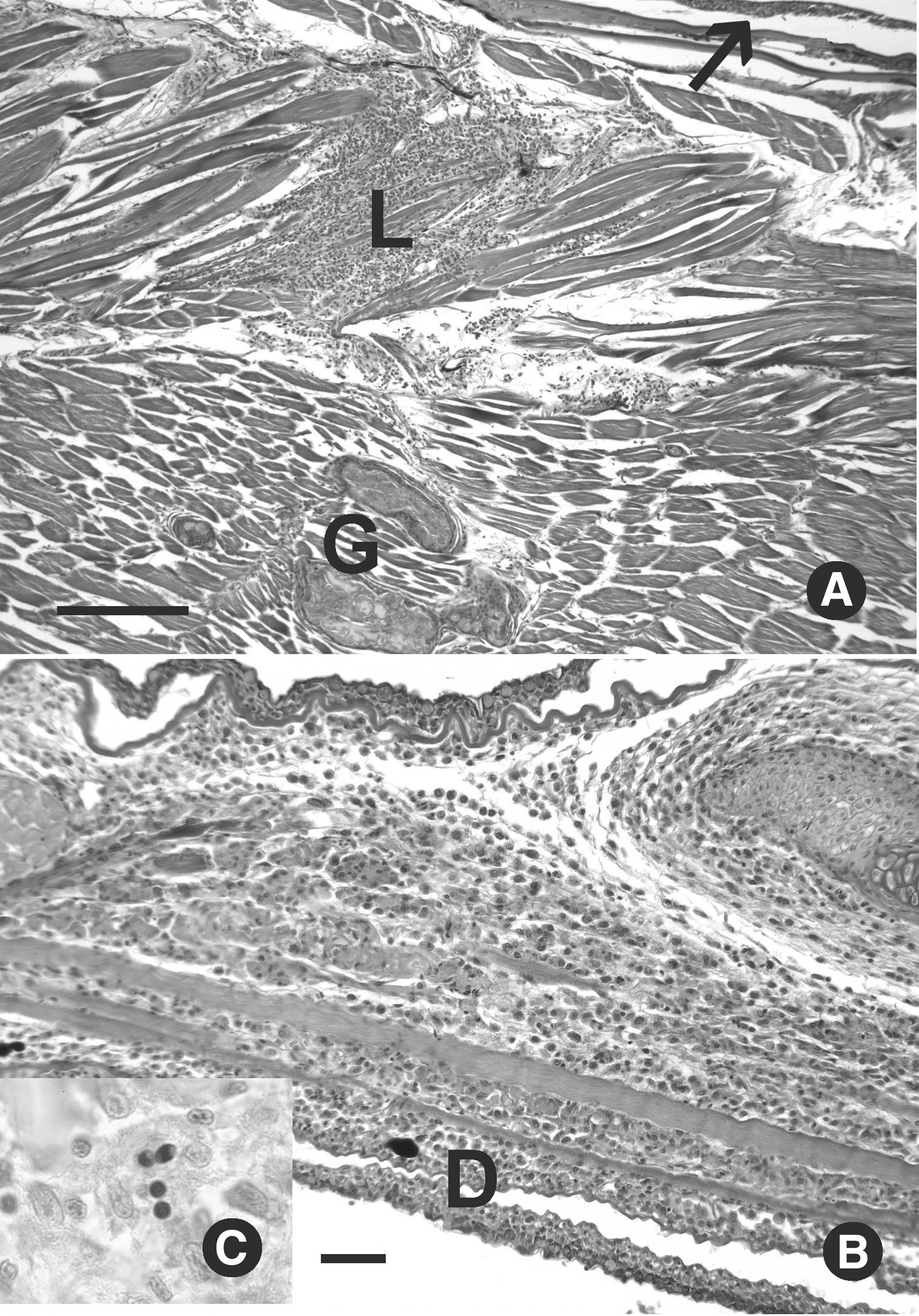
Myositis associated with Pseudoloma neurophilia infection.
Pseudoloma neurophilia Infections in Exposed WIK, AB, and 5D Zebrafish Lines Examined by Histology After 8 Weeks Exposure
Prevalence (% infected within the exposed groups) includes fish with either myositis or meninxitis and no visible aggregates in the spinal cord or nerve roots. Aggregate intensity, no. of clusters of spores observed in the central nervous system.
Meninxitis and myositis were observed in 24% and 11% of fish, respectively (Table 1 and Fig. 2). Myositis was characterized by multifocal regions of chronic inflammation and myodegeneration (Fig. 2). Some of the lesions extended into the dermis, and one AB female had prominent inflammatory lesions throughout the lower jaw (Fig. 2B). Two of the five fish with myositis exhibited multiple granulomas in the somatic muscle with degenerative eosinophilic debris consistent with lysed muscle fibers (Fig. 2). Luna and acid fast staining revealed spores within these inflammatory lesions, with no evidence of acid-fast bacteria (Fig. 1D). No mortality occurred in either exposed or control fish over the course of the exposure.
Significant lesions that were not associated with the infection were as follows: a large proliferative lesion in the pancreas consistent with a poorly differentiated carcinoma in one 5D male (Fig. 3A), nematodes in the intestinal epithelium that were consistent with P. tomentosa in two WIK females (Fig. 3B), and ectopic testis observed in the pancreas of one 5D female (Fig. 3C), while the ovary of this fish appeared normal. Of note, the ovary of one of the fish with the nematode infection was also very atrophied. All four of these fish were excluded from further analysis.
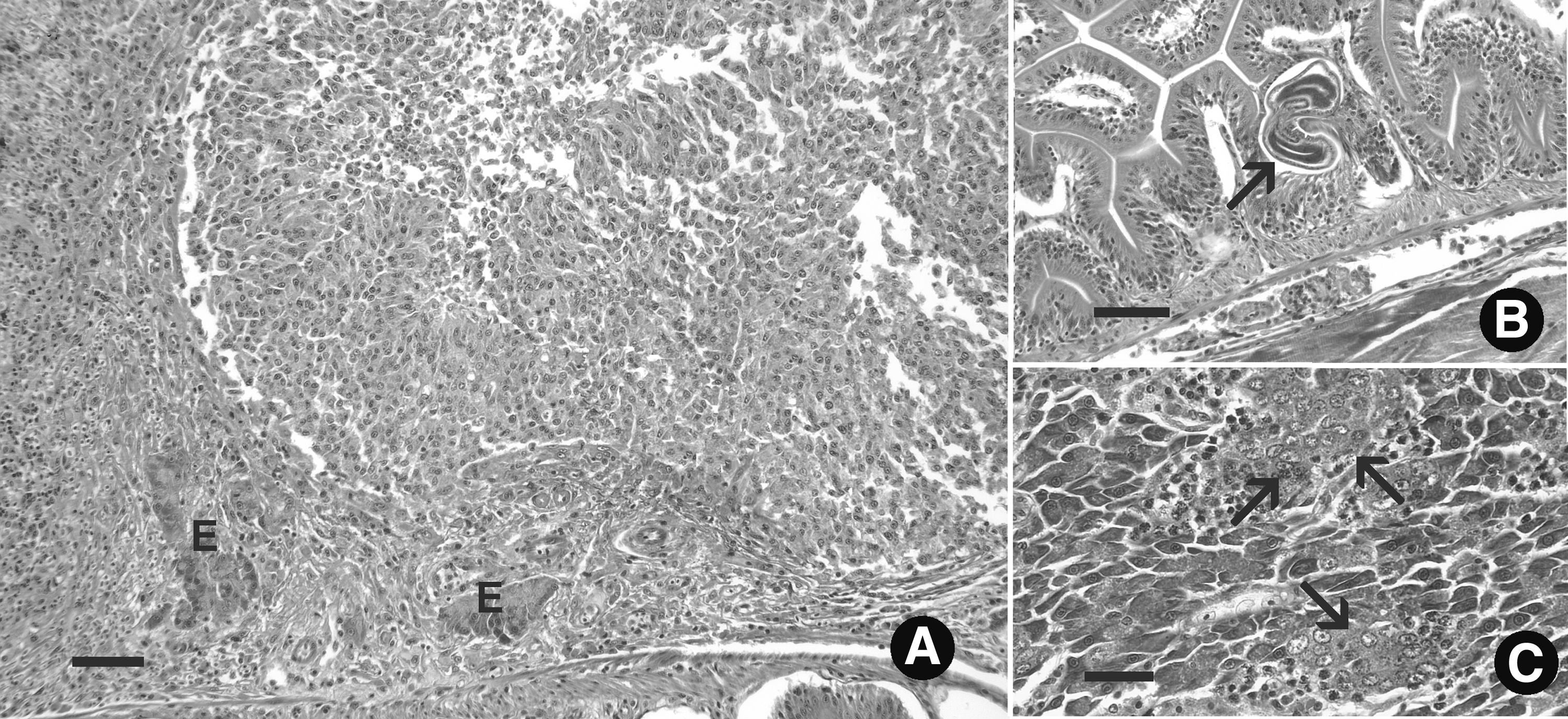
Lesions not associated with Pseudoloma neurophilia. Hematoxylin and eosin.
Ovary histology and size
The ovaries of most females from both the exposed and control groups appeared within normal limits, all stages of development and numerous, large vitellogenic eggs (Fig. 4A). There was no statistical difference between fish strains within exposure categories, and hence the three groups were evaluated together. The four females from the exposed group that had the smallest ovaries had poorly developed ovaries with no vitellogenic eggs; two ABs and two WIKs (Fig. 4B). P. neurophilia infection was detected by histology in three of these fish. The ovaries of the remaining infected fish appeared normal, except for their overall smaller size as a group.
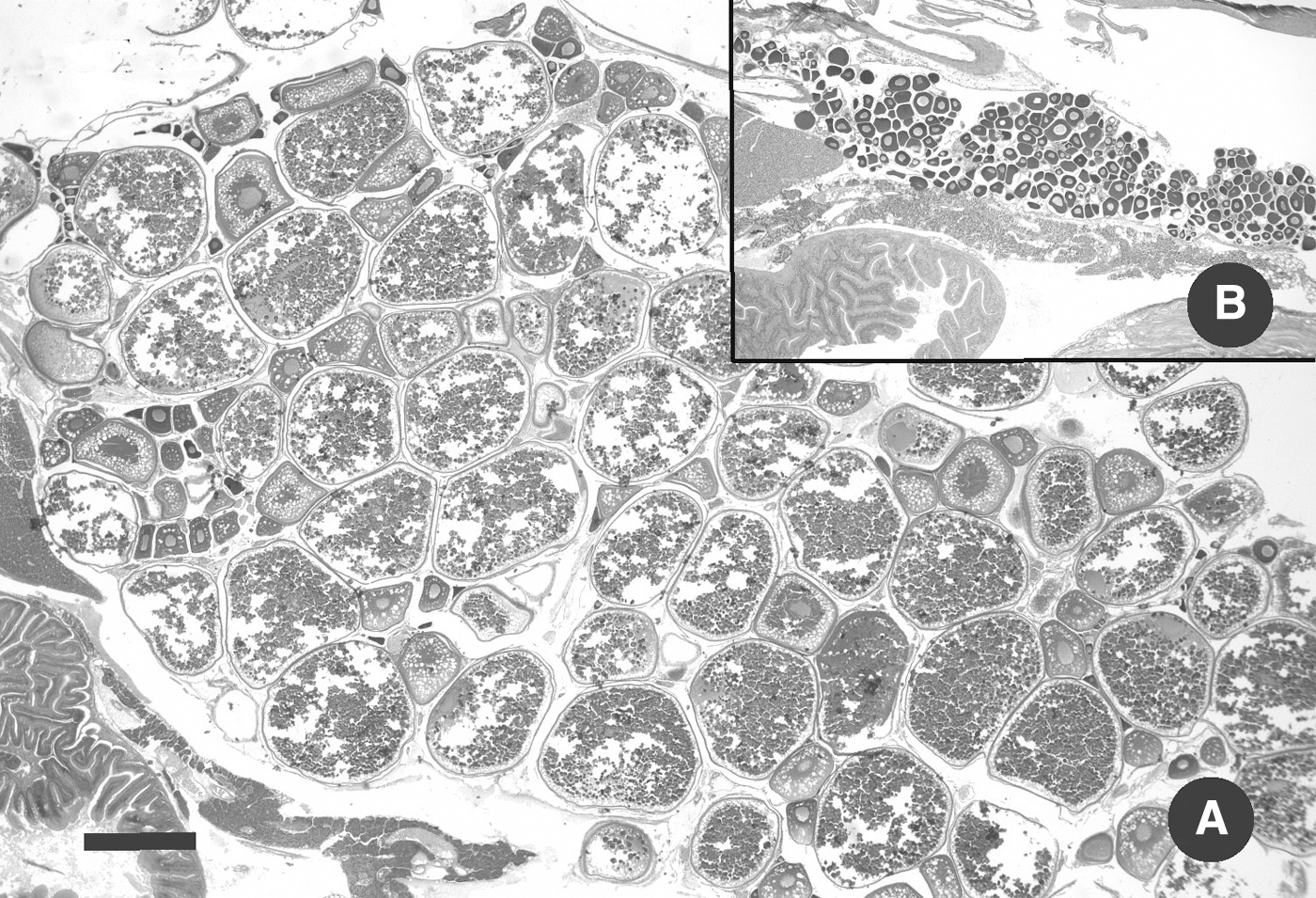
Ovaries. Hematoxylin and eosin. Scale bar = 0.5 mm.
Two control fish with large ovaries had very mild egg associated inflammation and fibroplasia, each with a small focus of eosinophilic debris. Based on measurements of four sections/fish, the range of controls was 15–109 mm2 (n = 24), while exposed fish ranged from completely immature ovaries with as low as 2 mm2 and with the maximum of 64 mm2 (n = 52). Hence, the mean of the ovarian area of the exposed fish was only 65% that of the controls.
Body condition scores
Females from exposed groups had a significantly higher length/width ratio (Figs. 5 and 6), indicating an overall decrease in body thickness (AB: 5.97, n = 16; WIK: 6.31, n = 18; 5D: 6.04, n = 20) compared to unexposed controls (AB: 5.25, n = 8; WIK: 5.43, n = 9; and 5D: 5.78, n = 9). Exposed females from the three strains combined had an average length/width ratio of 6.07 (SD = 0.36, n = 57) compared to control females with an average of 5.48 (SD = 0.40, n = 25). Negligible differences in length/width ratio were observed between exposed males (AB: 6.06, n = 7; WIK: 6.10, n = 4; and 5D: 6.08, n = 13) and control males (AB: 5.95, n = 10; WIK: 6.19, n = 9; 5D: 6.17, n = 10). Overall average length/width ratios for exposed males from the three lines combined was 6.09 (SD = 0.25, n = 25) and 6.11 (SD = 0.40, n = 28) for controls. There was a moderate negative correlation between decreased female body thickness (i.e., higher length/width ratio) and gonadal area (Fig. 7) (Spearman r = −0.44, p < 0.0001).

Dorsal view of zebrafish in arena. Image is a frame capture taken from video shot in an experimental arena containing six exposed WIK zebrafish just before euthanasia.
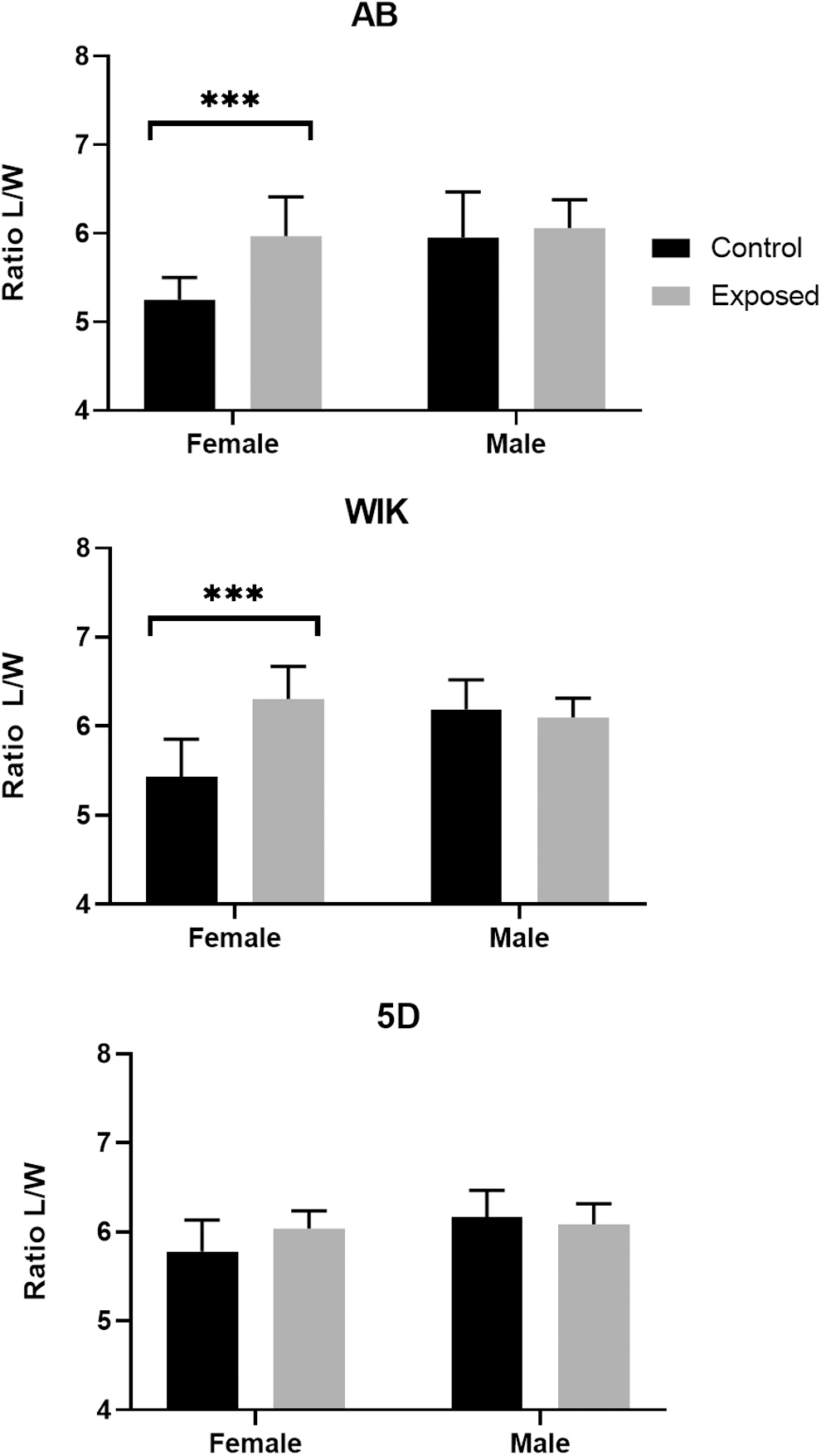
Decreased condition factor of female zebrafish infected with Pseudoloma neurophilia. The condition factor of female zebrafish was significantly decreased in AB and WIK fish as observed by an increase in the length to width ratio in exposed animals (AB: 0.72, WIK: 0.88, t-test, p < 0.0001). An increase in the length to width ratio was observed in the 5D female animals (0.26), however, this difference was not statistically significant (t-test, p = 0.67). Only minor differences in the condition factor of males in the AB, WIK, and 5D groups were observed (0.1, 0.8, and −0.09, respectively) (***, p ≤ 0.0001).
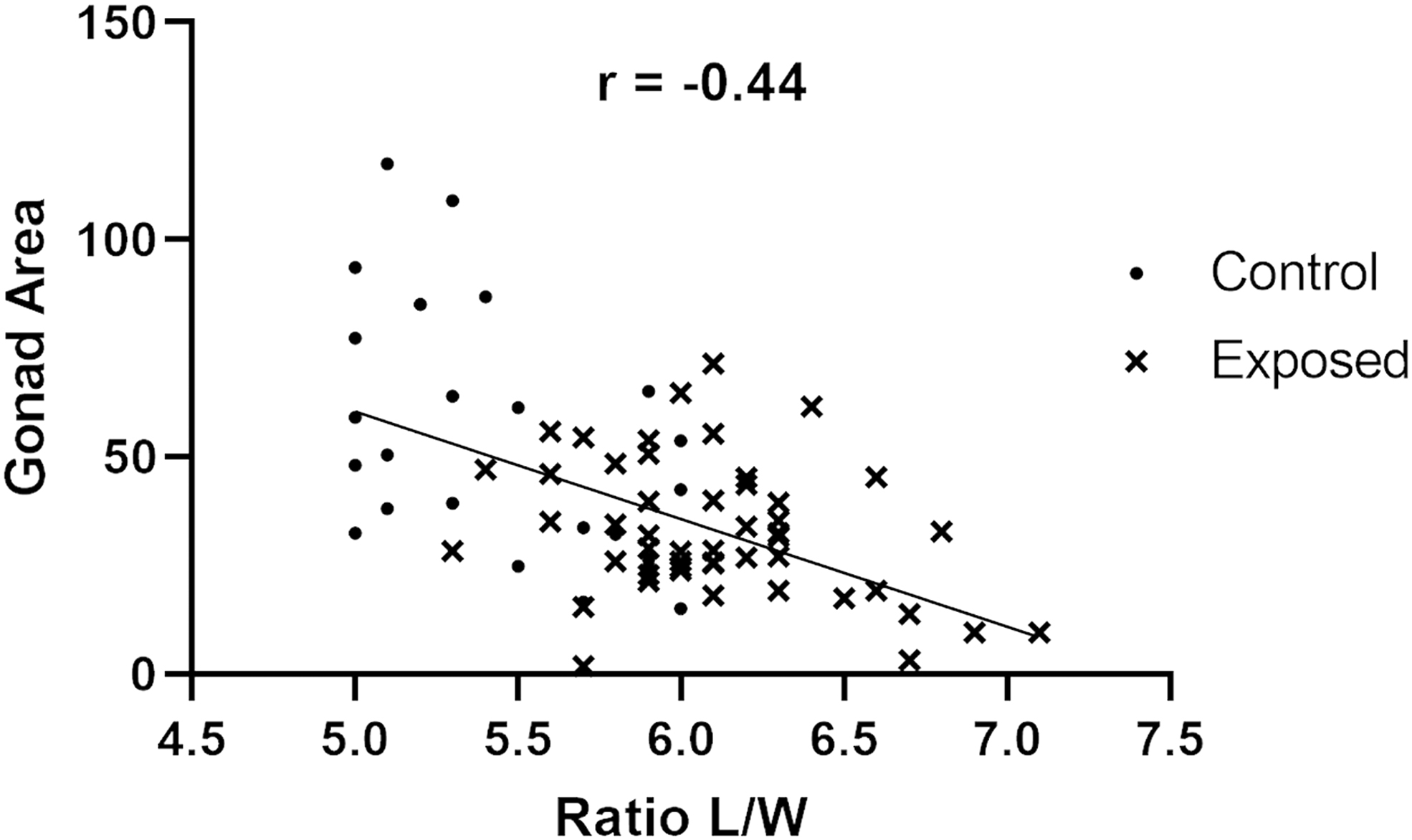
Gonad area is negatively correlated with condition factor area in female zebrafish. Total gonadal area visible on histological slides was measured in triplicate step sections from each individual and compared with the ratio of length to width of the individual measured from video analysis. For all female fish in the study, there was a negative correlation between gonadal area and body condition (Spearman r = −0.44, p < 0.0001, n = 76). Overall, reduced body condition was observed in exposed fish (
Intratank analyses
No significant difference was observed with length/width ratios for all three strains. Regarding gonad areas, fish in which the infection was detected had less gonad area than fish from the same tank in which the infection was not detected: WIK, 33.7 mm versus 27.6 mm; 5D, 43.3 mm versus 30.3 mm; and AB = 27.1 mm versus 17.9 mm. WIKs showed no significant difference (p = 0.18), whereas gonads were significantly smaller in fish with detectable infections for both the 5D (p = 0.014) and AB (p = 0.0152) groups.
Discussion
Experimental exposure of zebrafish to P. neurophilia using infected carcasses resulted in a high prevalence of infection comparable to other studies. P. neurophilia is a chronic and persistent infection, and the prevalence of infection increases with over several months following exposure.7,18 In the present study, infected carcasses were particularly effective: Ramsay et al. 7 observed only 16% infection by histology in unstressed fish exposed in a similar manner after 8 weeks, while we observed about 45% at this time period. Comparable to the present study, Kent and Bishop-Stewart 19 observed 30% infection in zebrafish at 8 weeks following exposure of fish to infected spinal cords. There is a possibility that cryptic P. neurophilia infections could be missed by histological examination. However, as multiple sections from each fish were examined by an experienced pathologist, this is unlikely.
The fish in the present study also exhibited the typical range of lesions associated with the infection, 12 including multiple aggregates of spores in the spinal cord and nerve roots, meninxitis, and myositis. Differences in occurrence of aggregates and parasite-associated lesions were observed between strains (e.g., myositis was not observed in the WIK strain). However, constraints imposed by the experimental design (e.g., one exposed tank per strain) prevent meaningful statistical analyses to possibly attribute differences in these histological observations to strain-specific variation.
In a large retrospective study of over 500 infected fish submitted as diagnostic cases to ZIRC, 12 meninxitis was observed in ∼50% of infected fish. Similarly, this inflammatory lesion around the spinal cord was observed in ∼62% of infected fish in the current study. This same survey found the presence of myositis (16%) and we found slightly less at 11%. Spagnoli et al. 12 reported that this muscle lesion was significantly greater in fish submitted to the Zebrafish International Resource Center diagnostic service with clinical disease compared to apparently healthy sentinel fish. Previous studies have associated increased myositis with infection and immunosuppression due to crowding stress 20 or exposure to gamma radiation. 8 Hence, these studies and that of West et al. 21 indicate that the presence of these lesions increased with time or poor health status.
In contrast, in this study, we observed profound myositis, including the formation of granulomas after only 8 weeks exposure. Infection of zebrafish by P. neurophilia begins in the epithelial cells of the intestine and pharynx, rapidly progressing to visceral organs. Muscle infections have been demonstrated in larval zebrafish as early as 72 h postexposure, at the same time as infection can be observed in the spinal cord and in the absence of brain infection. 22 As muscle infections are somewhat rare in subclinical fish, it is possible that the P. neurophilia-associated myositis and granuloma formation observed in these adult zebrafish are the result of transient infection related to early-stage progression of the parasite.
While many infected fish appear clinically normal 12 and other diseases cause emaciation in zebrafish, the infection is associated with increased emaciation 4 and spinal deformities. 5 Ramsay et al. 7 reported reduced size and condition factor (length/weight) in experimentally infected TL and AB lines of zebrafish. With the former, infected fish weighed less and had reduced condition factor at 10 weeks postexposure and exposed fish weighed 27% less overall. While not evaluated by sex, subjective observation indicated that control females had fuller bodies than exposed females. 7 With the AB line, control fish were significantly larger than exposed fish at 20 weeks postexposure. Consistent with Ramsay et al., 7 we observed significantly reduced body scores in females, but not males, in two of the three lines.
While the body score was decreased in the third line (5D), this difference was not significant, likely as a result of female control fish of this line being generally thinner than fish from the other two lines. This could be due to differences between development rates between the strains (i.e., a characteristic phenotype of the 5D strain) or an effect isolated to this particular clutch of fish. The trend of reduced body scores in females occurred with all three tanks (strains), but was not statistically significant with the 5D group.
Rearing conditions were very similar between tanks, but given that there were no tank replicates for exposed or control fish within lines and fish were held on physically separate systems, we could not exclude the possibility of cryptic tank effects causing differences in body scores and gonad size. However, all exposed tanks contained some fish, in which neither infection nor lesions consistent with the infection were detected, and this allowed for evaluations of effects on infection status with our fitness endpoints within tanks. With all three lines of exposed female fish, those with detected infections had less gonadal area than fish in which no infection was observed, and this was statistically significant for the 5D and AB tanks.
In a retrospective study of the ZIRC database, evaluating 1100 fish, Chow et al. 23 found that males without clinical disease had a significantly higher prevalence of infection than females, but this was not the case for those submitted with disease signs. Therefore, our data, supported by Ramsay et al. 7 and Chow et al., 23 suggest that infections may be more common in males, but females show greater reduction in body scores, and are therefore easier to diagnose. This difference is likely due to effects on ovarian development. Spagnoli et al. 12 found that fish submitted as having clinical disease tended to have more myositis, but not higher numbers of parasite aggregates in the spinal cord. This agreed with our findings, in that although we observed differences in body condition comparing females from exposed with control groups, we did not see direct correlations with lesion or parasite scores. That study also found that clinical fish did not have significantly different parasite scores than those submitted as clinically normal. That retrospective study involved hundreds of infected fish, and our experimental data support this finding.
Even with multiple sections, histology probably misses light infections or damage caused by transient infections, which provides an explanation for the strongest differences in body scores occurring by simply comparing exposed versus unexposed populations. This is similar to results observed while studying the effects of the parasites on behavior.9,12
Sexually mature female zebrafish tend to be larger than males, which is particularly evident in females with mature, large ovaries. 24 We observed a negative correlation between body scores, ovary mass, and development that is, females with large ovaries containing numerous mature, vitellogenic eggs had thicker bodies, and thus lower length to width ratios, while the thinnest fish (i.e., some fish within the exposed groups) had very small ovaries with only immature eggs and reduced body scores. Only one infected female was somewhat of an outlier, with a small ovary (1.8 mm2) but a low length to width ratio (5.7). This fish had significant myositis, with lesions that extended through the dermis, and it is possible that edematous changes resulted in a wider body. Although Ramsay et al. 7 did not examine ovaries, they found that infected fish, particularly those subjected to stressors, had reduced egg numbers during spawning associated with the infection, which agrees with our observations.
Several factors may influence the fecundity of female zebrafish infected with P. neurophilia. It has been previously observed that P. neurophilia is present in high numbers in the ovaries and can be shed during spawning and transmitted vertically. 25 However, neither spores nor inflammatory changes were observed even during close examination of immature ovaries. The parasite targets the nervous system, and thus it is not surprising that it causes behavioral changes.9,10 Neural alterations may extend to gonadal development and breeding. Stahl et al. 26 reported severe atrophy with immature follicles in mice chronically infected with the neural parasite, Toxoplasma gondii, with no infection observed in the ovaries and uteri. They observed a wasting syndrome in infected mice and a failure to litter successfully even before signs of wasting. This disruption of the reproductive process was attributed to hypothalamic dysfunction caused by edematous changes and focal disruptions within the thalamus and hypothalamus.
The hypothalamo-pituitary-gonadal axis appears to be highly conserved between zebrafish and mammals. 27 Thus, disruption of the endocrine signaling of the hypothalamus, either directly associated with damage caused by a parasite or by inflammatory responses related to infection could result in the acute wasting of gonadal tissue observed in P. neurophilia-infected female zebrafish. In a comprehensive histological examination of P. neurophilia infections, Spagnoli et al. 12 described high numbers of the parasite primarily localized within axons composing the ventral white matter of the spinal cord and the hindbrain with few parasite clusters in gray matter. Nonetheless, necrosis of neuron cell bodies in the gray matter of the hindbrain was observed in areas distant from white matter infected with P. neurophilia, suggesting that infection of axons by the parasite can affect distal cell bodies. The wasting and arrested ovarian development observed here could be explained by either destruction of cells within the thalamus or hypothalamus by effects from distal axons infected with P. neurophilia or by disruption of signaling from the hypothalamus or pituitary caused by damage to neurons responsible for conducting signals from these organs to the ovaries.
In conclusion, we established chronic infections of P. neurophilia in three commonly used wild-type zebrafish lines and quantified body condition using an imaging system and morphometric analysis of fish from the dorsal aspect. We observed a decrease in condition factor specifically associated with infected female fish. Analysis of histological endpoints related this decrease in condition factor to an ovarian wasting syndrome without apparent parasite infection of the ovarian tissues, suggesting a possible neurological etiology. Future studies to establish a causal mechanism behind this phenomenon are planned. The data presented here provide a quantitative basis to validate less labor intensive, observation-based methods such as that described by Clark et al. 11 that can be routinely used by laboratory zebrafish colony staff.
Footnotes
Acknowledgments
The authors wish to thank Robert Hinz, Francisco Romero-Ferrero, and Gonzalo G. de Polavieja from the Collective Behavior Laboratory at Champalimaud for assistance with image acquisition and analysis for this study.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was partially funded by the Champalimaud Foundation and developed with support from the research infrastructure Congento, cofinanced by Lisboa Regional Operational Programme (Lisboa2020), under the PORTUGAL 2020 Partnership Agreement, through the European Regional Development Fund (ERDF) and Fundação para a Ciência e Tecnologia (Portugal) under the project LISBOA-01-0145-FEDER-022170. This project was also funded, in part, by the NIH ORIP R24OD010998 to M.L.K. Travel for M.L.K. was provided by the Department of Biomedical Sciences, Oregon State University through their “mini-sabbatical” program.