
Abstract
Hoplias malabaricus is one of the most widespread freshwater fish in the Neotropics; all karyotyped populations exhibit diploid numbers ranging from 2n = 39 to 2n = 42, with variations that make up to seven karyomorphs, suggesting the existence of a species complex or paraspecies. In this study, we explored the diversity of H. malabaricus of the Magdalena River Basin using karyotypic analysis. Cytogenetic analyses were carried out on 32 specimens and six sample sites from the upper, middle, and lower Magdalena River, a trans-Andean basin in Colombia, using standard and molecular techniques and fluorescence in situ hybridization (FISH). The upper and middle Magdalena samples were 2n = 42 and showed unique fluorescence patterns, whereas the lower Magdalena samples had 2n = 40, generalized fluorescence patterns, and one specimen with a mosaic karyotype and heteromorphic chromosome pair. The genetic discontinuity along the Magdalena River fits the museum hypothesis, where the 2n = 42A predates the modern configuration of the Magdalena Basin in the Late Miocene, and the 2n = 40C karyomorph represents a more derived condition. Our results underline the complex biogeography of the trans-Andean region, where the Magdalena-Cauca Basin should be considered a freshwater fauna mosaic with divergent evolutionary affinities.
Introduction
H
Cytogenetic,4–6 phylogeographic,7–9 and DNA barcoding10–12 studies on cis-Andean H. malabaricus populations have uncovered high levels of cryptic diversity. Cytogenetic data indicate the existence of seven karyomorphs (A–G) that differ in their diploid numbers, chromosome morphology, and in some cases, several sex chromosome systems4,7,13; occasionally, some karyomorphs hybridize. 14 In the last years, molecular cytogenetic studies have been extensively performed in several H. malabaricus karyomorphs.15,16
All evidence indicates H. malabaricus is a species complex 4 or paraspecies (sensu Albert et al. 17 ) due to the existence of several molecular independent lineages and karyomorphs, some of them occurring in sympatry.4,9,12 Due to its low vagility, this taxon represents an excellent model for the study of allopatric, 18 parapatric, and sympatric 19 conditions, biogeographical analyses, 9 and studies focusing on survival strategies.
The Neotropical freshwater fish fauna is unique, because it is the largest in the world, and most of its diversity is at low-level taxic differentiation. 20 Since the late Cretaceous and during the Cenozoic, the Andes orogeny determined the evolution and diversification of Neotropical fish.21,22 The Andes uprising and the foreland basin caused drastic ecological transformations in the local fish fauna, determining the extinction of fish that characterized older drainage systems. 21 In each drainage, the faunal composition is a result of extinction, speciation, vicariance, and dispersal. 23
In Northern South America (NSA), the Magdalena-Cauca River Basin is the largest trans-Andean basin with 1612 km and a minimum age of ∼10 Ma due to the final uplift of the Eastern Cordillera. 21 The maximum elevation of this basin is of 3685 m above the sea level. 24 It harbors 222 fish species, which represent roughly 15% of the Colombian ichthyofauna. 25 Such richness is considerably lower than what is observed in other geographically close drainages, such as the Orinoco and the Amazon Basins, which harbor ∼1000 and 2000 fish species, respectively 17 ; nonetheless, almost 50% of the freshwater fish in the Magdalena-Cauca are endemic.
Biodiversity loss is an old trend for the Magdalena-Cauca drainage; paleontological data show that this basin lost at least four species typical of the Amazon and Orinocan region: the lungfish Lepidosiren, Colossoma, Brachyplatystoma, and Arapaima.26,27 Therefore, the Magdalena-Cauca Basin is an intriguing scenario for studying the effects of the Andean orogeny and its consequences for speciation, extinction, and immigration of Neotropical freshwater fish.
In Colombia, H. malabaricus occurs in all hydrological basins: Amazon, Orinoco, Magdalena-Cauca, Pacific, and Caribbean 25 ; interestingly, this species is absent in the upper Cauca River.28–30 According to Rincon-Sandoval et al., H. malabaricus exhibits two well-structured lineages, and samples from the Magdalena River Basin form a clade, including Rancheria, Catatumbo, and Tiura Basins. 31 However, the genetic structure of H. malabaricus along the Magdalena Basin is entirely unknown.
Cytogenetic studies on freshwater fish are scanty and restricted to a few endemic species 32 in Colombia. Here we investigated the cytogenetic diversity of H. malabaricus from the Magdalena River Basin. Our results indicate an apparent parapatric condition, with conspicuous differentiation between upper Magdalena (UM), middle Magdalena (MM) regions, and lower Magdalena (LM), with minor differentiation between the UM and MM regions using conventional and molecular fluorescence in situ hybridization (FISH) techniques.
Materials and Methods
We carried out cytogenetic analysis on 32 specimens (10 from UM, nine from MM, and 13 from LM; Table 1 and Fig. 1.), samples were collected on live specimens and cell division was stimulated with yeast 24 h before processing 33 ; fish were euthanized with clove oil. 34 Voucher specimens were deposited in the ichthyological collection of the Pontificia Universidad Javeriana in Colombia (MPUJ). Specimen identification followed Oyakawa, 1 and Oyakawa & Mattox. 35
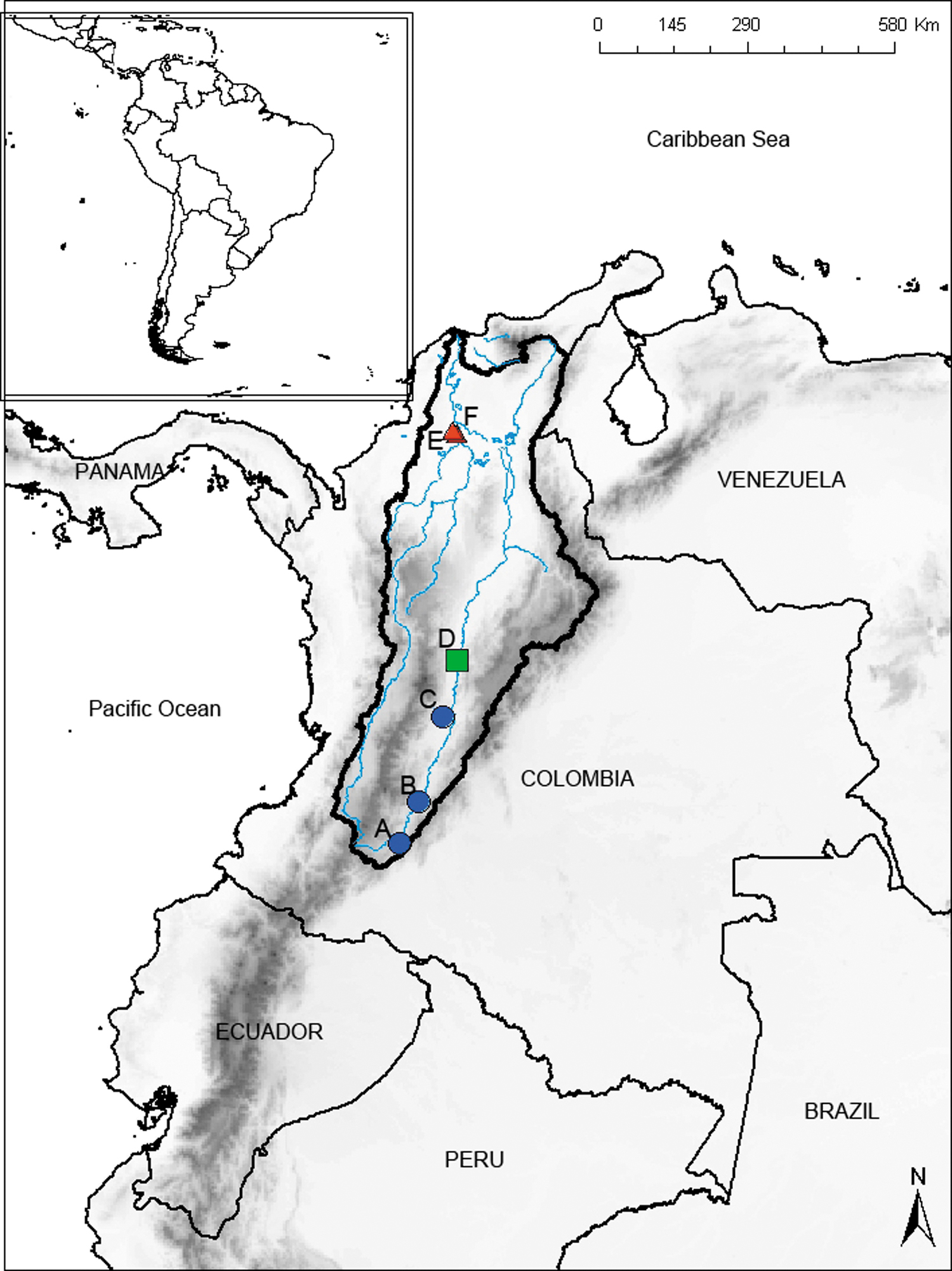
Collection sites of Hoplias malabaricus from the Magdalena River (surrounded by a dark line). Blue circles show upper Magdalena localities: La Boa Creek, Yaguayá, Huila
Collection Sites of Hoplias malabaricus from the Magdalena River (Colombia—South America)
Protocols were carried out according to the Ethics Committee on Animal Use Guidelines (CEUA 29/2013) of the Federal University of Viçosa. Mitotic chromosomes were obtained from anterior kidney cell suspension using the conventional air-drying method. 36 Chromosomes were stained with Giemsa 0.5%, and images were obtained with an Olympus BX53 microscope using cellSens Dimensions software (Olympus). At least 30 metaphase spreads per individual were analyzed to confirm diploid numbers, karyotypic structure, and FISH patterns. Chromosomes were classified as metacentric (m), submetacentric (sm), and acrocentric (a), according to their arm ratios, modified from Levan et al. 37
FISH included three multigene families isolated from the H. malabaricus genome, and some microsatellite motif repeat probes. The first probe was a 5S rDNA repeat copy containing 120 base pairs (bp) of the 5S rRNA encoding gene and 200 bp of the nontranscribed spacer (NTS). 38 The second probe corresponded to a 1400 bp segment of the 18S rRNA gene obtained via PCR from nuclear DNA. 39
The third probe contained a copy of the repetitive satellite 5SHindIII-DNA sequence with 360 bp composed of a 95 bp segment with similarity to the 5S rRNA gene of the first probe and a 265 bp segment similar to the NTS of the first probe; the 5SHindIII-DNA is exclusive of H. malabaricus. 38 The 5SHindIII-DNA and 5S rDNA were labeled with Biotin-14-dUTP by nick translation (Roche, Basel, Switzerland), while 18S rDNA was labeled with digoxigenin-11-dUTP, also by nick translation (Roche, Basel, Switzerland).
Microsatellite motifs with sequences (A)30, (C)30, (CA)15, (GA)15, (TA)15, (CAC)10, (CAT)10, (GAC)10, (GAG)10, and (CGG)10 were synthesized according to Kubat et al. 40 and commercially labeled with Cy3 at the 5′ terminus during synthesis (Sigma, St. Louis, MO). All FISH experiments were carried out according to Yano et al. 41 The 5SHindIII-DNA and 5S rDNA probes were detected with avidin-FITC (Sigma), while the 18S DNA probe was detected with anti-digoxigenin/rhodamine (Roche).
We build ideograms for all the karyotypes that had the report of the 5SHindIII-DNA probe in the Neotropical region, including the UM/MM and LM.
Results
Specimens from the UM and MM showed 2n = 42, FN = 84, and a karyotype composed of 22m+20sm with no morphologically differentiated sex chromosomes (Fig. 2). These samples showed 5SHindIII-DNA satellite probes restricted to chromosome pair numbers 5 (in subtelomeric position) and 6 (in pericentromeric position) (Figs. 2 and 3). However, these samples differed in their 5S rDNA probe patterns: they hybridized in the subtelomeric region of chromosome pair number 4 and on the pericentromeric region of chromosome pair number 8, whereas in MM samples, these probes showed pericentromeric markings on chromosome pair number 4 and two very conspicuous subtelomeric and pericentromeric markings on chromosome pair number 17 (Fig. 4).
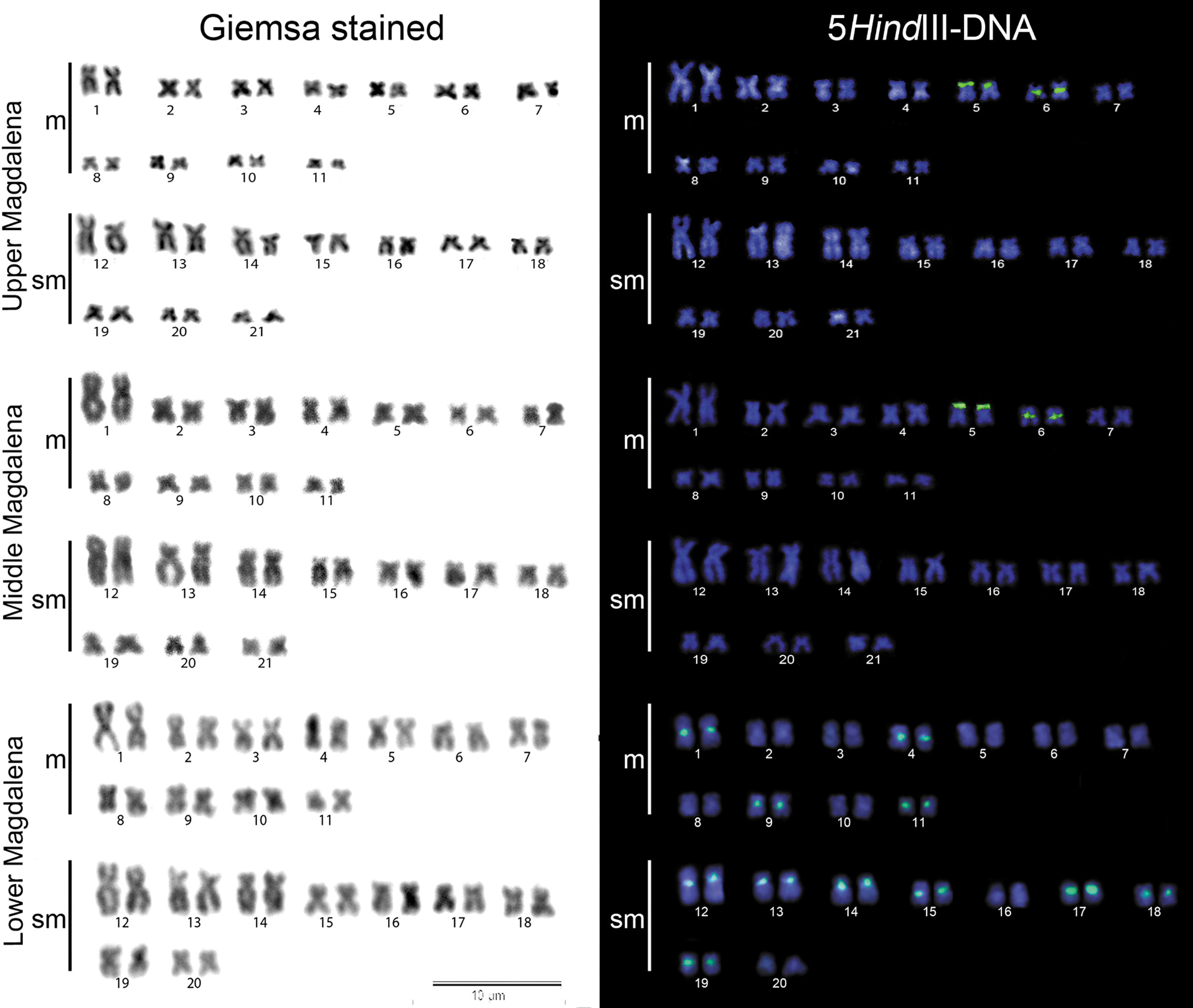
Conventionally stained karyomorphs and 5SHindIII-DNA probe of H. malabaricus from the upper (2n = 42A), middle (2n = 42A), and lower Magdalena River (2n = 40C) in Colombia. Color images are available online.
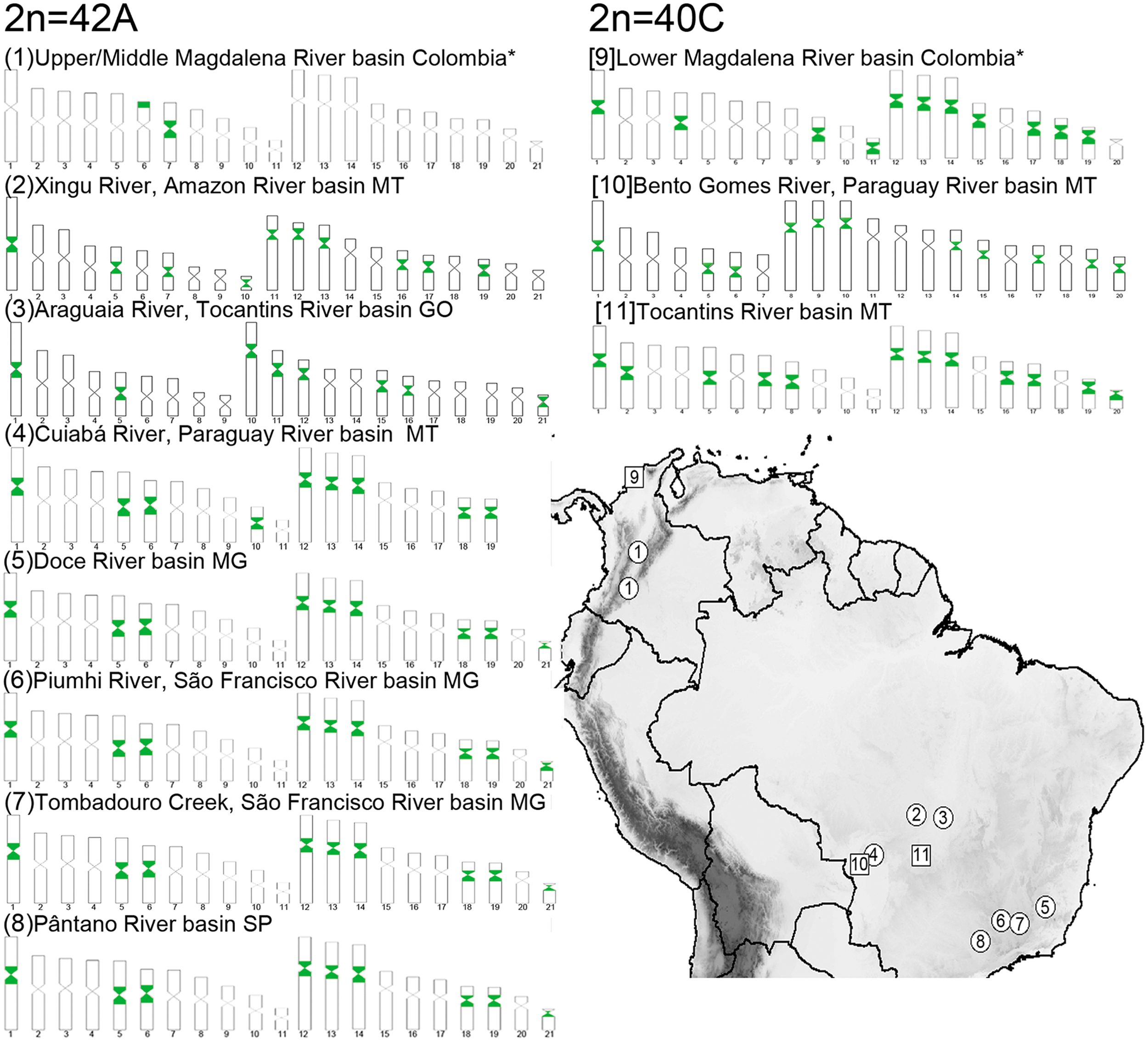
Geographical variation of 5SHindIII-DNA probe patterns in H. malabaricus. Ideograms 1 and 9 correspond to the present study; 2, 3, and 6 from Blanco et al. 18 ; 4, 5, and 8 from Cioffi et al. 200939; 7 in Santos et al. 7 ; 10 from Cioffi et al. 39 ; and 11 in Vitorino et al. 49 Circles in the map represent populations with karyomorph 2n = 42A, squares show karyomorph 2n = 40C from Bertollo. 4 Color images are available online.

Karyomorphs with FISH using 5S rDNA and 18S rDNA probes of H. malabaricus from the upper (2n = 42A), middle (2n = 42A), and lower Magdalena River (2n = 40C) in Colombia. FISH, fluorescence in situ hybridization. Color images are available online.
18S rDNA probes were always subtelomeric and marked some different chromosome pairs in the UM and MM samples: in the UM, these probes occurred on one of the homologues of pair number 21, chromosome pair numbers 12 and 16, and on one of the homologues of pair number 21, whereas in MM samples, 18S rDNA probes hybridized to both homologues of chromosome pairs 15, 19, and 21 (Fig. 4).
All specimens from LM showed 2n = 40, FN = 80, and a karyotype composed of 22m+18sm, and no morphologically differentiated sex chromosomes (Fig. 2). In sharp contrast with the UM and MM samples, in the LM, 11 chromosome pairs hybridized with the5SHindIII-DNA probe in their pericentromeric regions (Fig. 2 and 3). Chromosome pair numbers 6 and 10 showed pericentromeric 5S rDNA sites, whereas 18S rDNA hybridized on the pericentromeric and telomeric regions of chromosome pair number 4. A syntenic pericentromeric site for both rDNAs was evidenced in pair number 6 (Fig. 4).
Of all the microsatellite DNA probes, only the (CA)15 and (GA)15 gave consistent results. In all samples, (CA)15 probe markings were stronger on larger chromosomes, whereas the (GA)15 pattern was more homogeneous among the chromosome pairs. In addition, conspicuous interstitial sites were observed in the largest two submetacentric pairs using both probes (Fig. 5).
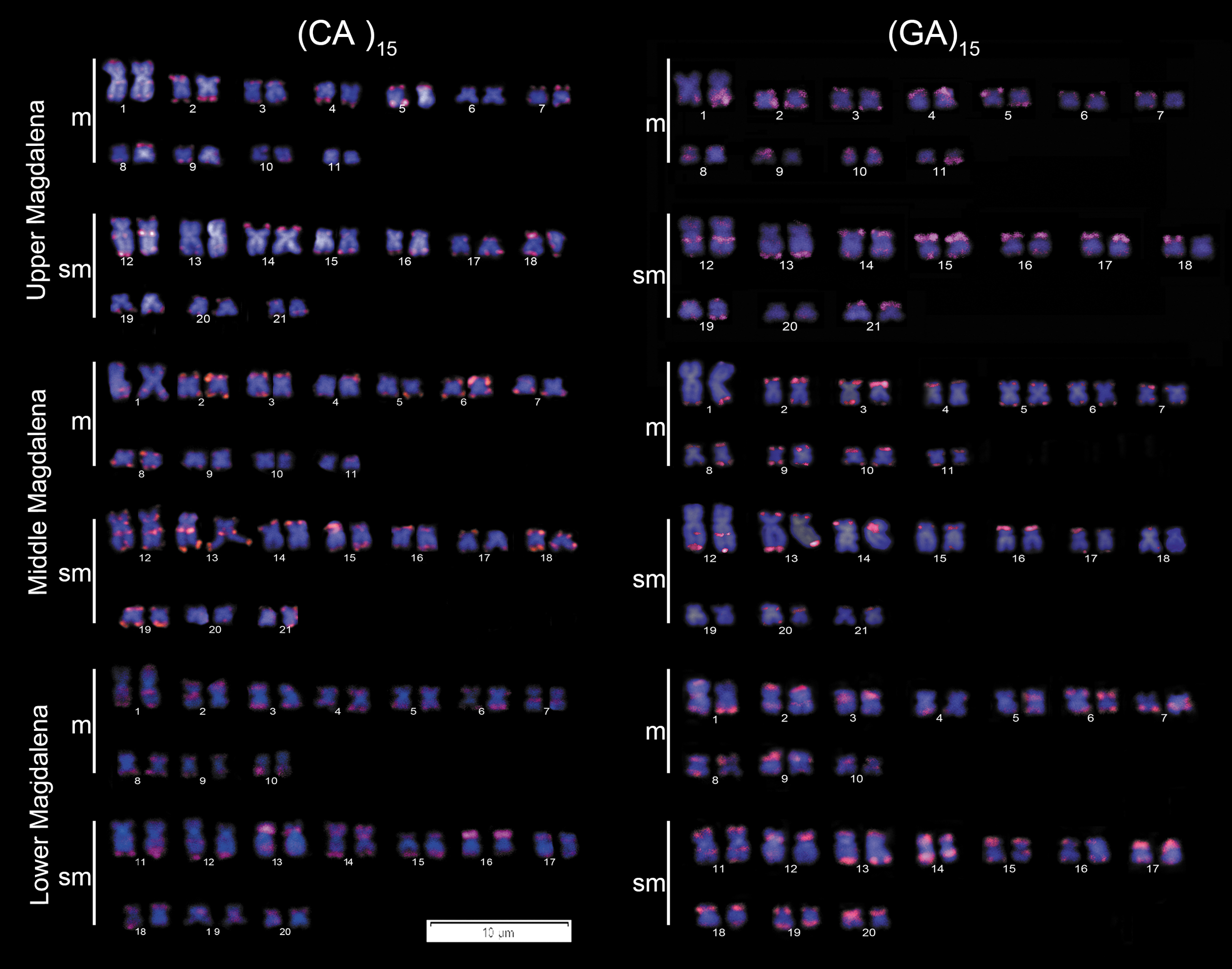
Karyomorphs with FISH using repetitive DNA probes (CA)15 and (GA)15 of H. malabaricus from the upper (2n = 42A), middle (2n = 42A), and lower Magdalena River (2n = 40C) in Colombia. Color images are available online.
In the LM, one male specimen (NI46) showed a heteromorphic chromosome pair, formed by an acrocentric chromosome and a small metacentric and intraindividual variation in chromosome number: 59% of cells were 2n = 40, similar to other individuals from the LM (Fig. 6).

Conventionally stained and FISH of sample NI46 from the lower Magdalena population. Color images are available online.
In this specimen, the 5SHindIII-DNA probe marked 10 chromosome pairs, besides the acrocentric chromosome and its putative homologue (Fig. 6); the 5S rDNA probe marked three chromosomes, showing a pericentromeric marking on chromosome pair number 17 and an interstitial marking on the acrocentric (Fig. 6). The 18S probe hybridized with eight metacentric/submetacentric chromosomes. The (CA)15 repetitive DNA probe hybridized with 15 pairs, including the metacentric member of the heteromorphic pair, whereas the (GA)15 showed 12 bitelomeric markings in different chromosome pairs and both members of the heteromorphic chromosome pair.
Discussion
The paraspecies H. malabaricus is one of the cytogenetically most studied taxa in the Neotropics, and this study is the first one conducted in the trans-Andean region. Based on the karyotype classification of Bertollo et al., 4 the UM/MM karyomorphs were broadly identified as 2n = 42A, whereas the LM karyomorph was considered a 2n = 40C; the individual with an acrocentric chromosome may represent a new karyomorph or some hybrid form.
Together with 2n = 40F, both Magdalena karyomorphs are the most widespread in the Brazilian rivers 6 ; the 2n = 42A karyomorph occurs in the Ctalamochita River (in Argentina) at the southernmost distribution range of this paraspecies. 42 Da Rosa et al. list all the locations where this paraspecies was cytogenetically studied; 2n = 42A and 2n = 40C occur in 37 and 21 out of 79 locations. In the Amazon Basin, sympatry involving these and other karyomorphs was observed in the Trombetas, Madeira, and Aripuanã Rivers. Sympatry has also been observed in the upper and lower Paraná River, and in southeastern Brazil. 6 Sympatric or allopatric distribution of karyomorphs in Neotropical basins reflects the geomorphologically dominated hydrology of each drainage: complex evolutionary histories result in sympatric karyomorphs, such as in the Paraná and Amazon Basins,4,43 whereas isolated basins are usually characterized by one widespread karyomorph. São Francisco and the Doce River Basins harbor two karyomorphs with allopatric distribution: the widespread 2n = 40F occurs along most of its basins, whereas 2n = 42A is restricted to two southern tributaries (the Piumhi 18 and the Pará 7 Rivers). Another exception is the Doce River Basin, where the widespread 2n = 42A karyomorph is syntopic with the 2n = 42B karyomorph in two Holocene lakes. 8
FISH probes also indicated further differentiation among the Magdalena regions. In cis-Andean karyomorphs, the 5SHindIII-DNA probe occurs in a large number of centromeric sites, ranging from 16 to 24 chromosomes in each metaphase, and always involves the largest three submetacentric pairs.7,13,16,18,38,44 This general pattern was also observed in the LM. However, the UM and MM were characterized by telomeric and centromeric 5SHindIII-DNA markings on only two chromosome pairs, placing these samples in a unique condition among all South American populations (Fig. 3). Studied so far, the evolution of copy number of these sites has been considered a reversible, unordered “loss or gain” character condition during the divergence of H. malabaricus. 44 However, if copy number increases through transposable elements (e.g., Crombach Hogeweg, 45 ), the multiplication of these copies may be an irreversible process.
Typically, 5S rDNA sites are located in only one or two chromosomal pairs, showing levels of variation within and among karyomorphs from different basins.18,39,44,46,47 Likewise, in the Magdalena, the chromosome pairs and probe sites were highly variable among samples. The usefulness of this probe as a populational marker is realized when UM and the MM are compared: in the former, chromosome number 8 showed pericentromeric markings, whereas, in the latter, chromosome pair number 17 showed an additional, telomeric marking. In LM, a syntenic condition for both rDNA clusters was observed. Such an uncommon trait was observed for the first time in a 2n = 42A population from the Cuiabá River in Central Brazil, 48 where it was located in a submetacentric chromosome. Those results from the Magdalena River is the first case where this syntenic condition occurs in a metacentric chromosome and may represent rare independent events within this paraspecies.
Cis-Andean H. malabaricus have multiple 18S rDNA markings ranging from 8 to 12 sites, mainly in telomeric regions.18,39 In sharp contrast, the UM and MM presented only six subtelomeric 18S rDNA sites. This probe also occurs in bitelomeric position in several 2n = 42A Brazilian populations in the Cuiabá, 48 São Francisco, 18 and Doce Rivers, 48 and in one 2n = 40C in the Araguaia River. 49 Notably, the LM presented a unique pattern with only two chromosomes bearing these sites, where chromosome number 4 showed both bitelomeric and centromeric sites.
Microsatellites represent one of the most dynamic and widespread components of the genomes. 50 However, the (CA)15 pattern observed in the Magdalena is similar to two other H. malabaricus populations from southeastern Brazil, in the coastal Doce (2n = 42B), and the upper Paraná (2n = 40D) Basins, 51 and therefore, they seem to be very stable within the paraspecies karyotypic evolution. These markers show a conservative pattern also in other Erythrinidae fishes,52,53 and in its closely related family Lebiasinidae. 54 Although Cioffi et al. concluded that this and other repetitive sequences could have some role in the diversification of the family Erythrinidae. 51 Highly homogeneous marking among chromosome pairs has been observed in many species of Prochilodontidae and Anostomidae (Dergam, unp.).
A mosaic condition involving diploid number and chromosome morphology, observed in the LM NI46 specimen, is an unusual mechanism in fish. Furthermore, acrocentric chromosomes are very rare in H. malabaricus, along this paraspecies distribution range: one acrocentric occurs in males of the G karyomorph from the Amazon Basin, these males are 2n = 41, contrasting with 2n = 40 females. 4 The presence of an acrocentric in mosaic condition is similar to the one reported by Diniz & Bertollo (2006) in the erythrinid Hoplerythrinus unitaeniatus (Spix and Agassiz, 1829) from the São Francisco River Basin. 55
Chromosomal mosaicism was previously reported in H. malabaricus in a fish culture station in Parana Brazil; the authors attribute this fact to possible hybridization events between karyomorphs. 56 These authors hypothesized that high levels of environmental pollutants might cause this within-individual variation. Maybe this condition of the LM region is a random effect or the result of gene flow between 2n = 20 and 2n = 42 karyomorphs, and hybrids between these karyomorphs have been reported in the upper Paraná Basin. 14
Besides the mostly allopatric condition observed in the Magdalena River, minor karyotypically differences between the UM and the MM suggest the existence of some level of the structure along the river. In South America, H. malabaricus is known for its lentic water preferences, 57 and the main channel may represent an ecological barrier for dispersal. Congruence between karyotypic and molecular lineages has been demonstrated in southeastern Brazil. 9 Rincon-Sandoval et al. studied the phylogeography of H. malabaricus in NSA and Middle America (Tiura Basin) and found two well-structured lineages (Sp1. and Sp. 2). 31 Sp1 includes samples from the Magdalena, Rancheria, Tiura, and Catatumbo River Basins, and Sp2. included samples from the Catatumbo, Atrato, Rancheria, Sinu, San Jorge, and Cauca River Basins. However, the samples from the Magdalena River included in this study (n = 5) corresponded to the UM and did not show differentiation along the basin.
Hoplias malabaricus from the Magdalena River encompasses the main karyomorph dichotomy observed elsewhere in the Neotropical region, particularly of their FISH markings and their molecular makeup. A general area cladogram based on 32 fish taxa (including 333 species) consistently point four trans-Andean basins as the sister clade area for all the remaining South American taxa. 58
In summary, the H. malabaricus from the Magdalena River encompasses the main karyomorph dichotomy observed elsewhere in the Neotropical region, but conspicuously divergent from all other cis-Andean populations in at least some of the FISH markings and the molecular makeup. This dichotomy pattern is probably the result of the vicariant event that resulted from the uplift of the eastern Cordillera of Colombia during the Miocene, which led to the isolation of the current cis- and trans-Andean basins (including the Magdalena River).59,60
Footnotes
Acknowledgments
This research was supported by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES). We are grateful to the Smithsonian Tropical Research Institute (STRI) in Panamá for sharing tissues, and Pedro Caraballo (Universidad de Sucre—Colombia) and Cristhian Conde (Universidade Federal de ViçosaBrazil) for logistic support in the field.
Disclosure Statement
No competing financial interests exist.
Funding Information
M.B.C. is supported by FAPESP (2016/22196-2) and CNPQ (304992/2015-1).