
Abstract
Gymnotus is the most studied genus of the order Gymnotiformes, but the morphological similarities of the different species make it difficult to identify taxa reliably. The present study is a continuation of the ongoing research into the taxonomic diversity of the stocks of Gymnotus sold as live bait in the Pantanal, Brazil. These studies have been based on cytogenetic analyses, DNA barcoding, and the analysis of coloration patterns. The results of the cytogenetic analysis confirmed the presence of three distinct strains, recognized as Gymnotus paraguensis, G. sylvius, and G. pantanal. However, the results revealed that the molecular operational taxonomic units identified as G. paraguensis actually include a relatively diverse set of fish, separated by considerable genetic distances. As the G. paraguensis specimens also presented considerable variation in coloration patterns, further genetic diversity analyses were conducted on these individuals, to test the hypothesis that more than one species is present in this cytotaxonomic unit. The haplotype network revealed a regional pattern in the distribution of this species. The results indicate that the observed variation in coloration patterns is associated with a high degree of phenotypic plasticity in G. paraguensis. These findings emphasize the importance of using an integrative approach for a more accurate diagnosis of Gymnotus, in particular, the species marketed as live bait for the fisheries of the upper Paraguay River basin in the Brazilian Pantanal.
Introduction
The Pantanal biome is a vast wetland plain in western Brazil, which includes part of the southern Mato Grosso State and the northwestern extreme of Mato Grosso do Sul State, and extending westward into northern Paraguay and eastern Bolivia. The Brazilian Pantanal is divided into 11 distinct wetland sectors, or “pantanais,” based on their relief, soils, and vegetation, and the local flood regime, which is reflected in a highly diversified aquatic biota. 1 The region's fish fauna is thought to include more than 270 valid species, with some still to be described. 2
One important component of this biodiversity is the Neotropical knife fishes of the Gymnotiformes, which has ∼250 species distributed between the Salado River in the Argentinian Pampas and the San Nicolas River in southeastern Chiapas, Mexico.3–5 The Gymnotidae is the most diverse gymnotiform family, and contains two subfamilies, the Gymnotinae (46 species) and the Electrophorinae (4 species). 6 A recent study proposed a new classification for the genus Gymnotus, which now includes six subgenera: Gymnotus, Tijax, Tigre, Tigrinus, Lamontianus, and Pantherus. 7 This subgeneric classification, however, is not totally accepted. 8
Gymnotus has a number of unique characteristics that distinguish it from the other gymnotiform genera, such as cycloid scales, mouth in a superior position, prognathous mandible, eyes in a lateral arrangement, and, in particular, the presence of diagonal light/dark bands along the whole length of the body. 9 These fish are also facultative air breathers, able to inhabit hypoxic aquatic environments,10–12 which determines their popularity as live bait, given their capacity to survive under these conditions for long periods of time.
Fishing of all types is intense throughout the Pantanal biome, and in particular on the upper Paraguay River and its tributaries, where the live bait market is a recurring practice. Many species of fish are commercialized as live baits in the Pantanal region. Among these, it is worth mentioning, due to their high capture frequency, the species Hoplerythrynus unitaeniatus (Erythrinidae); Callichthys callichtys (Callichthyidae); Synbranchus marmoratus (Synbranchidae); Lepidosiren paradoxa (Lepidosirenidae); and Gymnotus spp. (Gymnotidae) among others.13,14 Gymnotus species are the most common fish used as live bait in this region, with the largest quantities being captured in the Brazilian state of Mato Grosso Sul, and then sold as bait in the Mato Grosso State, 15 corresponding to about 50% of fish preference as live bait in the region, followed by crabs. 16
The most recent review of Gymnotus recorded six species from the upper Paraguay River basin: G. carapo australis Craig, Crampton and Albert; G. cuia Craig, Malabarba, Crampton and Albert; G. inaequilabiatus Valenciennes; G. pantanal Fernandes, Albert, Daniel-Silva, Lopes, Crampton and Almeida-Toledo; G. paraguensis Albert and Crampton; and G. sylvius Albert, Fernandes-Matioli and Almeida-Toledo. However, the Secretaria de Meio Ambiente, do Planejamento, da Ciência e Tecnologia de Mato Grosso do Sul (SEMAC-MS) Resolution No. 22/2011 recognized the commercialization of only two species, G. inaequilabiatus and G. paraguensis, as live bait.
Gymnotus species are very similar in their morphology and coloration patterns, which hampers their diagnosis and the correct identification of species.4,9,17,18 The diagnosis of these species is based primarily on the coloration pattern and the arrangement of the banding on the body of the adults. However, the identification of species based on these criteria alone is subject to error, which may often lead to the underestimation of local diversity and the ontogenetic variation of the specimens. For the definition of strategies for the conservation and management of fishing resources and regulation of the trade in live baits, it is necessary to correctly identify both local and commercial species. In this sense, recent advances in the use of chromosomal markers and DNA sequences have contributed to the more reliable identification of specie in the Gymnotus,9,19 and in the establishment of robust hypotheses on the phylogeny of this complex group.5,7
Chromosomal studies have, for example, revealed distinct karyotypes in the different Gymnotus species,19,20 with diploid numbers ranging from 2n = 34 chromosomes in G. capanema 21 to 2n = 54 chromosomes in G. carapo, G. mamiraua, G. paraguensis, and G. inaequilabiatus.19,20,22–24 In addition to this variation in the number of chromosomes, a system of multiple sex chromosomes of the X1X1X2X2/X1X2Y type has been described in G. pantanal, which has 2n = 39 chromosomes in the males and 2n = 40 in the females. 25
The investigation of the karyotypes of Gymnotus has often been complemented with the analysis of mitochondrial genes for the identification of species. These genes are appropriate for this task, given their high mutation rates, maternal origin, lack of recombination, their presence in a wide range of animal cells, and the fact that ∼90% of the mitochondrial genome is composed of codifying regions.26,27 This makes mitochondrial sequences an excellent tool for the elucidation of a range of taxonomic uncertainties, as well as the analysis of the genetic structure of populations.
In this context, the present study continues the ongoing investigation of the knife fish species marketed as live bait in the Pantanal region of Brazil, whose initial results were discussed by Sousa et al. 19 The present study adopted an integrated approach that included morphological, cytogenetic, and DNA barcoding data. The barcoding data were analyzed using a General Mixed Yule Coalescent (GMYC) Model and Automatic Barcode Gap Discovery (ABGD), which provides a robust delimitation of taxonomic units for the verification of genetic and morphological variability, as well as the possible population structuring of the natural stocks of the different species. These analyses provide important insights into the diversity of the fish found in a biologically rich, but poorly studied, region of the Neotropics. The findings of the present study also highlight the fact that the current Brazilian legislation on the capture and marketing of live bait is outdated and does not refer specifically to the species identified in this study. Clearly, then, this legislation requires a thorough review and detailed revision.
Materials and Methods
Study area
The fish specimens analyzed in the present study were all collected by the research team with the help of local fishermen who harvest live bait at five different sites in the Pantanal, including three in the Mato Grosso State, Brazil: (i) Pirigara (Poconé Pantanal: 16°59′33.99′′S, 56°26′10.92′′W), (ii) Estirão River (Barão de Melgaço Pantanal:16°16′36.58′′S, 55°59′17.57′′W), and (iii) Baía das Conchas (Cáceres Pantanal: 16°27′59′′S, 58°09′09′′W), and two in Mato Grosso do Sul: (iv) Paraguay River (17°55′23.56S, 57°27′38.23′′W) and (v) Baía do Castelo (18°35′29.02′′S, 57°32′54.72′′W). Voucher specimens were deposited at the Fish Collection of the Universidade Federal de Mato Grosso (CPUFMT) in Cuiabá, Mato Grosso (G. paraguensis: CPUFMT 4168, 4169, 4426- 4428; G. pantanal: CPUFMT 4165, 4429, 4430; G. sylvius: CPUFMT 4166, 4167, 4424, 4425). The species were identified through meticulous comparisons with the original descriptions, in addition to more publications on the taxonomy of the genus.17,18,28–30 This study was carried out in strict accordance with the recommendations provided in the Guide for the Care and Use of Laboratory Animals. The collection of samples was authorized by the competent environmental authorities: SEMAC—MS and Instituto Chico Mendes de Conservação da Biodiversidade (ICMBIO—license 15226-3).
Chromosomal and molecular studies
The chromosomal preparations were obtained from kidney cells, using the cell suspension technique adapted for use in fish.31,32 The DNA of the specimens was extracted using the saline extraction protocol, 33 and the cytochrome c oxidase I (COI) gene was amplified by polymerase chain reaction (PCR) using the primers Fish F1 5′ TCA ACC AAC CAC AAA GAC ATT GGC AC 3′ and Fish R1 5′ TAG ACT TCT GGG TGG CCA AAG AAT CA 3′. 34
The PCR was run in a final volume of 25 μL containing 1.5 μL of dNTP (0.075 mM), 2.5 μL of 10 × buffer (1 × buffer), 0.5 μL of each primer (0.2 mM), 0.75 μL of MgCl2 (1.5 mM), 0.2 μL of Taq DNA polymerase (Ludwing; 0.04 U), 1 μL of the DNA, and ultrapure autoclaved water to complete the final volume. Initial denaturation was conducted at 95°C for 3 min, followed by 35 cycles of denaturation at 95°C for 30 s, annealing at 50°C for 60 s, and extension at 72°C for 40 s, and a final extension at 72°C for 7 min. The products amplified by PCR were visualized in 1% agarose gel and purified by the enzymatic method using Illustra ExoProStar (GE Healthcare Life Sciences) enzymes. The fragments were sequenced in an automated ABI 3500 sequencer (Applied Biosystems®) supplied by Genotyping Biotechnology Research and Innovation and Myleus Biotechnology, both in Brazil.
Molecular data analysis
The sequences obtained through these procedures were aligned in the Muscle Alignment software 35 in Geneious®36and verified manually in the BioEdit Sequence Alignment Editor 7.0.5.2. 37 The nucleotide sequences were deposited in the GenBank database (accession numbers MH426604–MH426700). The GenBank COI sequences of G. pantanal (KR491603; KR491604; KF359492), G. sylvius (KR491602; MN167131; KF359491; GU701782), G. cylindricus (KJ135116; KR491601), G. coropinae (KR491595; KR491596), G. tigre (KR491625), and G. pantherinus (KR491598; KR491599) were used as the reference sequences for the alignment, and the COI sequence of Electrophorus electricus (MN195141) 38 was used as the out-group.
The saturation of the nucleotide sequences by substitution (transitions and transversions) was verified in DAMBE. 39 The genetic distances within and between species were determined by the Kimura 2 parameter (K2P) nucleotide substitution model, 40 run in MEGA v6.0. 41 The tools available in this software, together with the divergence pattern suggested by the intra- and interspecific distances, were used to plot the arrangement of the taxa in a dendrogram generated by the neighbor-joining (NJ) algorithm 42 with 1000 bootstrap replicates. The optimal threshold (OT) was calculated for the data using the LocalMinima function, implemented in SPIDER (SPeciesIDentity and Evolution in R) 43 in the R software, and used as the reference parameter for species delimitation in the present study.
The species were delimited using the GMYC phylogenetic approach, which requires an ultrametric Bayesian input tree, 44 which was generated in BEAST v.2.4.6. 45 The HKY + I + G nucleotide evolution model was selected for the analyses, based on jModelTest 2.1.7. 46 Three independent runs of 60 million generations were conducted in LogCombiner v.2.4.6, 47 with the first 25% of the runs being discarded as burn-in. The effective sample size was determined in Tracer v.1.6. 47 The GMYC analyses were run on their web server using the single-threshold model. The ABGD method was used to analyze genetic distances in the web interface, using the K2P model of nucleotide substitution, at the default setting. 48
Analyses of population-level genetic diversity were run only for G. paraguensis, which had the largest number of specimens, with the amplest geographic spread, being represented by specimens from all the study localities, which presented marked variation in their coloration patterns. The number of haplotypes (H) was determined by DnaSP v5—DNA sequence polymorphism, 49 and the relationships among the haplotypes were evaluated through the compilation of a haplotype network of the COI data by the median-joining method, 50 run in the fluxus-engineering network program.
The indices of haplotype (h) and nucleotide (π) diversity, and the genetic differentiation of the subpopulations based on the fixation index 51 (Fst), were calculated in Arlequin 3.5, 52 with tests of significance being based on 10,000 permutations. The Bonferroni correction 53 was applied to adjust the p-values. An analysis of molecular variance (AMOVA) was used to verify the partition of the variance within and between the subpopulations.
Analysis of the morphological data
Morphometric analyses were conducted on 66 specimens: 43 identified as G. paraguensis, 13 as G. pantanal, and 10 as G. sylvius. Only adult specimens were included in this analysis, that is, individuals with a total length (TL) of at least 150 mm. 17 Twelve morphometric variables were measured with a digital caliper that had a precision of 0.01 mm,17,30 with minor adjustments. The matrix of morphometric data was dimensionalized using the principal component analysis (PCA), 54 run in the Vegan package of the R platform.
Given its importance for the diagnosis of the taxonomy of Gymnotus, the coloration pattern of the specimens was also analyzed. This analysis focused on the composition and distribution of the bands and spots found on the body of the adult specimens.
Results
Cytogenetic data
The analysis of the karyotypes of 86 Gymnotus specimens revealed the presence of three chromosomally distinct strains: G. paraguensis, with a diploid number of 2n = 54 (48m-sm +6st-a) (66 analyzed specimens: Fig. 1a); G. sylvius with 2n = 40 (36m-sm +4st-a) (10 specimens: Fig. 1b). These species have no morphologically differentiated sex chromosomes. G. pantanal with a diploid number of 2n = 39 (15m-sm +24st-a) in the males and 2n = 40 (14m-sm +26st-a) in the females, and we confirm the presence of a X1X1X2X2/X1X2Y sex chromosome system in G. pantanal.19,25

Karyotypes obtained by Giemsa staining:
Molecular data
A total of 257 partial sequences (546 bp) of the COI gene were analyzed here, and no evidence of saturation by either transition (R 2 = 0.969) or transversion (R 2 = 0.9198) was found, and the index of substitution saturation (Iss) was lower than the critical value (Iss.c). The NJ dendrogram confirmed the existence of three clusters, with branch support of over 90%. G. paraguensis (205 specimens) was the most common species, and was collected at all sampling localities, followed by G. sylvius (21 specimens), which was present at all localities except Barão de Melgaço, and G. pantanal (17 specimens), which was absent from Cáceres (Fig. 2).

Dendrogram obtained by the neighbor-joining method for the COI sequences, based on the Kimura 2 parameter distance, for G. paraguensis, G. sylvius, and G. pantanal collected in the Pantanal wetlands, Brazil. G. cylindricus, G. coropinae, G. tigre, and G. pantherinus were used as reference sequences for the analyses. The bars at the side of the tree represent the results of the analyses of species delimitation. Electrophorus electricus was included as the out-group. COI, cytochrome c oxidase I.
Overall, intraspecific distances ranged from 0.29% to 10.62%, although the distances within G. paraguensis, G. sylvius, and G. pantanal varied from 0.29% to 0.48%. Interspecific distances ranged from 5.51% to 27.50% (Table 1), with an OT of 3.28%. The GMYC and ABGD analyses recovered the same three groups identified by the chromosomal analysis and the NJ dendrogram.
Intra- and Interspecific Kimura 2 Parameter Genetic Distances Among Gymnotus pantanal, G. paraguensis, G. sylvius, G. pantherinus, G. coropinae, G. cylindricus, and the Out-Group, Electrophorus Electricus
The values are percentages (%) with their respective standard deviations (±).
Intraspecific distance.
G. sylvius from Ribeira do Iguape, Atlantic Basin.
G. paraguensis was recorded at all sample sites and at higher frequencies than the other species, but despite having similar karyotypes and belonging to the same Molecular Operational Taxonomic Units (MOTU), the specimens varied considerably in their coloration patterns. This reinforced the need for a more detailed, population-level study of this species. For this, a haplotype network was compiled using 204 G. paraguensis sequences, which presented 68 variable sites, and a total of 77 haplotypes (Fig. 3). The haplotype diversity (h) of this species was 0.893(±0.015), and the nucleotide diversity (π) was 0.00473 (±0.00024). After the Bonferroni correction, the Fst values were significant (p ≤ 0.0033). The Fst index was not significant for most G. paraguensis populations, except for the sample from Cáceres, which presented a moderate degree of genetic differentiation in comparison with the other localities (Table 2). The differentiation observed in G. paraguensis was also confirmed by the results of the AMOVA, which showed that 89.57% of the genetic variation of this species is found within the populations and only 10.43% among the populations, due to the sharing of haplotypes. The fixation index obtained by the AMOVA was 0.104, which indicates moderate structuring 51 (0.05–0.15), influenced primarily by the population from Cáceres, which was significantly different (p < 0.0001) in comparison with the other populations.
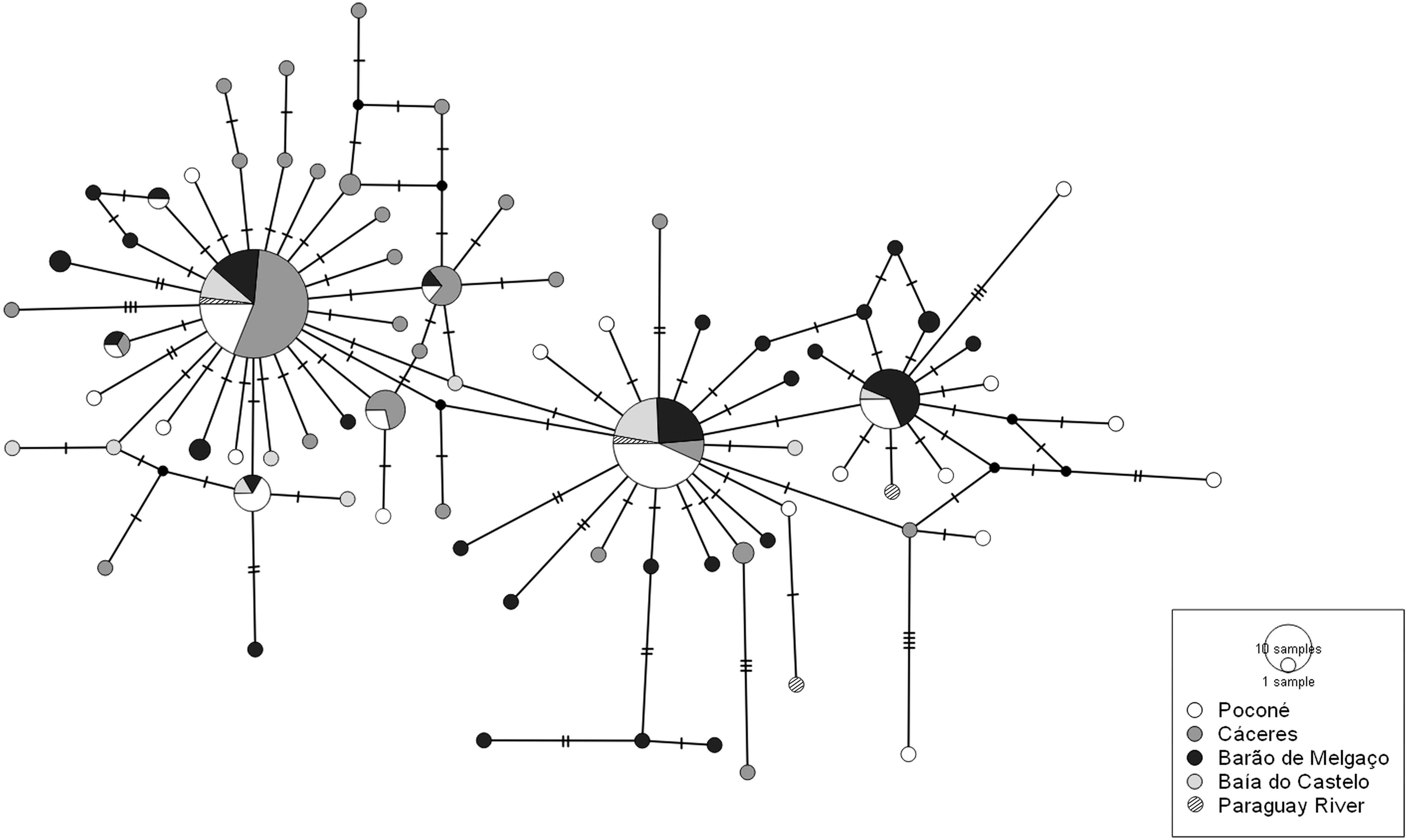
Haplotype network based on the sequences of COI gene of G. paraguensis from Mato Grosso (Poconé, Cáceres, and Barão de Melgaço) and Mato Grosso do Sul (Baía do Castelo, and the Paraguay River). Each circle represents a different haplotype, and its size is proportional to the number of specimens with that haplotype, while the different colors represent the localities.
Pairwise Differences (Fst) Among Gymnotus Paraguensis Populations from Five Localities in the Pantanal Wetlands of Mato Grosso and Mato Grosso do Sul States, Brazil
Significant p-value (p < 0.003) after Bonferroni correction.
MT, Mato Grosso; MS, Mato Grosso do Sul.
Morphological data and brief diagnosis for species
The species herein analyzed from the upper Paraguay River basin present the following characteristics:
Gymnotus pantanal
Twenty-one to twenty-five bands, obliquely oriented, with wavy irregular margins on ventrolateral surface, extending from tip of tail to pectoral-fin base; head not banded, spotted, or blotched; variable color pattern in subadults and adults were identified 55 ; straight body shape in the profile dorsal; overall body subcylindrical; anal fin with 180–233 rays (mode = 222, n = 13); karyotype 2n = 39 (15m-sm +24st-a) for male and 2n = 40 (14m-sm +26st-a) for female chromosomes. The range of analyzed specimens was 160–250 mm TL, mean 190.6 ± 27.6 mm TL. Among the “isqueiros” (people who collect fish to sell), the species is known as “tuvira piramboia,” due to swimming fast and spry (Fig. 4b).

G. sylvius
Gymnotus paraguensis
Twenty-three to twenty-six nearly arranged bands, unpaired and pigmented; dark bands with near straight; conspicuous contrast margins, sometimes discontinuous dark pigment bands in the middle to posterior portions of body with irregular margins, but always defined; portions of head irregularly banded or with scattered chromatophores; considerable variation in its coloration pattern and the distribution of the stripes; anal fin with 175–240 rays (mode = 208, n = 40); karyotype 2n = 54 (48m-sm +6st-a) chromosomes. The range of analyzed specimens was 150–221 mm TL, mean 200 ± 28.33 mm TL (Fig. 5).

G. paraguensis (in median 150 mm TL) from different localities within the study area in the Pantanal wetlands of western Brazil. The heads are shown in close-up to highlight the different patterns of markings. Example of specimens with bands
Gymnotus sylvius
Twenty-three to twenty-seven dark brown-colored bands with wavy, irregular margins and pale interbands wider than dark bands; bands broken into blotches and spots above lateral line at midbody (dorsum), commonly not in distinctive bands; head not banded, spotted, or blotched; anal fin with 207–239 rays (mode = 216, n = 10); karyotype 2n = 40 (36m-sm +4st-a) chromosomes. The range of analyzed specimens was 260–350 mm TL, mean 335.2 ± 31.34 mm TL (Fig. 4a).
The first principal component (PC1) explained 35% of the total variance (Fig. 6a), influenced primarily by head length (82.5%), interorbital distance (72.8%), and the height of the head (71.3%). The second principal component (PC2) explained 28.3% of the variance, influenced by the body width (85.4%), mouth width (69.8%), and the body depth (63.1%).
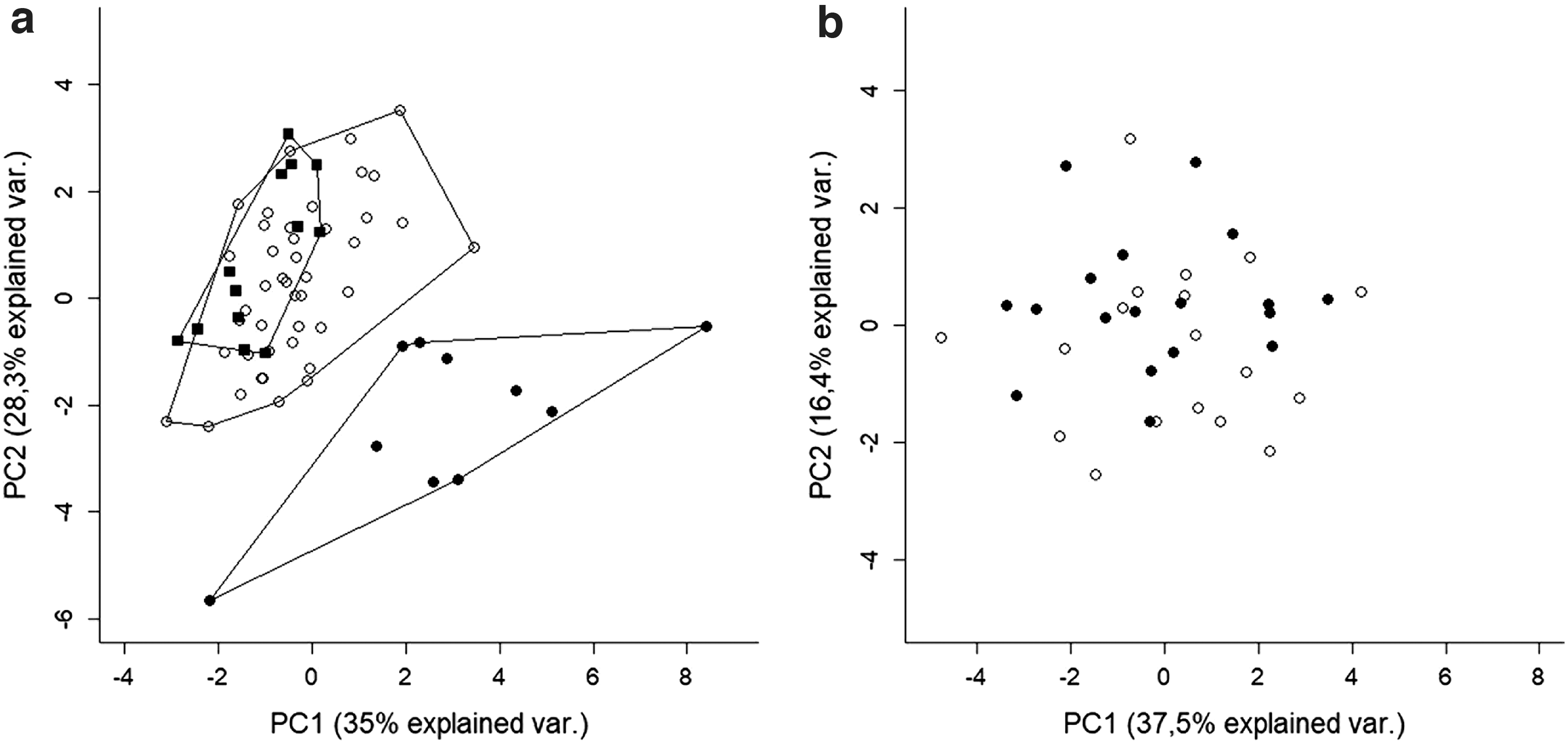
Principal component analysis of 12 morphometric data for:
As G. paraguensis was represented by a much larger sample than the other two species, and its coloration pattern was varied, one possibility is that these specimens did in fact represent more than one taxonomic unit, but diagnostic characters, karyotypes, and results of the molecular analyses fit in the diagnostic limits of G. paraguensis. Given these potential contradictions, a second PCA was run on the morphometric data of this group alone (Fig. 6b). In this second PCA, the first axis (PC1) was responsible for 37.5% of the variance in the data, with the second axis (PC2) recovering 16.4% of the variation. The variables that contributed most to the formation of PC1 were the interorbital distance (86.1%), mouth width (83.0%), and the preorbital length (79.2%), while the variables of PC2 were the eye diameter (75.2%), postorbital length (74,7%), and head length (44.4%).
Discussion
Although the exact number of Gymnotus species used as live bait in the basin of the upper Paraguay River is still unclear, integrated analyses, such as that presented here, provide an important tool for the understanding of the diversity of this fish group and its exploitation by the live bait trade. Our analysis, based on cytogenetics, DNA barcoding, and morphological data, confirmed that three different Gymnotus species were marketed as live bait at the study sites, which is consistent with the findings of Sousa et al. 19 However, the legislation of the Brazilian states of Mato Grosso and Mato Grosso do Sul regulates the live bait trade in only G. inaequilabiatus and G. paraguensis, 56 together with Gymnotus cf. carapo. 57 Given this, the findings of the present study further reinforce the conclusion of Sousa et al. 19 that G. pantanal and G. sylvius should be contemplated by legislation in the upper Paraguay River basin, given their role in this trade, highlighting a deficiency that requires urgent resolution by the competent authorities.
Gymnotus are captured in large numbers by the bait fishers of the Pantanal region, and in many cases, this trade is the principal source of income for their families.16,58 The occurrence of G. sylvius has been confirmed for the upper Paraguay Basin based on karyotype analyses as well as molecular analyses. GenBank COI sequences of G. sylvius from the upper Paraná River basin and the Southeast Atlantic basins were compared with the sequences obtained in the present study (Table 1). This analysis showed that the G. sylvius specimens from the Pantanal (Paraguay basin) and the upper Paraná River basin are genetically close to each other (intraspecific distance = 0.46 ± 0.10). However, this analysis also indicated that the sequences of the specimens from the Atlantic basin belong to a distinct MOTU, which suggests the possible existence of a distinct species that has been identified as G. sylvius. These data reinforce the hypothesis that this species is present naturally in the Pantanal but has not been identified previously from this region by ichthyologists due to the difficulty of identifying the species based on its external morphology.
Three distinct karyotypes were identified in the present study, and the COI sequences were analyzed in detail, using the GMYC and ABGD approaches, which confirmed the existence of three different species in the samples obtained from the live bait trade. These findings confirmed that the Gymnotus species marketed as live bait in the Pantanal region are distinct from those cited in the legislation. The genetic distances recorded among the three taxonomic units identified here are consistent with the criteria proposed for Neotropical fish species, that is, a species-level differentiation of 2%–3%.27,59,60
The low levels of genetic variation found in some congeners are assumed to be a consequence of a relatively recent diversification process, with many species still being found in sympatry or syntopy in the wild. 61 Previous studies of several Neotropical fish species have reached similar conclusions, with a mean genetic divergence among congeners of ∼5%, 60 and exclusive alleles being found in many cases, confirming the low genetic variability found in these species. 62
Similar values were recorded in the basin of the Itapecuru River, 63 in northeastern Brazil, which permitted the reliable identification of 92.19% of the fish species, with interspecific differences of <3% in most cases, demonstrating considerable potential for the discrimination of cryptic species. In the present study, the variation found among congeners was much greater, which allowed the different MOTUS (G. paraguensis, G. pantanal, and G. sylvius) to be distinguished with little uncertainty. To guarantee even more conclusive findings on the observed genetic distances, the “optimal threshold” of interspecific distance was calculated, which indicated that a distance of 3.28% would be the most appropriate threshold for the delimitation of the MOTUS identified in the present study, which is consistent with the findings of a number of previous studies of different fish groups.64–66
In the specific case of G. paraguensis, which was the most common species at all the sampling points and was also the most heterogeneous, given the considerable variation in banding patterns and coloration (Fig. 5), we decided to run a more detailed genetic analysis. The haplotype network compiled from the DNA barcode sequences of this species revealed a clear pattern of regionalization (Fig. 3). There is a predominance of larger haplotypes in the populations from Cáceres, Poconé, and Barão de Melgaço, and the Cáceres population was the most differentiated. However, haplotypes were shared widely among the different study areas, which confirmed that all the specimens collected that had a karyotype with 2n = 54 chromosomes belonged to a single taxonomic unit, referred to here as G. paraguensis.
The low levels of genetic divergence found among the populations of the three Gymnotus species analyzed here may reflect their exploitation as live bait, which likely results in many individuals being transported from one region to another, where they are released, in a type of “man-made gene flow,” which may facilitate the homogenization of the native stocks. Gymnotids are sedentary fish, which migrate only short distances9,61 and are found in sheltered rivers and floodplain habitats, which are typical of the Pantanal wetlands. However, “tuviras” are known to migrate passively under floating rafts of macrophytes, known locally as “camalotes” that move along of the rivers of the upper Paraguay River basin. This may often result in these fish migrating for many kilometers before settling.
The unregulated exploitation of these fish as live bait for carnivorous fish may also alter their population structure and the environments they inhabit. 12 The typical flood pulse of the Pantanal region may also contribute to the homogenization of the Gymnotus populations by facilitating migration, including the formation of the drifting “camalotes.”
Both the genetic and cytogenetic analyses, and the coloration patterns, indicated that G. pantanal and G. sylvius are more easily identifiable. However, considerable variation in the coloration pattern of G. pantanal has already been observed in local populations from the Jacutinga and Pinheirinho streams, located in the upper Paraná River basin. 55 G. paraguensis in the upper Paraguay River basin presents a wide polymorphism in the color pattern as mentioned above, with large numbers of oblique, alternating lighter and darker bands along the entire length of the body.
The variation in the coloration pattern observed in G. paraguensis indicates a high level of phenotypic plasticity in this species (Fig. 5). A similar scenario has been observed in Trichomycterus davisi ( = Cambeva davisi), which presents a high degree of intraspecific variation in its coloration patterns. 63 As this type of variation may be ontogenetic, the identification of species based exclusively on morphological criteria should be treated with caution and a good number of adult specimens. 4
Other studies 29 have referred to possible variation in the shape of the body, especially of the head, both within and among populations of G. carapo, which may be related to the adoption of different substrate-probing strategies. Our results reinforce these conclusions, given the similar level of variation found among the specimens assigned to the MOTU designated as G. paraguensis, which may also reflect the phenotypic plasticity of this species.
In addition to all these considerations, any effective legislation for the regulation of the exploitation of bait fish will have to overcome the fundamental problem of the adequate identification of the gymnotid species involved in the trade. This is especially important, considering that the studies which provide data on the size at first maturation in Gymnotus generally refer to G. carapo,67,68 a species originally described from the coastal drainages of Suriname and French Guiana, including the Brokopondo, Nickerie, and Suriname Districts of Suriname in the Lucie, Suriname, and Corantijn Rivers, and all the departments of French Guiana. 29 It thus seems likely that many studies have analyzed different species, identified erroneously as G. carapo, generating parameters distinct from those expected to represent a single species. Inconsistencies have already been reported in the regulation of Gymnotus species harvested for live bait, such as the fact that the legislation refers only to G. carapo in the Tietê River (Paraná basin), whereas Faria-Pereira et al. 69 had identified only Gymnotus cf. sylvius and Gymnotus cf. cuia.
The cytogenetics of this group nevertheless indicates that it is quite complex. Given the poor taxonomic resolution of the genus, the published karyotypes usually refer to G. carapo, with an ample variation in diploid numbers. For example, Milhomem et al.20,70 cited diploid numbers of 2n = 40 or 42 for Gymnotus cf. carapo (identified as G. carapo orientalis 29 ) from the Brazilian state of Pará, while Milhomem et al. 21 reported 2n = 48 for a population identified as Gymnotus cf. carapo ( = G. carapo madeirensis sensu Craig et al. 29 ) from the Humaitá River. A diploid number of 2n = 52 has been reported for G. inaequilabiatus ( = G. carapo australis sensu Craig et al. 29 ) from the upper Paraná River, 21 in São Paulo State, Brazil.
A karyotype with 2n = 54 chromosomes has also been described by Scacchetti et al. 24 for another population of Gymnotus cf. carapo, a taxon recognized as G. carapo australis, from the upper basin of the Paraguay River. 29 While Craig et al. 29 concluded that these data indicate that G. carapo has a highly polytypic karyotype, 71 Milhomem et al. 70 concluded that the diploid numbers of 2n = 40 and 2n = 42 in sympatric populations of G. carapo (identified as G. carapo orientalis) from Pará represent cryptic species. Regardless of the species concept applied to the interpretation of these data, the karyotypes described up to now further reinforce the taxonomic complexity of the genus Gymnotus. It would nevertheless appear to be most parsimonious to believe that these karyotypically diverse units represent distinct species, and that the variation in coloration patterns observed in G. paraguensis reflects the phenotypic plasticity of this species, a common phenomenon in fish.55,72,73 What is clear is that the subspecies proposed by Craig et al. 29 are not supported by the data, and Fricke et al., 8 for example, do not apply the subspecies category to G. carapo in their catalog of fishes (www.researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp).
Overall, then, when we integrate the different types of data, that is, morphological and genetic, to the understanding of the diagnostic limits of closely related species, we can obtain a more reliable identification of the taxa. These analyses provide essential scientific guidelines for the elaboration of more effective legislation for the regulation of the knife fish bait trade of the upper Paraguay basin, in particular by highlighting how the diversity of these fish has been underestimated and providing insights for the recognition of cryptic species that have been misidentified in previous studies.
Footnotes
Acknowledgments
We are grateful to the Conselho Nacional de Desenvolvimento Científico e Tecnológico and Empresa Brasileira de Pesquisa Agropecuária (EMBRAPA). The collection of fish specimens was authorized by the Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis (IBAMA/ICMBIO permits 12174-2).
Disclosure Statement
No competing financial interests exist.
Funding Information
Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, process 446925/2014-4 and 421733/2017-9 INAU II). Empresa Brasileira de Pesquisa Agropecu´ria (EMBRAPA). Coordenação de Aperfeiçoamento de Pessoal de Nível Superior-Brazil (CAPES)-Financing Code 001.