
Abstract
Over the past decade, the zebrafish has been increasingly employed in biomedical neuroscience research due to its numerous evolutionarily conserved features with mammals. Its simple brain and the several molecular tools available for this species make the zebrafish an appealing model to study mechanisms of complex brain functions, including learning and memory. Most learning paradigms developed for the zebrafish have employed visual stimuli as the associative cue. Spontaneous color preference is a potential confound in such studies. It has been analyzed in zebrafish using colored objects, but with conflicting results. It has rarely been explored with colored light, despite the increasing use of computer-generated visual stimuli. Here, we employ a light emitting diode (RGB-system) light-based color preference task in the plus-maze. In two independent experiments, zebrafish were tested in a four-choice or dual-choice condition by using four different-colored lights (red, green, blue and yellow). Our results suggest a light preference hierarchy that depends on context, since yellow was preferred over green in the four-choice condition whereas blue was preferred over all other colors in the two-choice condition. These results are useful for future color-light-based learning experiments in zebrafish.
Introduction
Survival in nature depends greatly on the ability of the animal to respond quickly to a number of constantly changing environmental stimuli. Color is a particularly salient stimulus for diurnal species, which may use this information during navigation, 1 foraging, 2 or mating. 3 Color cues may be important for a variety of animals in the wild, and thus they have been successfully employed in learning paradigms conducted in the laboratory too. However, not all colors may be created equal. Species-specific spontaneous color preferences may exist that evolved according to the ecological demands of the natural environment of the given species. For instance, honeybees have been found to prefer violet or blue color, 4 which happens to be the color most often associated with flowers offering a high nectar yield.
In an experimental setting, such spontaneous color preference could create a bias, especially in associative learning tasks. Spontaneous color preference/bias has been investigated in a number of species, including invertebrates4–6 and vertebrates.7–9 The zebrafish is one such vertebrate. It has been becoming one of the most preferred laboratory organisms of biomedical research due to its practical and biological simplicity. For example, this small (4 cm long) fish is highly prolific and can be housed in large numbers in small fish tanks in a cost-effective manner. Its brain weighs only 50 mg and has about 1 million neurons, making it the simplest laboratory vertebrate used in biomedical research. However, it has a sophisticated behavioral repertoire and evolutionarily conserved features at practically every level of biological organization studied, from the nucleotide sequence of its genes to the psychopharmacological effects of drugs. 10 The zebrafish is particularly appropriate for behavioral studies, because visual cues are experimentally/technically easy to manipulate, and vision is an important modality for the zebrafish in a variety of behavioral contexts.11,12 Indeed, visual stimuli have been successfully employed in appetitive as well as aversive conditioning with zebrafish.13–15 For this reason, and because the zebrafish is a tetrachromatic species, their color preferences have started to be widely explored (Table 1).
Summary of Findings Obtained and Basic Methods Employed in Color Preference Studies Conducted with Zebrafish
WT corresponds to the fish obtained in pet store, or genetically variable fish populations from other vendors.
LED, light emitting diode; WT, wild-type.
However, these studies have yielded conflicting results (Table 1). The reasons for this may be numerous and may include, for example, the use of genetically different populations/strains of zebrafish, different experimental protocols, and different colors employed.16–24 For example, testing adult wild-type (WT) zebrafish with different colored materials, Spence and Smith found zebrafish to prefer red, whereas another study demonstrated preference for blue and green. 16 A common inconsistency with color-based paradigms in zebrafish is the material/object that is used as a colored cue. Color preference using materials such as plastic, 16 gravel, 18 or paper 22 have all been conducted. An important problem with the colored material-based visual stimulus delivery method is that macroscopic as well as the microscopic surface characteristics of the objects employed may influence how light is reflected from the object. In other words, idiosyncratic object-specific features may complicate the interpretation of color preference/discrimination study results. Further, the onset and offset of a color-based stimulus is difficult to control using objects, as it may require moving or placing the colored physical object or barriers in or around the object, for example, lowering a color card, or removing a plastic sheet to reveal the color cue. To alleviate these methodological issues, investigators have started utilizing LCD screens or white or RGB system-based light emitting diodes (LEDs) as the source of the visual stimuli.12,23–29
The results obtained with colored objects and light often differed. As mentioned earlier, honeybees were found to prefer violet or blue color, 4 according to a study in which the investigators employed colored cardboard as a means of presenting the color. However, when using colored light, bees have been found to prefer green.30,31 Colored-light preference has been explored in zebrafish too.23,24 In these studies, LED panels as well as color filters were employed, and preference for spatial areas illuminated distinctively was investigated. The results found were often contradictory to those obtained using colored objects (Table 1). These results also highlighted the importance of other methodological aspects of color presentation, including the position of the light source, or the genotype (strain origin) of zebrafish on the colored-light preference. 24 Although these latter two studies using lights of different color improved our understanding of color preference in zebrafish, they suffered from two main drawbacks: One, the lights the fish were supposed to discriminate were used to illuminate entire areas of the tank. However, color discrimination in nature is more likely based not on colored backgrounds but on color reflected back from particular objects. Two, the light employed emanated from the bottom, which from a technical standpoint was a good method, but again, from an ethological/ecological standpoint was inappropriate due to the artificial nature of this setup, an issue that is clearly demonstrated by the light-position-dependent effects found in one of these two studies. 24 Notably, many learning tasks with zebrafish present visual cues on the side of the tank, including with the use of LCD screens.11,12 However, preference for lights of different color has not been explored using such setups.
In this study, we investigate, for the first time, the preference of zebrafish for colored light by using an RGB-LED system in a cross-shaped tank in which light panels were placed at the end of each arm. The goal of our study is to explore color preference in zebrafish in a context that is both ethologically relevant and appropriate for learning and memory studies. In the first experiment, we describe how experimental zebrafish choose among four differently colored LED lights presented simultaneously, a multiple-choice task performed in a plus maze. This first experimental condition aimed at testing the fish preference in a condition closer to a natural one (i.e., choice among multiple different visual stimuli appearing in the mid-water level). In the second experiment, another naive group of zebrafish was tested in a binary choice task, in which pairs of color lights are presented in all possible combinations and the preference by zebrafish is quantified. We chose to test the fish in this second setup, as the two-color discrimination method has been often employed in learning and memory tasks in a variety of species.13,32–35
Materials
Animals
Adult WT zebrafish (6 months old male) raised in the University of Toronto Mississauga UTM) zebrafish facility were used in these experiments. The parental generation of these fish was obtained from a local pet store (Big Al's Aquarium Warehouse, Mississauga, Canada). Strain-dependent idiosyncratic responses in a colored-light preference task have already been demonstrated. 24 We chose the WT population as the subject of our analysis for the following reasons. The WT zebrafish are genetically variable and have a high degree of heterozygosity in their genome, as the parents of this population originate from a commercial breeding facility (Singapore) where the effective population size is large. As discussed earlier in numerous publications using zebrafish (e.g., review by Gerlai 36 ), as well as mice and rats (e.g., review by Kafkafi et al. 37 ), a genetically heterogeneous population, or outbred strain, is more representative of the species and, unlike inbred strains, it is less likely to possess genotype-specific idiosyncratic features. We chose the WT population of zebrafish as the subject of our study to avoid such potential idiosyncratic features. We recognize that characterization of standard zebrafish strains is a must, but we argue that the first step in this characterization is the establishment of a baseline that can be best obtained by the analysis of zebrafish that have not experienced random genetic drift-related phenotypical changes. All experimental subjects were individually housed in 2.8 L plastic tanks (ZT280; Aquaneering, San Diego, CA) that were part of a multi-phase filtration stand-alone high-density rack system (ZD660; Aquaneering) for the duration of the experiment. This system allowed us to maintain stable water parameters (Temperature = 27°C, pH = 6.8, Salinity = 250 μS, NH₄OH = 0 ppm, NO2− = 0 ppm, NO3− = 0 ppm). Pellets were provided to the fish twice a day (Zeigler, Tropical-fish micro pellets). Zebrafish were kept on a 14 h light: 10 h dark cycle (lights started to turn on at 6:30 am and slowly increased in brightness to maximum by 7 am. Lights started to turn off at 8 pm, with lights completely off by 9 pm). For each experiment, a sample size around 20 fish has been chosen to match the number of fish usually employed in previously published color discrimination and learning tasks with zebrafish, thereby facilitating a comparison with these studies.
Apparatus and color stimuli
The test tank was a plus-shaped maze (Fig. 1A). The physical dimensions of the four arms of the maze were identical (length: 50 cm, width: 11 cm, height: 20 cm). Plastic sliding doors were placed at the start of each arm. A center area was created when all doors were closed, and this served as the departure box for the test fish. At the end of each arm, an RGB LED panel (Medium 16 × 32 LED; Adafruit, New-York, NY) controlled by a Raspberry Pi card (Raspberry Pi 3B+; Raspberry Pi Foundation) could display different colors. The colors used in the experiment were: red (wavelength peak: 615 nm), green (peak: 509 nm), blue (peak: 454 nm), and yellow (peak 1: 614 nm, peak 2:454 nm). The brightness of the LED was controlled across all colors and was measured at the center of each arm as 145 lux.
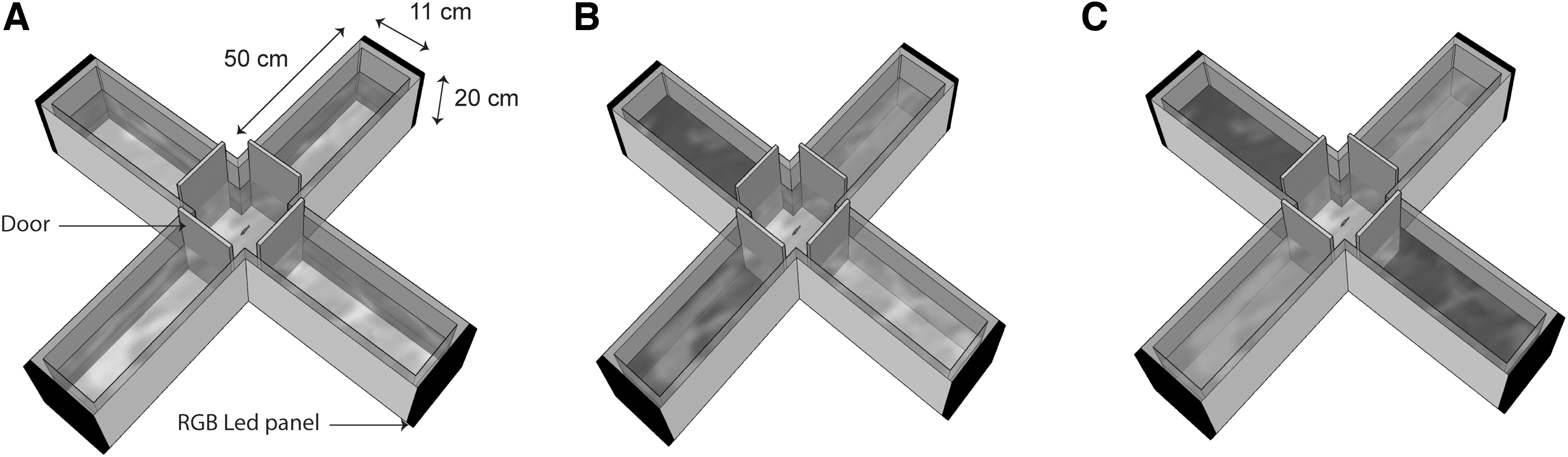
The plus maze employed to test color preference.
Procedure
Habituation
During the experiment, fish were transferred with a net from their home-tank to the experimental plus maze. To reduce stress potentially induced by handling, a habituation protocol was implemented as follows. On the first day, 24 fish were transferred into the plus maze for 1 h. On the second day, 2 groups of 12 fish were transferred into the plus maze for 1 h each. On the third day, four groups of six fish were transferred for 30 min each into the experimental tank. On the fourth day, groups were once again divided into two equal parts; thus, eight groups of two fish were placed into the tank for 30 min each. On the last day, each experimental fish was individually transferred into the tank and stayed there for the duration of the test, which in Experiment 1 was 6 min, and in Experiment 2, 12 min. Also, during the 3 last days of habituation, fish began the habituation for the operation of the departure box, that is, the doors of this box were simultaneously opened to habituate the test fish to the procedure.
Experiment 1: multiple color choice
In the first experiment, fish were tested for their preference of light in a four-choice color task. The rationale for this was that in nature, unlike in most binary color discrimination studies, zebrafish do not encounter only two colors to choose from. Fish (n = 22) were placed singly into the departure box for 2 min (Fig. 1B). At the same time, the LED panels started to display the four different-colored lights from the side at the end of each arm of the maze. Each color light was pseudo-randomly assigned to an arm for each fish. Subsequently, all four doors were simultaneously opened, and the experimenter left the experimental room. Fish explored the plus maze for 4 min, after which the colored lights were turned off and the fish was removed from the tank.
Experiment 2: binary color choice
In the second experiment, fish were tested in a context of a usual learning and memory experiment. That is, the zebrafish were presented with two colors to choose between. Fish were placed singly into the departure box for 2 min (Fig. 1C). At the same time, the LED panels started to display the two colored lights, again positioned at the end of the arms of the maze. Opposite arms always displayed the same color (Fig. 1C). First, the doors were simultaneously opened, and the experimenter left the experimental room. Fish explored the plus maze for 4 min, after which the lights were turned off for 2 min. Subsequently, the position of the colored light was switched and the fish were allowed to explore the maze for 4 additional minutes (e.g., if green was displayed on the right and left arms during the four first minutes, then subsequently green was displayed on the top and bottom arms for the last 4 min). After 12 min of recording, the lights were turned off and the fish was removed from the tank. Every color combination was tested (Red vs. Green, Red vs. Blue, Red vs. Yellow, Green vs. Blue, Green vs. Yellow, Blue vs. Yellow).
Quantification of behavior and statistical analysis
The experiments were recorded by using an overhead camera (HDR-CX430V; Sony) mounted on the ceiling above to the plus maze. The camera was also present during habituation. The videos were analyzed with the software Boris. 38 A fish was considered as choosing one arm once its entire body crossed the door limit in the arm. For each fish, the cumulative time spent in each arm and each color was quantified.
For each experiment, the time spent in each arm (side or color) was first square root-transformed to meet the criteria of normality and homogeneity of variance. Subsequently, the data were analyzed by using repeated-measures analysis of variance (ANOVA) with fish ID as a random factor (individual effect). In addition, the color combination used in Experiment 2 was employed as a co-variable.
The Tukey Honestly Significant Difference post hoc test was used for multiple comparison. Statistical analyses were conducted by using the R 3.6.2. 39 The package nlme was used for repeated-measures ANOVA of time.
Results
Experiment 1: multiple color choice
Fish did not spend significantly more time in any one arm than in the others, that is, they showed no spatial bias (Fig. 2A; F = 1.44, df = 3, p = 0.24). A significant color preference was observed in the amount of time that the fish spent in the colored arms (Fig. 2B; Color effect: F = 4.1, df = 3, p = 0.01), that is, fish spent significantly more time in the yellow arm compared with the green one (Fig. 2B; Red vs. Green: z = 1.43, p = 0.91; Red vs. Blue: z = 0.12, p = 1; Red vs. Yellow: z = 2.02, p = 0.26; Green vs. Blue: z = −1.31, p = 1; Green vs. Yellow: z = 3.46, p = 0.003; Blue vs. Yellow, z = 2.15, p = 0.19).

Arm preference of fish during the four-color choice task (experiment 1, n = 22).
Experiment 2: binary color choice
Fish explored the four arms regardless of the combination of color used, that is, the Color Combination effect was found to be non-significant (F = 0.33, df = 5, p = 0.89). Consequently, the data with regard to arm were pooled (Fig. 3). Fish spent significantly more time in the left arm compared with the other arms (Fig. 3; F = 19.85, df = 3, p < 0.0001). This side bias, however, did not lead to a tendency in the color discrimination results, since the position of color was changed halfway through the test. Consequently, a fish that spent all its time in one arm spent an equal time in both colors in its binary choice test.
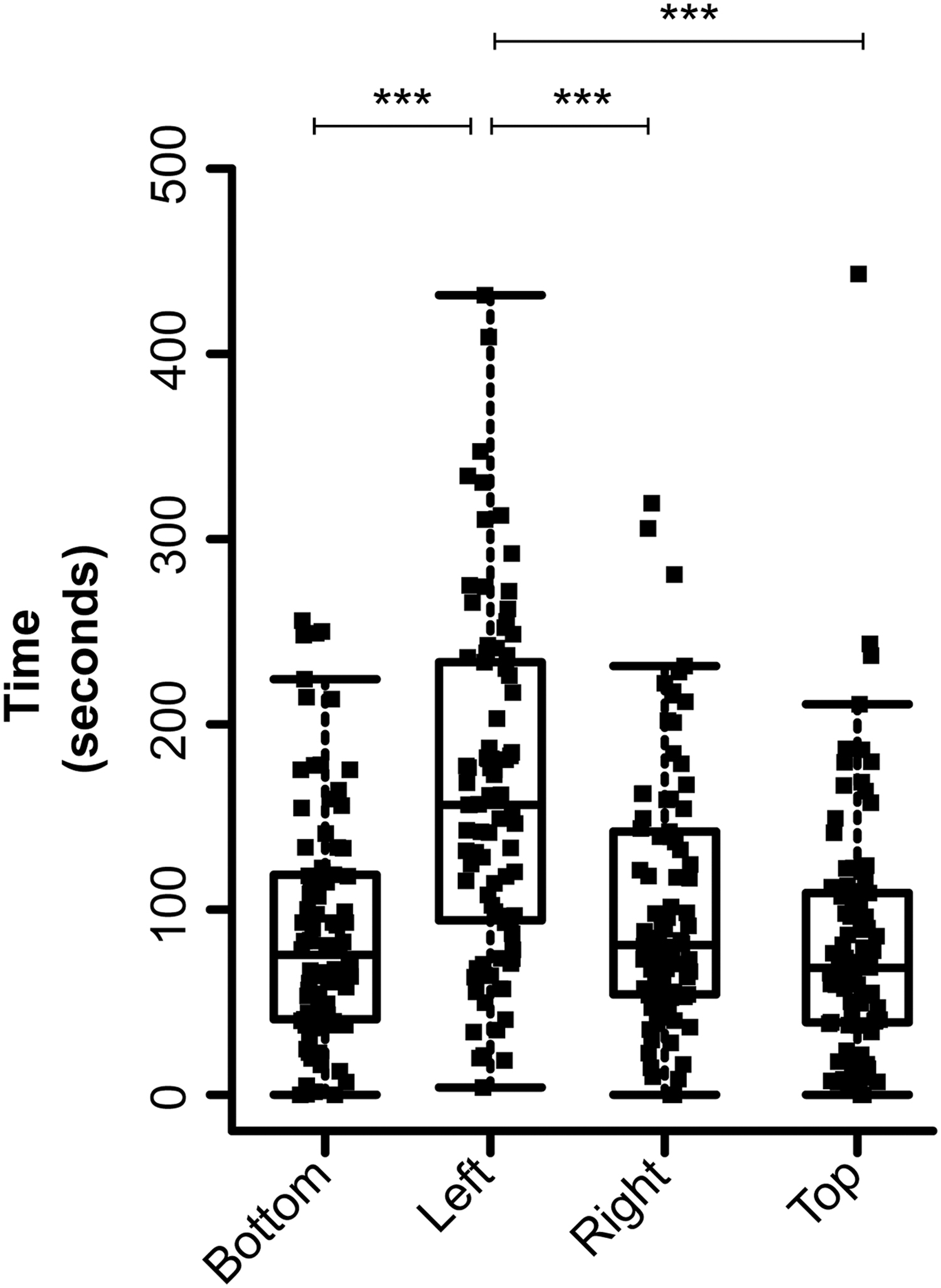
Spatial distribution of fish during experiment 2 (n = 86). Cumulative time (in seconds; median, quartile, and individual points) spent in the bottom, left, right, and top arm whereas the colored lights were displayed. Results suggest significant bias toward the left arm (Bottom vs. Left: z319 = 6.21, p < 0.001; Bottom vs. Right: z319 = 1.32, p = 1; Bottom vs. Top: z319 = −0.83, p = 1; Left vs. Right: z319 = −4.88, p < 0.001; Left vs. Top: z319 = −7.04, p < 0.001; Right vs. Top: z319 = −2.15, p = 0.19). ***p < 0.001.
When red was contrasted with green, experimental fish (n = 13) did not spend significantly more time exploring one or the other color (Fig. 4A; F = 1.61, df = 12, p = 0.23). However, when red was contrasted with blue, fish (n = 15) spent significantly more time in the blue arms than in the red arms (Fig. 4B; F = 11.2, df = 14, p < 0.01). When fish (n = 14) had to choose between red and yellow, they spent significantly more time in the yellow arms compared with the red (Fig. 4C; F = 7.66, df = 13, p = 0.016). For a choice between green and blue, fish (n = 15) spent significantly more time in the blue arms than in the green ones (Fig. 4D; F = 21.03, df = 14, p < 0.001). When green was contrasted with yellow, fish (n = 15) spent significantly more time in the yellow arms than in the green ones (Fig. 4E; F = 8.02, df = 14, p = 0.01). Finally, in a blue–yellow dual, fish (n = 14) spent significantly more time in the blue arms than in the yellow arms (Fig. 4F; F = 13.66, df = 13, p = 0.003).
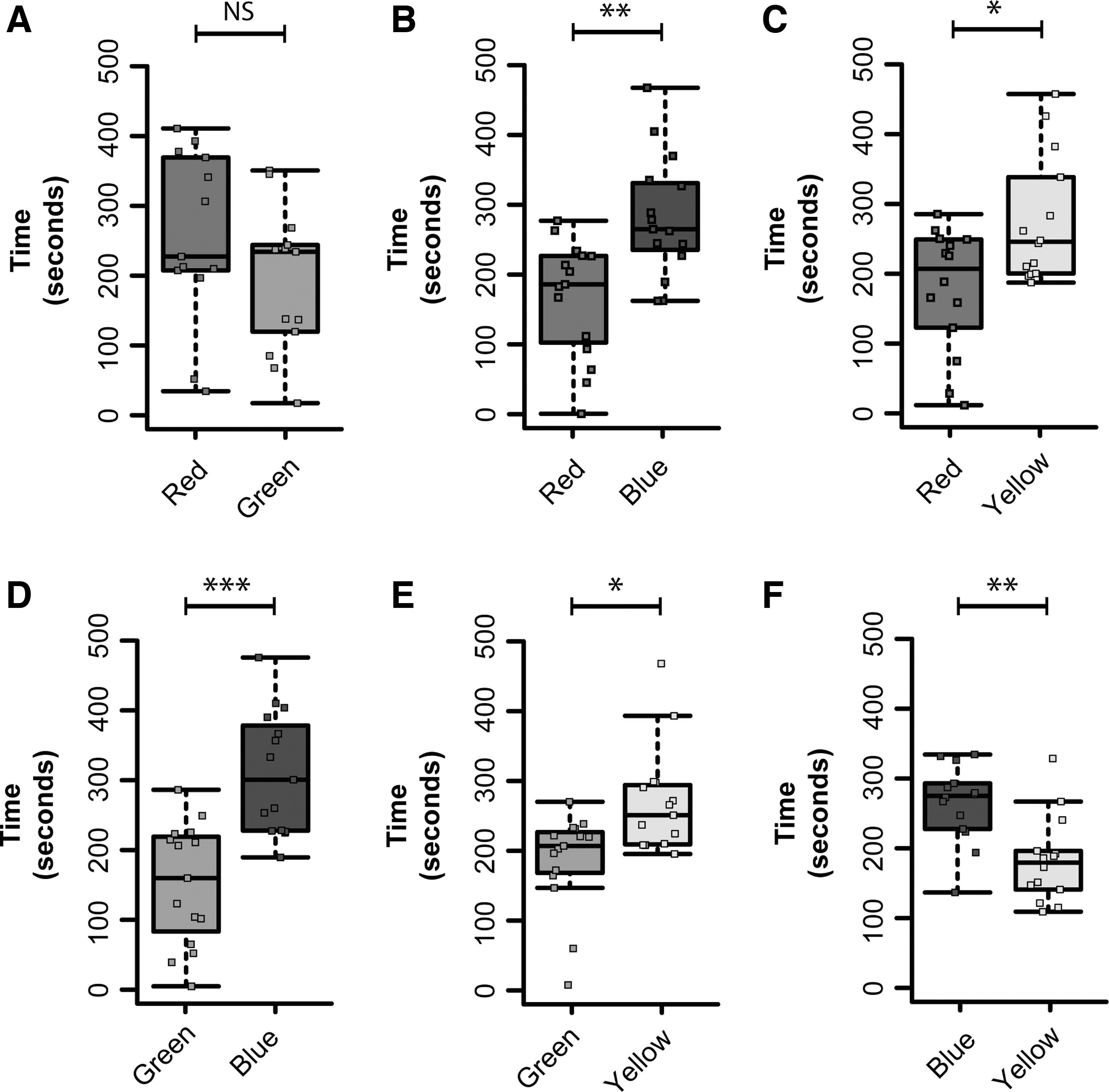
Color preference of fish in the binary color choice task (experiment 2). Cumulative time spent in the arm with red versus green
Discussion
Zebrafish have been shown to utilize color cues in a variety of behavioral contexts.13,14,40 The color preference in zebrafish has been explored by using colored objects as stimuli, including colored paper or gravel, but more recently also employing lights of distinct wavelengths (for references and summary of methods see Table 1). Our study is the first to study how zebrafish choose between and among lights of distinct wavelengths emitted by RGB LEDs positioned on the side of their test apparatus. The colored light stimuli we employed and the manner in which we presented them are particularly important considering the ethology of the zebrafish and the increasing use of computer presented visual stimuli in a variety of behavioral contexts, including in the analysis of learning and memory. We found a significant light preference hierarchy in zebrafish that depended on whether the fish choose between two or among four colors. For example, zebrafish showed significant preference toward yellow over green in the four-choice condition, whereas blue was preferred over all other colors in the two-choice condition.
The results cited earlier only partially confirm previously published findings. Notably, prior studies exploring color preference in zebrafish are replete with inconsistencies (Table 1). The discrepant findings may be due to a variety of methodological differences (e.g., experimental setup, strain, colored stimuli employed etc.). Despite these inconsistencies, our current results point toward an emerging picture in the color preference of zebrafish. For instance, except when opposed to the blue light in dual choice, we observed preference for yellow light in both our four-choice and binary choice tasks. Interestingly, this same preference was also observed in a prior study that employed an RGB LED computer screen to test social response to moving images of conspecifics whose color and body proportions were experimentally controlled. In this study, the authors found zebrafish to prefer the unnaturally yellow-colored image. 40 However, such yellow preference was not detected in studies that did not employ RGB light emitting systems (LEDs or computer screens) but instead used colored objects.l8,19 Color preference in these latter studies may be significantly affected by macro- and microstructural surface properties of the object that may alter the reflection, and thus appearance, of light.
Another possible explanation for the inconsistencies is the context under which color discrimination/preference is tested. Unlike in experiment 1 in which we tested a four-way color preference of zebrafish, in experiment 2 we analyzed their binary choice. This latter experiment did confirm yellow preference, against red and green, but not when yellow was contrasted with blue. In the latter contrast, blue was found to be significantly preferred. Blue was also found to be significantly preferred to all other colors. Choice-task specific findings have also been reported by Avdesh et al., who found fish tested in two different experimental contexts not to show the same color preference. 18 When these authors tested fish in a preference place box, they found them to be equally attracted to green, red, and yellow, whereas when they used a T-maze, they found the fish to be more attracted to green and red than yellow. In our own experiments, the change of context, that is, choice among multiple colors in experiment 1 versus between two colors in experiment 2, could similarly explain the differential findings between these two setups, results that highlight the importance of systematic color discrimination/preference analyses. We also note that Avdesh et al. observed aversion to, and not preference for, blue when using colored physical objects. 18 At this point, we have no explanation for this discrepancy, other than the fact that in our study we utilized light emitted by LED and not a physical object.
Finally, although this aspect was not the central focus of our study, we did observe a spatial bias in the choice made by our experimental zebrafish. Such spatial bias may interfere or confound color choice responses when, unlike in our current study, the colors to be chosen are not arranged in a spatially randomized or systematically alternating manner. Briefly, in our second experiment, we found zebrafish to prefer to move to the left irrespective of the color shown on that side. One could argue that finding this spatial bias significant was because for this analysis we pooled all fish of experiment 2, and thus the sample size was six times higher than for the other comparisons. Nevertheless, we note that the left bias we observed is in accordance with findings by others who reported lateralization of responses to visual stimuli in both larvae and adult zebrafish. For example, the left eye has been found to be more involved in tasks associated with novelty exploration and detecting/processing small changes in the environment, including the appearance of new color. 41 On the other hand, right eye dominance has been demonstrated by these very same authors for tasks that involved planned motor responses to bite an object (foraging response) as well as when the fish had to choose between two routes. 42 A similar finding was reported in a learning study with adult zebrafish, in which experimental zebrafish were found to choose a route to the right preferentially. 43 We emphasize, however, that the spatial bias (for the left) we observed in the current study was only in the second experiment despite that both experiments were conducted in the same room, with the same tank and at the same time.
Last, we briefly consider the neurobiology of color perception in zebrafish. Since the brightness of the visual stimuli we employed was equal, the preference observed during our experiments, we argue, must have been due to the wavelength of light, whose perception starts with the photoreceptors in the retina of the fish eye. Although the yellow objects used in other studies were characterized by a ∼550 nm wavelength corresponding to yellow, indeed, the yellow light used in our experiment was defined by two wavelength peaks, because the RGB-LED system produces this light by combining red (wavelength peak: 615 nm) and green (wavelength peak: 509 nm) lights. This itself may account for the differences between the results of studies that employed colored-object versus colored-light-based discrimination or preference tests. The zebrafish possesses five different photoreceptors: one rod (light intensity) and four cones (color) that differ in their wavelength sensitivity. Within the four cone types, the blue and the ultra-violet cones stand separately whereas the red- and green-sensitive cones are fused to form double cones.44,45 Importantly, despite this fusion, both red and green cones process light corresponding to their wavelength sensitivity separately, just like in most mammalian species. Consequently, the presentation of yellow light will excite both the red and the green cones. Notably, in the zebrafish, the red and green cone generated signals are pooled at an early stimulus processing stage. 46 This might explain why yellow is preferred to blue and red when a colored-light stimuli are used, but that this preference is not observed when object-based color stimuli are employed. Last, we note that blue and ultraviolet cones have been found to be involved in phototaxis (preference toward these wavelengths) in larval zebrafish when tested with an RGB LED system, 46 a finding that is in line with the blue preference we observed in the binary choice task (experiment 2) conducted with our adult zebrafish.
In conclusion, our results demonstrate significant colored-light preference in zebrafish when tested with an RGB LED system. The experimental zebrafish we employed in our study were naive to the experimental task and procedure, have never been trained or tested in color choice tasks, or in any other behavioral tasks, and thus their responses we regard as spontaneous, arising as a result of their genetic predisposition, that is, species-specific features. Whether the performance of our experimental fish has been influenced by the artificial laboratory environment, and how zebrafish in nature would respond is unknown at this point. Irrespective of this, however, our findings showing an untrained bias toward and against certain wavelengths of light highlight the need for systematic testing for color preference in new experimental contexts. The results of such studies will provide crucial information on how to employ visual stimuli in a variety of zebrafish studies, including analysis of learning and memory, our own long-term goal.
Footnotes
Authors' Contributions
A.B. designed the experiment. S.N. and C.B. collected the data. A.B. analyzed the data. A.B. and R.G. wrote the article.
Acknowledgment
The authors thank Gideon Humphrey for his help in measuring the wavelength of the colored light.
Ethical Statement
This study has been approved by the Local and University Animal Care Committee and complies with local, provincial, and federal guidelines that govern the ethical and human use of animals in research in Canada. Each of these experiments followed the protocol number 20012491.
Disclosure Statement
The authors declare no competing or financial interests.
Funding Information
A.B. was funded by the FYSSEN Foundation as a post-doctoral fellow. The project has been supported by NSERC grant (311637) and by The University of Toronto Distinguished Professorship Award to R.G.