
Abstract
Short-lived turquoise killifish (Nothobranchius furzeri) becomes a popular model species for neuroscience. However, the effects of sex and rearing conditions on behavior and brain monoamines in N. furzeri are unknown. In this article, we study the body mass, behavior in the novel tank diving test, levels of noradrenaline (NA), dopamine (DA), serotonin (5-HT), 3,4-dihydroxyphenylacetic acid (DOPAC), and 5-hydroxyindoleacetic acid (5-HIAA) in the brain of 108 day-old N. furzeri males and females reared in small (one male and one or two females in 4-L tanks) and large (four males and four females in 25-L tanks) groups. Males were heavier and had a lower NA level in the brain compared with females. The behavior of males and females did not differ in the novel tank diving test. Their DA, 5-HT, DOPAC, and 5-HIAA levels in the brain did not differ too. Males from small groups spent more time near the tank's bottom. Rearing in small groups reduced the DA level in the female brain and the DOPAC level in female and male brains. However, group size did not affect body mass, 5-HT and 5-HIAA levels in the brain. Thus, group size is important for behavior and neuroscience studies of N. furzeri.
Introduction
Short-lived turquoise killifish, Nothobranchius furzeri, inhabit ephemeral ponds in southeastern Africa. Their fertilized eggs survive the dry season in diapause. The larvae hatch immediately after the pond is filled with water, grow rapidly, reach sexual maturity within 4–6 weeks, continuously mate and spawn during the wet season, and die at the age of 6–8 months.1,2 Males and females of N. furzeri differ at the morphological and behavioral levels. Males are larger than females and are brightly colored. They spend the majority of time guarding their territory from other males, attracting and mating females. 1
Six- to eight-month-old N. furzeri show morphological and behavioral hallmarks of aging, such as reduced coloration in males, emaciation, spinal curvature, spine and face malformations,1,3 reduced spontaneous, exploratory activities, 4 and impaired learning performance.5,6 The fast life cycle and the hallmarks of aging make N. furzeri a convenient model species in aging research.1–3
N. furzeri becomes a promising model species for neuroscience, psychopharmacology, and ecotoxicology.7–12 Behavioral variation in males and females of N. furzeri was investigated in the emergence, open field, habitat choice, life skill tests, 7 and diurnal activity. 8 In addition, also the impact of pharmaceutical exposure on diurnal activity 9 and aggressive behavior 10 in N. furzeri was studied.
Exploration and anxiety are two opposite adaptive motivations in novel (and potentially dangerous) environments. The former provides information to successfully cope with a novel situation, whereas the latter may prevent trauma or death. Behavior of an animal placed in a novel environment results from a conflict between exploration and anxiety. The novel tank diving test is the most frequently used test for the conflict between exploration and anxiety in zebrafish (Danio rerio). Specifically, activity at the bottom of the tank reflects risk-averse behavior or anxiety, whereas activity at the surface reflects risk-prone behavior.13,14 Behavior of N. furzeri in the novel tank diving test has not yet been investigated.
Brain monoamines, serotonin (5-HT), dopamine (DA), and noradrenaline (NA) are involved in the mechanisms of stress, locomotor activity, exploration, and anxiety in mammalians15,16 and fish.17–19 However, there is no information about the level and metabolism of these monoamines in the killifish's brain.
N. furzeri males conflict for territory and females; thus, to prevent the possible influence of such intermale conflicts, killifish are usually reared individually or in small groups, including one male and one or two females.20,21 Thoré et al. 22 found that killifish reared individually in 10-L tanks were larger and spent more time in the center of an open field compared with those reared in groups of 5 or 10 individuals in 10-L tanks. However, it remains unclear whether these changes result from differences in free living space or group size in se. Moreover, there is no information about the effects of group size on the killifish's brain.
The aims of this study are to investigate the effects of sex and group size on locomotor activity, exploration, anxiety, and brain monoamines in adult N. furzeri. Here, we intend to study the behavior in the novel tank diving test and levels of NA, 5-HT and its metabolite, 5-hydroxyindoleacetic acid (5-HIAA), DA and its metabolites, 3,4-dihydroxyphenylacetic acid (DOPAC), and homovanillic acid (HVA) in the whole brain of adult males and females of N. furzeri housed in groups of two to three and seven to eight individuals in small and large tanks, respectively.
Materials and Methods
Animals
The experiments were carried out on 25 males and 30 females of the N. furzeri of ZMZ1001 strain. The killifish hatched in our laboratory from the eggs received from the European Research Institute of Biology of Ageing (ERIBA, Groningen, the Netherlands) according to the published protocol. 20 The larvae were fed ad libitum three times per day with freshly hatched brine shrimps (Artemia salina) for the first 3 weeks after hatching. At the age of 14–28 days post-hatch, young fish were reared in a 50-L tank equipped with filter model XL-860 (Xilong). Young and adult killifish were fed ad libitum two times per day with frozen blood worms (Chironomus plumosus). To avoid competition for food, we added as many worms as fish could eat in an hour.
At the age of 4 weeks, 14 males and 18 females were placed in groups of 2 (male and female, 10 tanks) or 3 (male and 2 females, 4 tanks) into fourteen 4-L glass tanks (21 cm in length, 20 cm in depth, and 11 cm in width, water depth was 17.5 cm). The other 11 males and 12 females were placed in groups of 4 males/4 females, 4 males/4 females, and 3 males/4 females into three 25-L glass tanks (41 cm in length, 33 cm in depth, and 21 cm in width, water depth was 29 cm). The tanks volumes (4 or 25 L) were selected to guarantee a similar living space of 2–3 L per one fish in small and large groups. The fish were kept in these tanks until subjecting each individual fish to a novel tank diving test. Water in the tanks was constantly filtered and aerated with filters model 019 (Barbus), and its temperature was 27°C.
Each tank was equipped with a plastic plant of 6 cm (small tanks) or 14 cm (large tanks) and a dish (10 cm in diameter, 3 cm in high) filled with sand for spawning. Every day (at 17:00), the tanks were cleaned and 20% of water was substituted by tap water filtered through an expert hard filter (Barrier). The 12 h light/12 h dark (“light on” mode at 09:00) photoperiod was maintained.
All manipulations were carried out in accordance with the recommendations of the Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the protection of animals used for scientific purposes and was approved by the Committee on the Ethics of Animal Experiments of the Russian National Center of Genetic Resources of Laboratory Animals of Institute of Cytology and Genetics of Russian Academy of Sciences.
Novel tank diving test
When we started carrying out the test, the age of males and females was 100–108 days (Fig. 1). Since it was the first application of the novel tank diving test to killifish, we used the protocol of this test developed for zebrafish. 23 The test was conducted in the daytime (11:00 a.m. to 15:00 p.m.) in a glass test tank (24 cm in length, 15 cm in depth, and 7 cm in width, Fig. 2A). The test tank was placed into a wooden cabinet and illuminated with two photodiode lamps 5 W each with semitransparent light diffusers: one 40 cm above and another 10 cm behind the tank. This illumination provided the perfect contrast range between fish and background. The test tank was filled with 1680 mL of water at 27°C, 10 cm in depth. So, the water column was 24 cm in length, 10 cm in depth, and 7 cm in width (Fig. 2B).

Timeline of the experiment. dph, Days post-hatching.

Tracking killifish in the novel tank diving test by EthoStudio software.
The fish was individually transferred from its home tank into the test tank, then the tank together with the fish inside was placed into the cabinet, and the recording was started immediately. The water column (24 cm in length and 10 cm in depth) was virtually divided into the lower, middle, and upper thirds of 24 cm in length and 3.3 cm in height each (Fig. 2C). The fish position was automatically recorded for 5 min at the rate of 30 fps by C920 Pro HD Web camera (Logitech, Lausanne, Switzerland) connected to a computer (Windows 7) via a USB 2.0 port.
The stream of frames was automatically analyzed in real time by the EthoStudio software 23 and saved on a hard disk as a compressed video file. The EthoStudio software frame by frame separated the pixels associated with the fish from those associated with the background by applying the threshold algorithm23,24 and calculated the coordinates of the fish center (Fig. 2B). Moreover, the EthoStudio software automatically calculated the density map corresponding to the spatial distribution of fish-associated pixels in the tank (Fig. 2C).23,24
The sequence of coordinates and the density map were used to evaluate (1) the distance traveled (cm), (2) the immobility time (%), (3) the mean distance from the tank's bottom (cm), the time spent (%) in (4) the lower and (5) the upper thirds of the tank, and (6) the explored part of the tank (%). The fourth and fifth parameters were calculated as the ratios of the amount of fish-associated pixels in the lower and upper thirds of the tank, respectively, to the sum of fish-associated pixels in all three thirds.23,24 The last parameter was calculated as the ratio of amount of pixels visited by the fish to the total amount of pixels in the tank's water arena. 25 Each fish was recorded only once.
High performance liquid chromatography
Immediately after the novel tank diving test, the fish were euthanized by immersion into 0.1% tricaine methanesulfonate (Sigma-Aldrich) solution and then into cold water (+2°C) (Fig. 1); their bodies were immediately dried with dry napkins, and their masses were measured by using an Ohaus PA-512 electronic balance (Ohaus Corporation) with an accuracy of 10 mg. Then, their whole brains were immediately removed, frozen with liquid nitrogen, and stored at −80°C. The brain was homogenized in 100 μL of cold 0.6 M HClO4 by using a motor-driven grinder (Z359971; Sigma-Aldrich), and the homogenate was spun for 15 min at 12,000 rpm (+4°C).
The pellet was stored at −20°C until protein quantitation in accordance with the Bradford method, whereas the clear supernatant was diluted twice with pure water and used for assay of monoamines and their metabolites by high performance liquid chromatography (HPLC) on Luna C18(2) column (5 μm particle size, L × inner diameter 75 × 4.6 mm; Phenomenex) with electrochemical detection (750 mV, DECADE II™ Electrochemical Detector; Antec) and the glassy carbon flow cell (VT-03 cell 3 mm GC sb; Antec), system controller CBM-20 A, solvent delivery unit LC-20AD, autosampler SIL-20 A, and degasser DGU-20A5R (Shimadzu Corporation, USA). The mobile phase (pH = 3.2) contained 6.53 g of KH2PO4, 100 μL of 0.5 M Na2EDTA, 150 mg of 1-octanesulfonic acid sodium salt (Sigma), and methanol (13% volume; Vektor Ltd.). 26
The standard mixes containing 1, 2 and 3 ng of each NA, 5-HT, 5-HIAA, DA, DOPAC, and HVA were repeatedly assayed throughout the entire procedure and used to plot the calibration curves for each substance. The areas of peaks were estimated by using LabSolution LG/GC software version 5.54 (Shimadzu Corporation, Japan) and calibrated against the calibrated curves for corresponding standards. 26 The contents of NA, 5-HT, 5-HIAA, DA, DOPAC, and HVA were expressed in ng/mg protein assayed applying the Braford method and described elsewhere. All data are means of two replications.
Statistics
All data were tested by using the Kolmogorov's test and met the assumption of normality. Data were presented as the mean ± standard error of the mean and analyzed by two-way analysis of variance with “sex” and “group size,” including their interaction, as the independent factors. Post hoc analyses were carried out by using the Fisher's least significant difference multiple-comparison test, as appropriate. Statistical significance was set at p < 0.05.
Results
Effect of “sex” and “group size” on killifish behavior in the novel tank diving test
Marked effect of the “sex” factor was observed only regarding the body mass (Table 1). Males were heavier than females (Fig. 3). At the same time, no effect of the “group size” on body mass was shown (Table 1 and Fig. 3).
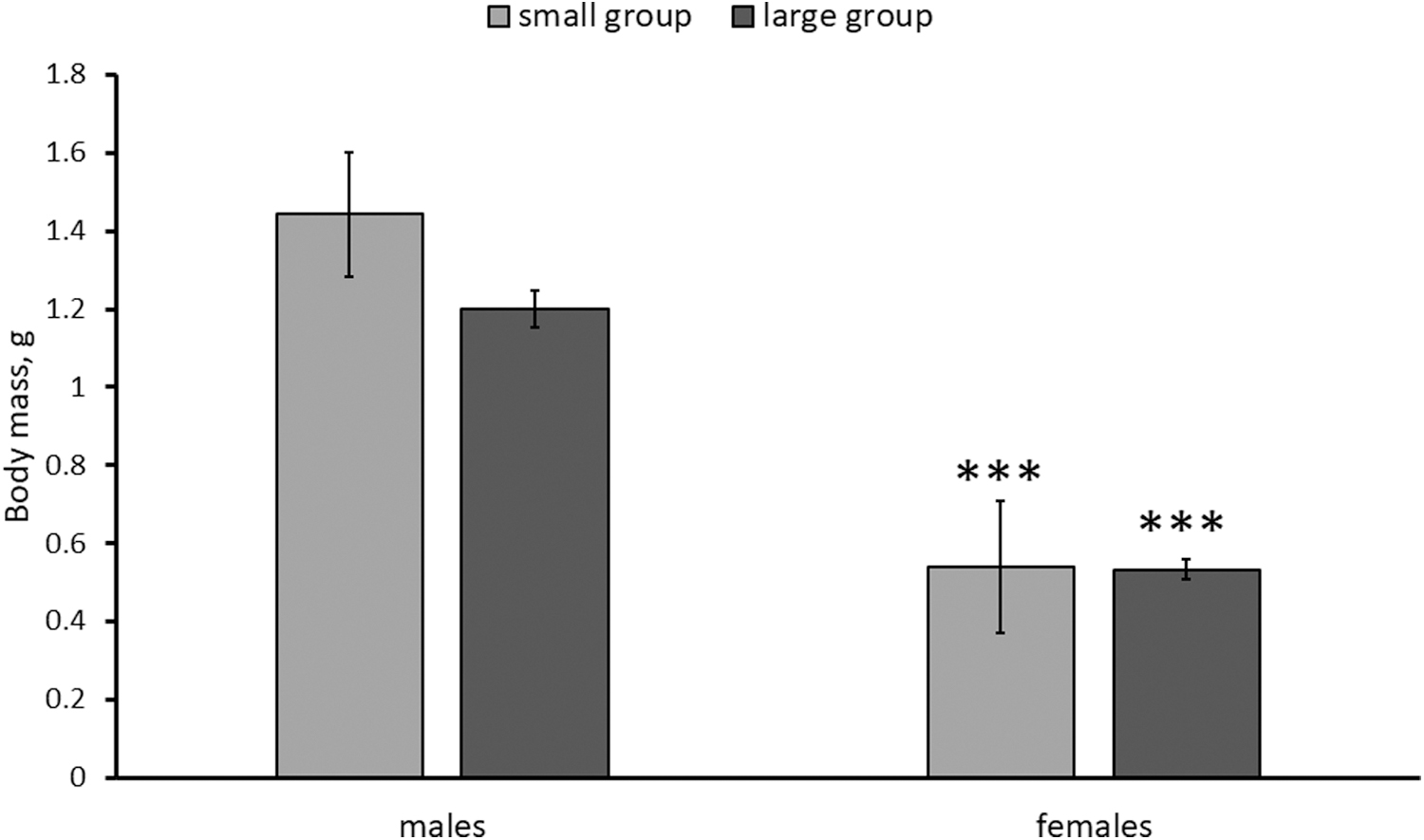
Body mass (g) in males and females of killifish reared in small and large groups. Numbers of males in small and large groups are 14 and 11, respectively. Numbers of females in small and large groups are 18 and 12, respectively. ***p < 0.001 versus males of corresponding groups.
Two-Way Analysis of Variance of the Effect of “Sex,” “Group Size,” and Their Interaction on the Variability of Body Weight, Distance Traveled, Immobility Time, Mean Distance from the Bottom, Time Spent in the Lower Third, Time Spent in the Upper Third, and Explored Part of the Tank in Males and Females of Nothobranchius furzeri Kept in Small and Large Groups
Significant values are marked in bold.
No effect of “sex,” “group size” and their interaction on the distance traveled, the immobility time, the explored part of the tank, and the mean distance from the tank's bottom was shown (Table 1). Males and females from small and large groups showed low locomotor activity (244 ± 48 cm males and 300 ± 45 cm females), remained immobile about 70% of the time near the tank's bottom, and explored less than 30% of the water arena (Fig. 4). At the same time, a significant effect of the “group size” factor on the time spent in the lower third was shown (Table 1). An effect of the “sex” × “group size” interaction on the time spent in the upper third was revealed (Table 1). Males from the large groups spent less time in the lower third and, correspondingly, more time in the upper third (Fig. 4).

Distance traveled (cm), immobility time (%), explored part of the tank (%), mean distance from the tank's bottom (cm), and time (%) spent in the lower and the upper thirds in the novel tank test in males and females of killifish reared in small and large groups. Numbers of males in small and large groups are 14 and 11, respectively. Numbers of females in small and large groups are 18 and 12, respectively. #p < 0.05, ##p < 0.01 versus males from small groups.
Effect of “sex” and “group size” on the levels of NA, DA, DOPAC, DOPAC/DA ratio, 5-HT, 5-HIAA, and 5-HIAA/5-HT ratio in the brains of killifish
Significant effect of the “sex” factor on the NA level has been shown (Table 2). This neurotransmitter concentration in the brain of females is higher compared with that in males (Fig. 5). A marked effect of the “group size” on NA, DA, and DOPAC concentrations in the brain was demonstrated (Table 2). The brain levels of DA were higher in females and those of DOPAC were higher in females and males from large groups compared with those from small groups (Fig. 6). The HVA level in the killifish brains was undetectable. No effect of “sex,” “group size” and their interaction on DOPAC/DA ratio, 5-HT, 5-HIAA levels, and 5-HIAA/5-HT ratio was observed (Table 2 and Figs. 6 and 7).
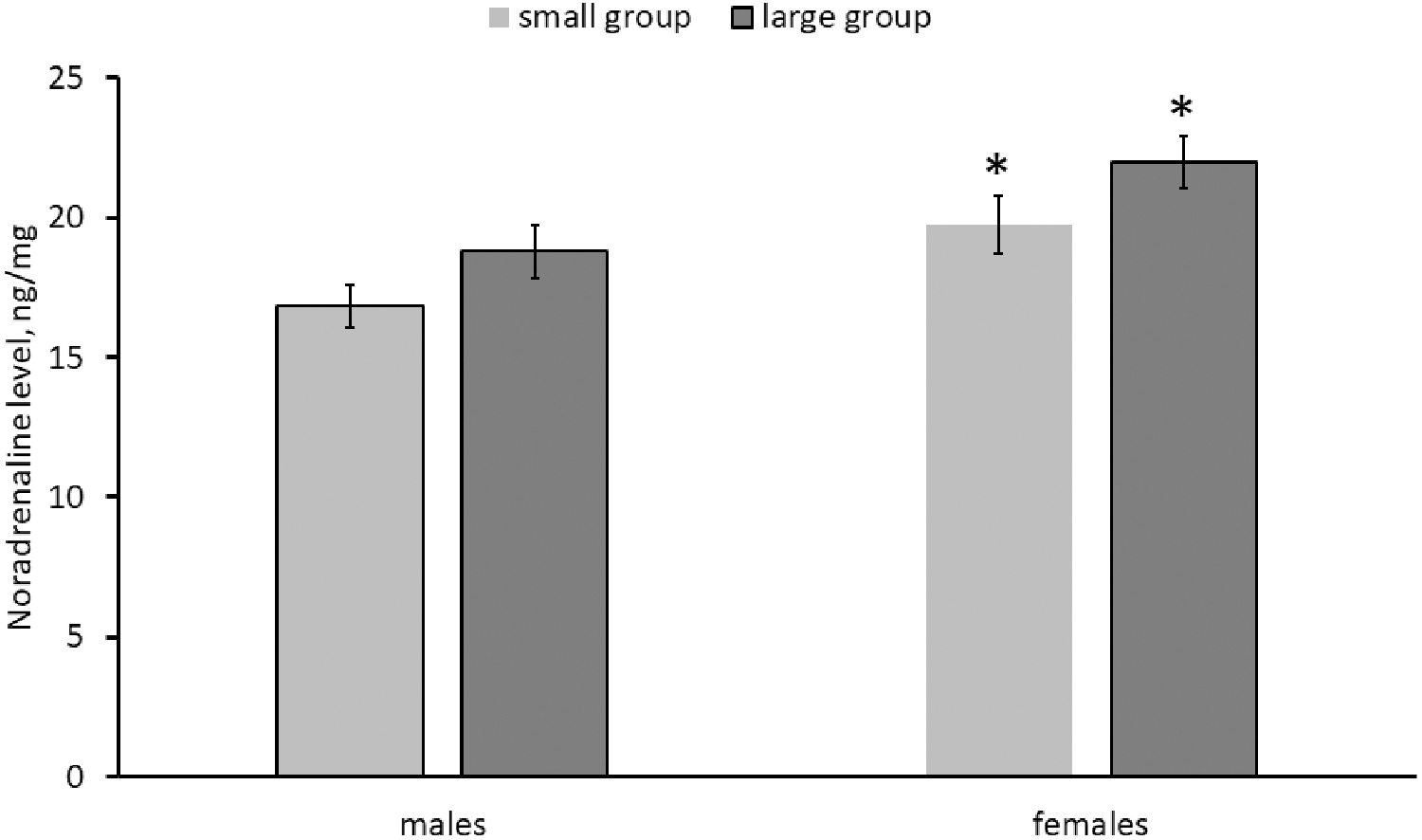
Noradrenaline levels in the brain of males and females of killifish reared in small and large groups. Numbers of males in small and large groups are 14 and 11, respectively. Numbers of females in small and large groups are 18 and 12, respectively. *p < 0.05 versus males of corresponding groups.
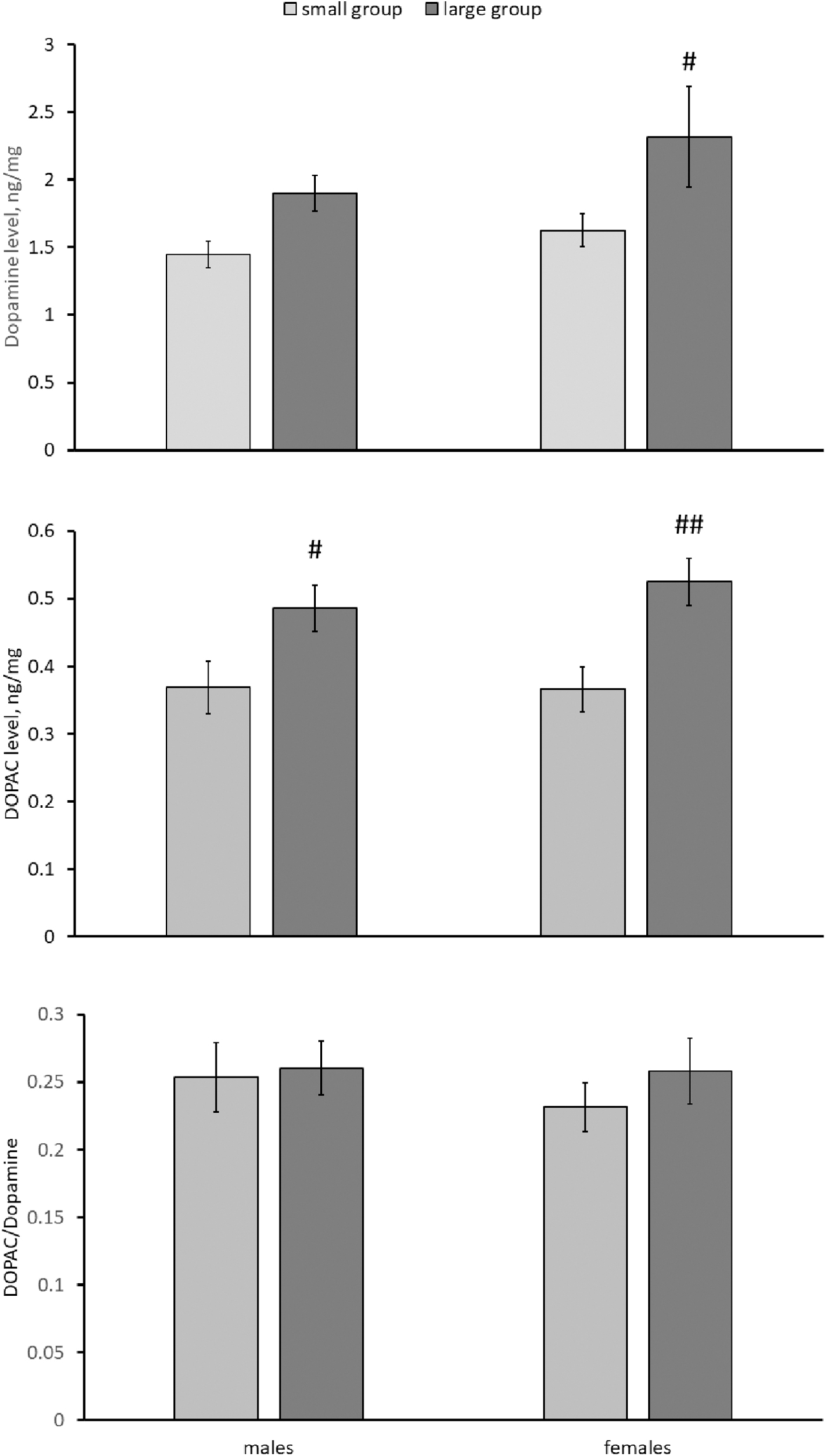
DA, DOPAC levels and DOPAC/DA ratio in the brain of males and females of killifish reared in small and large groups. Numbers of males in small and large groups are 14 and 11, respectively. Numbers of females in small and large groups are 18 and 12, respectively. #p < 0.05, ##p < 0.01 versus reared in small groups. DA, dopamine; DOPAC, 3,4-dihydroxyphenylacetic acid.
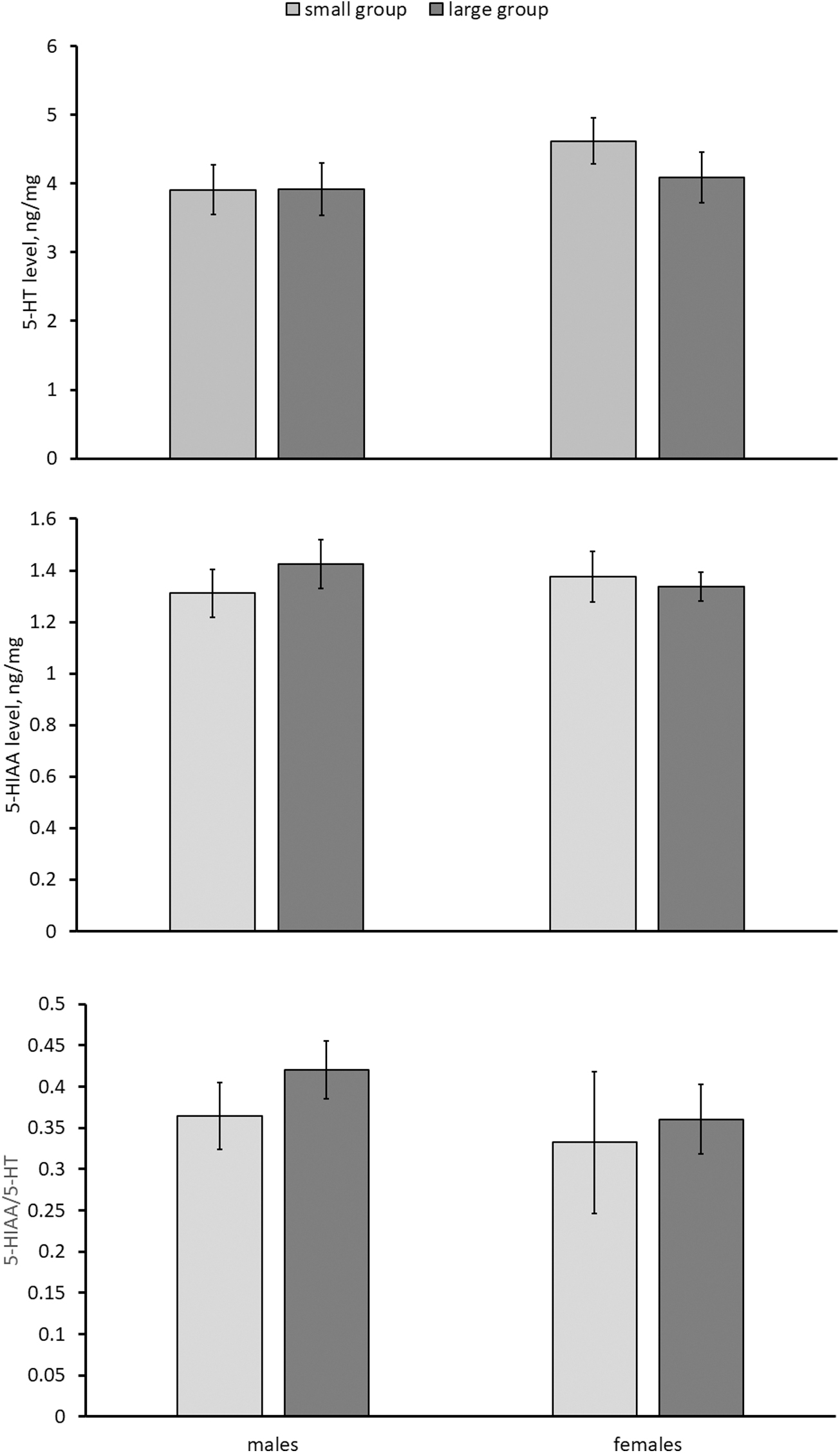
5-HT, 5-HIAA levels and 5-HIAA/5-HT ratio in the brain of males and females of killifish reared in small and large groups. Numbers of males in small and large groups are 14 and 11, respectively. Numbers of females in small and large groups are 18 and 12, respectively. 5-HIAA, 5-hydroxyindoleacetic acid; 5-HT, serotonin.
Two-Way Analysis of Variance of the Effect of “Sex,” “Group Size,” and Their Interaction on the Variability of NA, DA, DOPAC, 5-HT, 5-HIAA Concentrations as Well as on the DOPAC/DA and 5-HIAA/5-HT Ratios in the Whole Brain in Males and Females of Nothobranchius furzeri Kept in Small and Large Groups
Significant values are marked in bold.
5-HIAA, 5-hydroxyindoleacetic acid; 5-HT, serotonin; DA, dopamine; DOPAC, 3,4-dihydroxyphenylacetic acid; NA, noradrenaline.
Discussion
Since there was no information about killifish reaction to novel and potentially dangerous environments, the first aim of this study was to investigate the behavior of N. furzeri in the novel tank diving test as well as sex differences and the effect of rearing conditions on this behavior. This test is widely used to study locomotor, exploratory, and anxiety-related behavior in zebrafish,13,14 but until now it was not applied to killifish. Despite pronounced sexual dimorphism, no difference between the behavior of killifish males and females in the novel tank diving test was revealed. We found out that the mean traveled distance of males and females during a 5 min period was 244 and 300 cm, respectively. This result agrees with that of Thoré et al.,7,22 who showed that in the open field test killifish traveled about 1500 cm during 20 min (or about 380 cm during 5 min).
It should be noted that locomotor activity of N. furzeri in the novel tank diving test is four times lower compared with that in zebrafish: Earlier, we showed that zebrafish in this test traveled about 1200 cm during 5 min. 23
Both males and females spent most of their time in immobility near the bottom and practically did not explore the tank. This reaction of killifish to novelty looks like that of zebrafish placed in a novel tank that also spend more time near the tank's bottom.13,14,23 This preference of the tank's bottom in zebrafish is usually interpreted as a demonstration of anxiety toward novelty, since anxiolytics decrease the time spent near the bottom.13,14 However, we have no information that this observed preference of the tank's bottom in killifish is also fear reaction to novelty. In our facility, adult killifish in home tanks spent their time mainly near the bottom searching for their food–blood worms. Therefore, the behavior of killifish in the novel tank diving test may also result from their habit rather than anxiety. This question needs an additional experimental study.
Thoré et al. showed the effects of conspecific density on the size of N. furzeri: The killifish reared individually in 10-L tanks were larger compared with those reared in groups of 10 in 10-L tanks. 22 In our experiment, no effect of group size on body mass of males and females was found. This discrepancy seems to result from the difference in the experimental designs. In our experiment, the conspecific densities were similar in small (1 per 2 L) and large (1 per 2.5 L) groups, whereas in Thoré et al.'s experiment 22 the density in a small group (1 per 10 L) was 10 times less than in a large group (10 per 10 L). Thus, it seems that the density, but not the number of conspecifics affects the body size/mass in killifish.
Thoré et al. 22 also did not observe any effect of conspecific density on locomotor activity and velocity in killifish in the open field test. However, solitary killifish were more “bold” and they spent more time in the center of the open field test compared with those from high-density groups. 22 We also did not show any effect of group size on locomotor, exploratory activities, immobility time, and mean distance from the tank's bottom in the novel tank diving test. However, in this test, the males from large groups spent less time near the tank's bottom and correspondingly more frequently visited the surface compared with males from small groups. In experiments with D. rerio, a decrease in time spent near the tank's bottom is usually interpreted as an anxiolytic effect.13,14 However, additional information about the manifestation of anxiety in N. furzeri is necessary to correctly interpret the decrease in time spent near the tank's bottom in killifish males from large groups as an anxiolytic effect caused by an increase in the number of contacts and/or the tank size.
An alternative interpretation considers a possible influence of water depth on the vertical activity of N. furzeri males. Indeed, water depth in small tanks (17.5 cm) is less than in large tanks (29 cm) and the males reared in deeper water develop higher vertical activity to guard the tank's volume compared with males reared in shallower water.
Brain monoamines NA, DA, and 5-HT are involved in the mechanisms of stress reaction, locomotion, and anxiety.15–17 An agonist of the 5-HT1A receptor, buspirone, decreases the time spent near the bottom in the novel tank diving test as well as the time spent in the dark compartment in the light/dark test in zebrafish. 19 An inhibitor of 5-HT transporter, fluoxetine, increases surface dwelling and decreases locomotor activity in zebrafish.19,23 Therefore, the second aim of the present study was to investigate the metabolism of NA, DA, and 5-HT in the brain of N. furzeri as well as sex differences and the effect of rearing conditions on the expression of these neurochemical parameters.
We first revealed a significant increase in the NA level in the brain of females compared with males in killifish. At the same time, no sexual difference in the level of 5-HT, 5-HIAA, DA, DOPAC, as well as DOPAC/DA and 5-HIAA/5-HT ratios in the killifish's brain was observed. Earlier data on increase in the DA level and decrease in the 5-HIAA/5-HT ratio in the forebrain of female zebrafish compared with males were published. 27
For the first time, we have revealed the effect of the group size on DA and DOPAC levels in the brain. The levels of these substances were higher in killifish reared in large groups compared with those reared in small groups. At the same time, no effect of group size on 5-HT, 5-HIAA levels as well as on 5-HIAA/5-HT and DOPAC/DA ratios was observed. Unfortunately, we cannot explain this increase in DA and DOPAC levels in the brains of killifish kept in large groups due to lack of information about the possible role of this monoamine in killifish behavior.
As noted earlier, in natural systems, N. furzeri males conflict for territory and females. To prevent a possible effect of this conflict on the brain and behavior of killifish, males are usually reared individually or in small groups, including one male and one or two females. In the present study, we have shown that rearing in small groups affects the behavior in the novel tank diving test and the brain DA system in killifish males. At the same time, the group size does not produce any effect on the body mass or locomotor activity of killifish males. Thus, the rearing conditions can significantly affect the behavior and DA system in the brain of killifish and their influence must be taken into account in the study of behavior and neurochemistry in this species.
Conclusion
Turquoise killifish becomes a popular laboratory species in behavioral and neuroscience studies. We did not show any sexual dimorphism in reaction to novelty in the novel tank diving test. At the same time, NA concentration in the brain of females was higher than in males.
The most interesting and particularly important obtained results are related to the effects of rearing conditions on killifish behavior and brain monoamines. Rearing in small groups including one male and one or two females in 4-L tanks increases the time spent near the tank's bottom and correspondingly decreases the frequency of surface visits, as well as decreases the brain DA and DOPAC levels compared with killifish reared in large groups, including four males and four females in 25-L tanks. This result indicates that rearing conditions are important for killifish and they need to be considered when planning behavioral and neurobiological research.
Footnotes
Acknowledgment
The authors are grateful to Dr. Eugene Berezikov, the group leader of the Laboratory of Stem Cell Regulation and Mechanisms of Regeneration of the ERIBA for kindly providing the eggs of N. furzeri.
Disclosure Statement
No competing financial interests exist.
Funding Information
The studies are supported by the budget project (No. 0259-2019-0002) and implemented using the equipment of the Center for Genetic Resources of Laboratory Animals at ICG SB RAS, supported by the Ministry of Education and Science of Russia (Unique identifier of the project RFMEFI62119X0023).