
Abstract
In this study, an experimental protocol has been developed for comparing survival rates of mutant and wild-type zebrafish larvae under extreme starvation. Zebrafish larvae were placed in 96-well plates at fourth day postfertilization (dpf) and larvae were not fed at all from hatching to cease. Zdf1 zebrafish line was used, a strain carrying the (cancer) pathogenic TP53-M214K amino acid substitution. TP53-M214 corresponds to the human TP53-M246 and both residues are located on the DNA-binding domain of the p53 protein. Survival statistical analysis did not show any significant difference in the overall survival rates between homozygous mutant and wild-type larvae. When considering 15 dpf as the endpoint of the experiment (66% of larvae died), a borderline statistical significance was observed for the dominant model of inheritance (p = 0.015; relative hazard = 0.8320). Despite the fact yolk sac of larvae is depleted at 7–8 dpf, 34% of larvae survive until 15 dpf and 1.5% until 17 dpf. Concluding, three main results derive from this study: (1) pathogenic homozygous mutations in TP53 probably do not alter survival rates of zebrafish larvae under starvation; (2) zebrafish larvae can live up to 17 dpf without food, surviving only with their initial nutritional supplies; and (3) an easy and affordable protocol has been developed for estimating survival rates of zebrafish larvae under stressful conditions.
Introduction
Starvation is a challenge that many living organisms must anticipate in nature. Published evidence supports that starvation and other stress types can induce cell apoptosis.1–5 P53 protein is a major player of apoptosis, and it is considered as the most important tumor suppressor protein. Studies show that reduced apoptosis potential can help some mammals, such as Spalax ehrenbergi, Myospalax baileyi, and Microtus oeconomus, to survive under stressful environments (Table 1).6,7 Reduced apoptosis potential in those animals is due to certain amino acid residues in the p53 protein.
Published Research Evidence for Association of TP53 (or TP53 Orthologs) with a Survival/Evolutionary Advantage
TP53 knockout mice have high incidence of cancer.
These residues are found in a highly conserved DNA-binding domain of TP53. Zhao et al. found evidence that the 104 amino acid residue of p53 in M. baileyi and M. oeconomus is needed for divergent responses of IGFBP3 and Apaf1 to hypoxia and cold stresses. 6 Ashur-Fabian et al. 7 found similar evidence for the amino acid position 174 in S. ehrenbergi. This amino acid is reported to be affected in 57 different human tumors of various types. Cell death resistance may be a shield for animals living under adverse environmental conditions. In addition, TP53 mutations have been associated with a survival advantage or generally a positive effect in many other species such as drosophila, mice, C. elegans, and humans (Table 1).
Extreme starvation stress has not been yet studied in zebrafish larvae. The aim of this study was double. First, we investigated if a pathogenic mutation in TP53 gene, the most important gene in apoptosis and cancer, can affect survival rates in zebrafish larvae under starvation. We selected zdf1 strain, a well-studied TP53 mutant fish line carrying a single amino acid substitution (TP53-M214K). This strain has been studied extensively as a cancer animal model.8–10 We selected this strain since it is the only one being available in EZRC (European Zebrafish Resource Center) and not carrying a frameshift mutation but a missense one.
In addition, TP53-M214K is lying on a DNA binding domain of the protein, where evidence exists for selection pressures on those domains.6,7 Zdf1 fishes predominantly develop peripheral nerve tumors, in contrast with the TP53 knockout zebrafish strains that additionally develop angiosarcomas, germ cell tumors, and an aggressive Natural Killer cell-like leukemia. 11 TP53-M214 corresponds to the TP53-M246 residue of the human orthologue gene. Both residues are located on the DNA-binding region of the p53 protein. Four missense pathogenic mutations have been identified so far at the human TP53-M246 residue, all related with familial cancer cases (Human Gene Mutation Database).
Second, this is the first time that survival rates have been estimated for zebrafish larvae under extreme starvation. Surprisingly, our results show that zebrafish larvae can live much more after depletion of their yolk sac.
The approach used in this study is easy and affordable, and it can be used for estimating survival rates under stress for other mutant lines of zebrafish. It can also be developed more, establishing the first vertebrate experimental evolution protocol.
Materials and Methods
Ethics statement
This study has been approved by the Veterinary Services of the Ministry of Agriculture, Rural Development and Environment of Cyprus, under the license number CY/EXP/PR.L8/2018. The zebrafish (Danio rerio) experiments took place during the period of December 2018–December 2020, at the Medical School of University of Cyprus. The animal experiments have been performed or supervised by KV, certified for handling zebrafish by The International Zebrafish and Medaka Course (IZMC) at Karlsruhe Institute of Technology (KIT), accredited by FELASA (Federation of Laboratory Animal Science Associations). The zebrafish strains were purchased from EZRC.
Following the 3Rs principle for animal use in research, we calculated the number of animals needed for this study before proceeding to our research protocol. According to Dell et al., 12 it was calculated (Fleiss equation) that ∼624 zebrafish larvae are needed for this study. The simulation was performed by assuming two different genotyping groups, aiming at finding a minimum of survival rate difference of 0.1 between the two groups. Power of study was set to (1-β) = 0.8 (80%). We finally used 591 zebrafish larvae (see Results and Discussion section) after performing two successful zebrafish breedings.
Fish husbandry
Zebrafish were maintained in a ZebTEC Stand-Alone with “Active Blue” Technology recirculating system (Techniplast, Italy). This system maintains automatically standard husbandry conditions, those set to be 28.5°C, 7.5 pH, and 550 μS conductivity. Photoperiod was set to 14 h light and 10 h dark.
Experimental design
The main goal of the experiments of this study was to test for any differences in survival percentages between TP53 mutant larvae and wild-type larvae under starvation (S) conditions. All zebrafish strains were purchased from the EZRC. Experiments were performed by breeding five zdf1 pairs (received from EZRC as adults), combining five zdf1 heterozygous females and three zdf1 heterozygous males in each breeding. The expected Mendelian percentages of the offspring are 25% wild type, 50% heterozygous, and 25% mutant (homozygous). Zdf1 zebrafish carry the pathogenic mutation TP53-M214K. 8
A supplementary experiment was performed (Sm), investigating for any difference in survival rates between heterozygous TP53-M214K larvae and wild-type larvae. Breeding involved two zdf1 heterozygous females and one wild-type WIK male. This experiment investigated for any maternal effect of the TP53-M214K mutation.
Breedings were performed in appropriate breeding tanks (Techniplast, Italy) using system water and a room temperature of 28°C. Embryos were collected in the morning 2 h after first light and were placed in system water, containing 0.00005% methylene blue, in 200 mL open beakers. At fourth day post-fertilization (dpf), the larvae were placed in 96-well sterile plates (Thermo Scientific, BioLite 96 Well Multidish, Korea), one larva in each well, in system water with final concentration of 0.00005% methylene blue. The 96-well plates were placed on a bench at a room temperature of 27°C–28°C. Every morning, one-third of water of each plate well was changed with fresh system water, doing this daily until the end of the experiment.
Mortality was checked every morning (every 24 h) until all larvae of the plate had died. Photoperiod of the room was 14 h light and 10 h dark. Larvae were not fed at all until the end of the experiment.
Mortality of zebrafish larvae was evaluated under a stereoscope (Motic SMZ-161-TLED with Moticam BTU8 Tablet Camera, China), taking images of representative larvae. Dead larvae were considered those not moving after 1 min of observation, touching them gently with a pipette tip. Dead larvae were placed in appropriately labeled tubes for DNA extraction.
Larvae DNA extraction and genotyping
Larvae DNA extraction and fin-clip DNA extraction was based on the method described by Meeker et al. 13 In brief, the protocol includes 20 min incubation at 95°C with 100 mM NaOH, following by adding of 10 μL of 1 M Tris-HCl. Centrifuging follows and 3.5 μL of supernatant is used for polymerase chain reaction (PCR). The DNA extraction method is very fast, and hundreds of samples can be handled in a very short time. DNA yield is between 0.2 and 1.2 ng/μL (260/280 nm ratio: 1.7–1.8), thus a highly sensitive DNA polymerase was used, TopTaq DNA polymerase (Qiagen, Germany).
TP53-M214K mutation was genotyped by a polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) method. Primers Zdf1-For-b 5-CCAGAGTATGTGTCTGTCCATCTGT-3 and Zdf1-Rev-b 5-CTGAACTGTTTCGGATAGCCTAGT-3 were used, giving a 335 bp PCR product. PCR conditions in detail are available under request. Restriction enzyme MboII (New England Biolabs) recognizes the mutation (A allele) and cuts the PCR product in two fragments, 206 and 129 bp, after incubation at 37°C. PCR product of the wild-type allele (T allele) remain uncut after incubation with MboII (Supplementary Fig. S1).
PCR digestion reactions were evaluated in 3% agarose gel electrophoresis. The genotyping method was confirmed by DNA resequencing of representative PCR products. Sanger sequencing was performed in ABI PRISM™ 3130 genetic analyzer (Applied Biosystems) (Supplementary Fig. S2). Fin-clip was performed for parental fishes to confirm their genetic status.
PCR-RFLPs took place at the premises of the “Center of Excellence in Biobanking and Biomedical Research” of University of Cyprus.
Statistical analysis
Statistical analysis and Kaplan–Meier graphs were performed by the statistical package STATAv.13 (StataCorp LLC, Texas). Equality of survivor functions was checked by the log-rank test (Table 2). Survival analysis was performed for the dominant and the recessive genetic model as well. Significant level alpha was set to 0.01.
Survival Statistical Analysis for Zebrafish Larvae Exposed to Starvation
Analysis was performed for the Dominant (Dom.) and the Recessive (Rec.) model as well, per breeding event and totally. S experiment involves breeding of zdf1 heterozygous parents. Sm experiment involves breeding of zdf1 heterozygous mothers and wild-type fathers.
Genetic models (m: TP53-M214K).
Rec.: [m/m] versus [m/+, +/+].
Dom.: [m/+, m/m] versus [+/+].
NA, not applicable.
Results and Discussion
Totally 591 zebrafish larvae were successfully genotyped, having been exposed to starvation at 27°C–28°C in 96-well plates. Larvae were not fed at all, from hatching to cease. Offspring were the result of Zdf1 breeding. Zdf1 is the zebrafish line carrying the heterozygous TP53-M214K mutation.
Genotypic counts
The offspring genotypic ratios were the expected Mendelian ones. Percentage of homozygous TP53-M214K larvae is 28%, slightly above the expected 25%. This is not a statistically significant divergence, but it may be a sign of increased survival of TP53 homozygous larvae. This observation needs further investigation.
Larvae lifespan under starvation
Representative larva images of the three genotypic statuses (TP53-214M/M, TP53-214M/K, and TP53-214K/K) are presented in Figure 1. Not any obvious phenotypic differences are observed.

Images of zebrafish larvae exposed to starvation (plate S11). Not obvious phenotypic differences exist among the different genotypes. Yolk sac is depleted at 7 dpf (red arrows). Mobility of larvae is significantly reduced after 14 dpf. dpf, day post-fertilization, m: TP53-M214K mutation, +: wild type.
Under starvation, yolk sac of zebrafish larvae is depleted at 7–8 dpf (Fig. 1). Despite this, some larvae survive 10 more days after yolk sac depletion. The maximum observed lifespan for larvae coming from zdf1 strain breedings is 17 dpf (Fig. 2). Massive death events begin at 14 dpf. Mobility of larvae is significantly reduced after 14 dpf. Interestingly, one wild-type larva remained remarkably active until 17 dpf (Supplementary Video S1), representing a percentage of ∼0.15%. Probably some larva can control much better their nutritional supplies than others.

Genotypic survival curves for the starvation S experiment.
According to our knowledge this is the first time that the maximum lifespan of zebrafish larva (probably of any fish larva) is reported under starvation in laboratory conditions. We conclude that fish larvae are very resistant to environmental starvation. Obviously, this increases odds of survival in nature.
A main weakness of this study is that the survival potential of zebrafish larvae exposed to extreme starvation over 10 dpf has not been investigated. However, we believe that it is possible for a small percentage of zebrafish larvae to survive until adulthood if they start eating after 10–13 dpf of starvation. We based this notion on the fact that most zebrafish larvae are quite active until 14 dpf. Even in case that 1% of the starved larvae survive in nature, this is evolutionarily significant. That means, for example, that 10,000 larvae could survive out of 1,000,000, a not negligible number.
Starvation survival
Analytical survival statistics per breeding and totally can be found in Table 2.
Combining the two breeding events of the S experiment, only 34% of larvae survived by the 15 dpf and just 1.5% by the 17 dpf (maximum observed lifespan). Not any statistical significance was observed between mutant and wild-type larvae, under the recessive or the dominant model (Table 2 and Fig. 2). However, when considering 15 dpf as the endpoint of the experiment (66% of larvae died), a borderline statistical significance was observed (p = 0.015, log-rank test) for the dominant model of inheritance (relative hazard = 0.8320) (Table 2). A potential positive effect for TP53-M214K cannot be excluded at 15 dpf or earlier. Investigation of more TP53 mutations could give more useful evidence.
Results of the Sm experiment show not any significant effect of the TP53 mutation when this is inherited from the mothers (Table 2 and Fig. 3). This probably shows that maternal origin of the mutation is not of any significance regarding survival.

Genotypic survival curve for the starvation Sm experiment. Confidence intervals are illustrated as well. About half of the larvae of this experiment are heterozygous and the other half are wild type, all coming from heterozygous mothers and wild-type fathers. Maternal origin of the TP53 mutation has not any significant effect in the survival rates. m: TP53-M214K mutation; +: wild-type status; RH: relative hazard.
The results of the starvation experiments do not show any significant advantage or disadvantage for the TP53-M214K mutation in the overall survival under those adverse laboratory conditions.
Evolutionary experimental protocol
Simulating vertebrate evolution in the laboratory is challenging. We believe that our study represents the first of such approach, a method that could be developed more by other teams. The protocol is inexpensive and easy, summarized in Figure 4 and described in detail in Materials and Methods section. This method can be modified accordingly for testing other environmental stressors, such as low/high pH, oxidative stress, and high temperature.
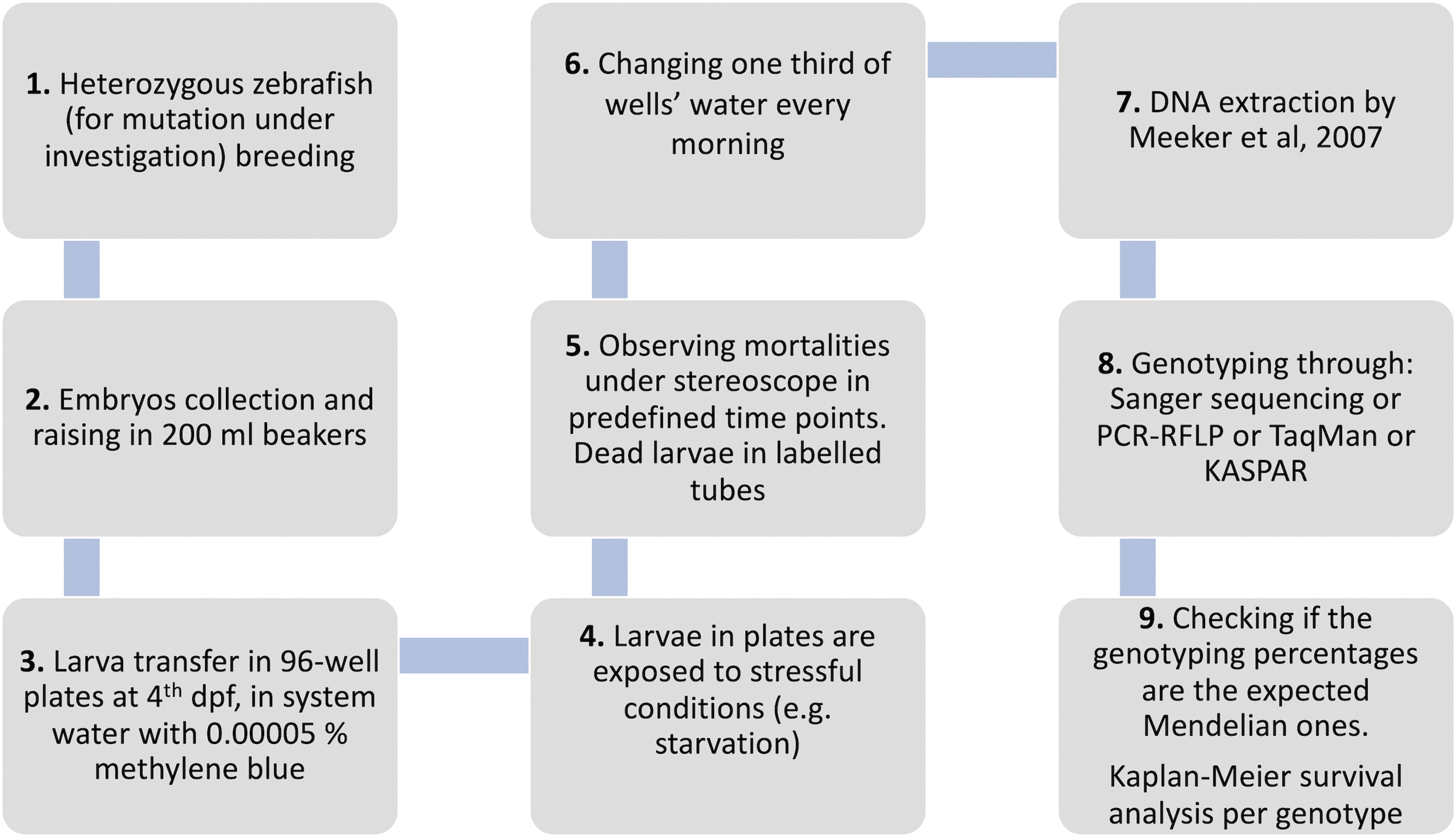
Experimental protocol developed for exposing zebrafish larvae to starvation stress.
Conclusions
Cancer incidence is varying across animal kingdom. Different explanations have been proposed.14–18 Natural selection implicating cancer genes under challenging environments could be part of this complicated equation. TP53 mutations have been associated with positive effects in a variety of species (Table 1). In this study, not any benefit or a negative effect has been observed for a TP53 mutation in homozygosity in zebrafish larvae under extreme starvation conditions. However, we showed for the first time that zebrafish larvae can live up to 17 dpf without food, surviving only with their initial egg nutritional supplies.
Footnotes
Authors' Contributions
K.V. had the initial idea of the project, established the zebrafish facility, designed the study, performed most of the laboratory experiments, performed the statistical analysis, constructed the figures, and wrote the first draft of the article. MP performed part of the DNA extraction and contributed to the assessment of the larva survival. C.K. performed part of the PCR-RFLP genotyping and she also performed the DNA resequencing. All the authors read and approved the submitted article.
Acknowledgment
We thank Prof. Constantinos Deltas for allowing the use of the premises of the “Center of Excellence in Biobanking and Biomedical Research” (University of Cyprus) for performing PCR-RFLP analysis.
Disclosure Statement
No competing financial interests exist.
Funding Information
There is no special funding for this research study. Laboratory consumables of this project are through the article 3/346 of University of Cyprus.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.