
Abstract
Role of telomerase in specimens with retarded growth (dwarfs) has not been thoroughly examined to date. Considering that some of the fish species show correlation between somatic growth and activity of telomerase, it has been tempting to assume that pattern of telomerase activity in specimens with retarded growth and these with normal growth rate may vary. In the present research, telomerase activity has been examined in liver, skin, and muscles in the androgenetic rainbow trout (Oncorhynchus mykiss) with growth deficiency and their normally developed siblings. Among the examined organs, the liver showed the highest telomerase activity in all studied fish, what may be linked to the enormous regeneration capacity of the liver tissue. Although dwarf specimens examined here displayed significantly lower body size and weight they did not exhibit any significant differences in the telomerase activity measured in liver and muscle when compared to the rainbow trout without growth deficiency. In turn, telomerase activity in skin was significantly upregulated in the normally developed androgenotes. The present study indicates that dwarfism in the androgenetic rainbow trout is neither associated with ceased telomerase activity nor its decrease throughout the ontogenetic development.
Introduction
Telomeres are eukaryotic terminal chromosomal regions composed of tandemly repeated DNA sequences associated with specific proteins. In vertebrates, telomeric DNA sequence is TTAGGG motif that extends in a 5′ to 3′ direction from double-stranded DNA to a single-stranded overhanging region. 1 Telomeres protect chromosomes from degradations and fusions,2,3 ensure proper chromosome topology in the nucleus, and are able to silence expression of genes located adjacent to the telomeric region.4,5 Telomeres shorten after each round of the cell division and this loss may be compensated by telomerase, a ribonucleoprotein enzyme composed of telomerase reverse transcriptase subunit (TERT) that adds telomeric DNA sequences to the 3′ end of telomeres using an integral RNA component (TR, telomerase RNA) as a template. 1
Telomerase has been detected in all eukaryotes including vertebrates, however, its activity varies sometimes considerably between species. Generally, in mammalian species including humans telomerase is mostly absent or extremely downregulated in the somatic tissues/cells and its expression is observed exclusively in the highly proliferating cells including germ line cells and stem cells.6,7 Moreover, upregulated telomerase has been reported in more than 90% of cancers and even up to 99% of the tumor originated cell lines.8–10
In contrast to humans, telomerase in mice, 11 birds, 12 fish,13–19 and in the aquatic invertebrates14,20 is active in the somatic tissues. The pioneer research on rainbow trout (Oncorhynchus mykiss) evidenced significant levels of telomerase activity in the kidney, liver, skin, heart, muscle, and brain irrespective of the fish age and size. 13 Studies on several model fish species exhibit a high correlation between TERT gene expression and variation in the telomerase activity, suggesting that telomerase activity in fish might be primarily regulated at the transcriptional level and not at the protein modifications stage.18,19
Observed levels of telomerase activity in fishes are mainly age and size dependent, usually displaying temporal increase during early developmental stages that are characterized by the fast somatic growth and accelerated cell proliferation.16,21 Upon maturity, telomerase activity varies between species and organs and can decrease, sustain, or increase.14,15,19,21 Levels of the telomerase activity in particular tissues are largely species-specific and reflect differential proliferative demands of cells. For instance, in zebrafish, platyfish (Xiphophorus maculatus), and in Nothobranchius furzeri lower activity of telomerase is detected in organs with the differentiated cells that are at the post mitotic state, such as brain and higher activity is reported in the undifferentiated cells for example, in gonads.21–23
Taking into account that in some of the fish species correlation between somatic growth and activity of telomerase in the somatic tissues has been observed,16,19 it is tempting to assume that pattern of telomerase activity in the specimens with growth deficiency (like dwarfs) and those with the normal growth rate may vary. Dwarfism is a condition characterized by body size reduction that occurs in humans 24 and in animals 25 including fishes.26–28 Dwarfism may be caused by genetic disorders (achondroplasia), growth hormone deficiency, or malnutrition.24,29 Individuals with the growth retardation may have reduced but proportionate body or reduced body size may be accompanied by the skeletal abnormalities. Individuals with growth deficiency and body deformations have been observed among fish from wild stocks and from aquaculture. 30 Remarkably increased ratio of dwarf specimens has been reported among homozygous doubled haploids (DHs) produced by induced androgenesis and mitotic gynogenesis in salmonid fish species.31–33
As growth retardation makes dwarf fish interesting objects to study the roles of genetic and physiological factors responsible for this issue, the main goal of the present research was to examine activity of telomerase in androgenetic rainbow trout with growth deficiency and normally developed individuals.
Materials and Methods
The study was carried out in strict accordance with the recommendations in the Polish ACT of January 21, 2005 of Animal Experiments (Dz. U. of. 2005 No 33, item 289). The protocol was approved by the Local Ethical Committee for the Experiments on Animals in Bydgoszcz.
Fish origin and sampling
Studied rainbow trout originated from the spring spawning heterozygous Rutki strain and from the homozygous DHs strain produced within induced androgenesis using gamete donors originated from the Rutki strain (Fig. 1). 34 Fish were reared under routine conditions in the Department of Salmonid Research (DSR), Inland Fisheries Institute in Olsztyn (IFI), Rutki, Poland.
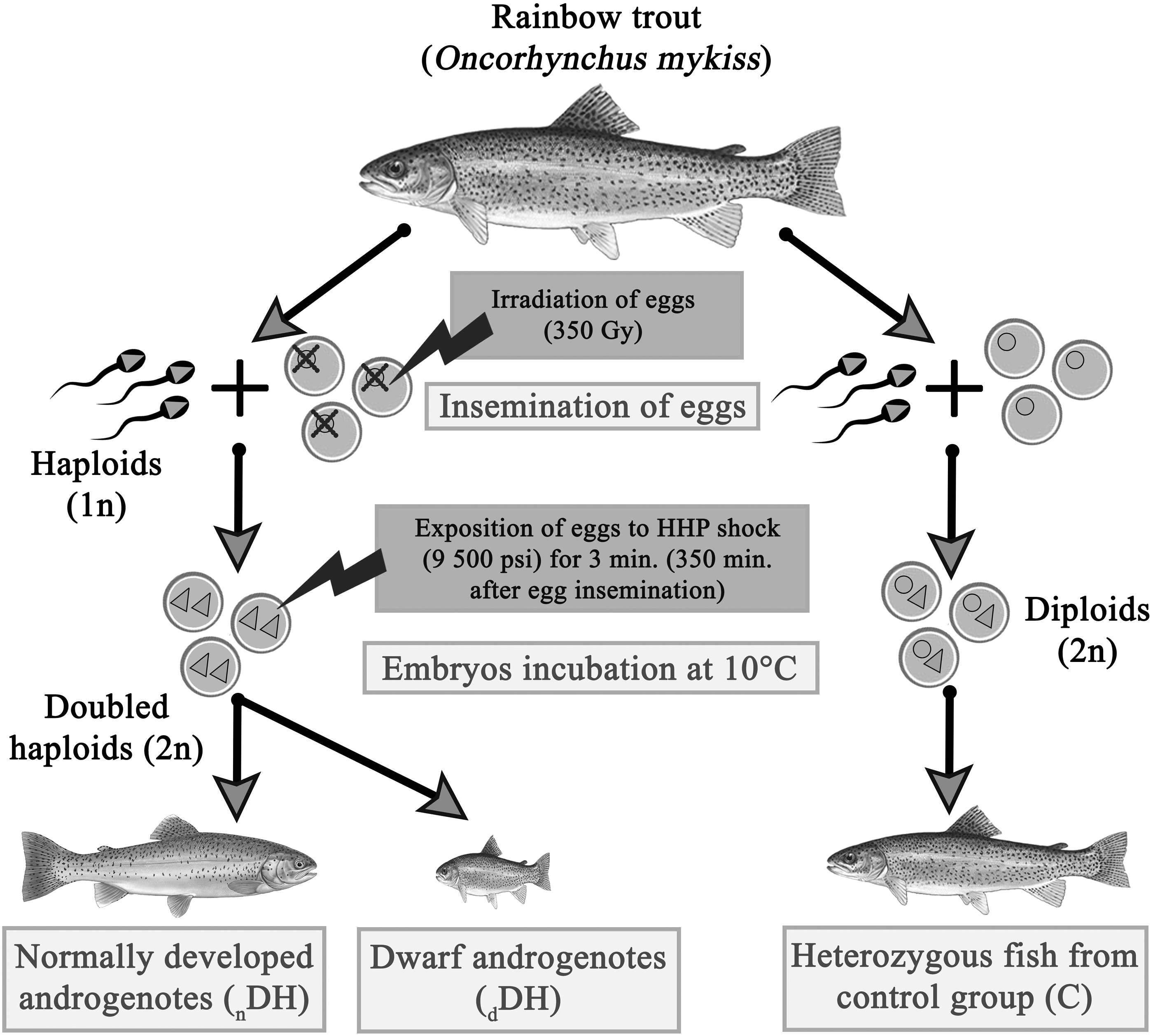
The graphical summary of artificial androgenesis induced in the rainbow trout (Oncorhynchus mykiss). HHP, high hydrostatic pressure.
In total, fifteen 1-year old rainbow trout including five androgenotes with growth deficiency (dwarfs) (dDH), five normally developed androgenotes (nDH), and five heterozygous individuals from the control group (C) were sampled on July 20, 2020. Fish were sacrificed by overdose of the clove oil in a dose of 150 mg/L and their body weight and length were measured. Next, 100 g of liver, muscles, and skin were collected from each fish and immediately used for further analysis.
Analysis of telomerase activity
The telomerase activity assay was carried out in at least two replications for all collected tissues from the examined fish specimens. The analysis was performed using TeloTAGGG Telomerase PCR ELISA Kit (Roche Diagnostics GmBH, Mannheim, Germany) according to the manufacturer's instructions. The assay includes positive control, which is human, telomerase-positive embryonic kidney cell line 293. This technique is based on highly specific amplification of telomerase-mediated elongation products (TRAP) combined with nonradioactive detection following ELISA protocol. 35 The TRAP method is based on two-steps amplification, where at first telomerase adds telomeric repeats (TTAGGG) to the 3′ end of the biotin-labeled synthetic P1-TS-primer. Then the elongation products are amplified by PCR using the primers P1-TS and P2, generating PCR products with the telomerase-specific 6 nucleotide increments.
Under the ELISA assay, the obtained PCR products were denatured and hybridized to a digoxigenin-(DIG)-labeled, telomeric repeat-specific detection probe. Hybridized products were immobilized via biotin labeled primer to a streptavidin-coated microplate and detected with an antibody against digoxigenin (anti-DIG-POD) that was conjugated to peroxidase. An absorbance of colored reaction products was measured at 450 nm by a microplate reader (SpectraMax i3; Molecular Devices), using as a reference wavelength of 690 nm.
Statistical analysis
Data were analyzed using R software version 1.3.959. Normal distribution was tested by the Shapiro–Wilk test, whereas Levane's test was used to determine homogeneity of variances between analyzed groups of rainbow trout. Student t-test or Kruskal–Wallis test (according to the data distribution) was used to determine whether there were any significant differences (p < 0.05) in telomerase activity between organs in fish from the examined groups.
Results
The mean values of body weight and length of normally developed androgenotes (nDH), androgenetic dwarfs (dDH), and heterozygous individuals from the control group (C) equaled 215.54 ± 36.25 g and 25.16 ± 1.33 cm, 76.46 ± 28.33 g and 18.66 ± 1.85 cm, and 184.82 ± 31.06 g and 26.08 ± 1.63 cm, respectively (Table 1). Dwarf androgenetic rainbow trout displayed significantly lower body length and weight when compared to normally developed androgenotes and rainbow trout from the control group (p < 0.05). Particular dwarf androgenotes had visible morphological deformations such as humpback, kyphosis, and lordosis (Fig. 2). There were no significant differences in the body size between normally developed androgenetic trout and fish from the control group.

Body size of dwarf
Body Weight and Length of Normally Developed Androgenetic Rainbow Trout (nDH), Dwarf Androgenotes (dDH), and Individuals from Control Group (C)
The liver was the organ with the highest level of telomerase activity that equaled 2.02 ± 0.55, 2.06 ± 0.36, and 2.52 ± 0.31 in normally developed androgenetic trout (nDH), androgenotes with reduced growth (dDH), and in the trout from the control group, respectively (Fig. 3). The differences in the telomerase activity were insignificant (p < 0.05).
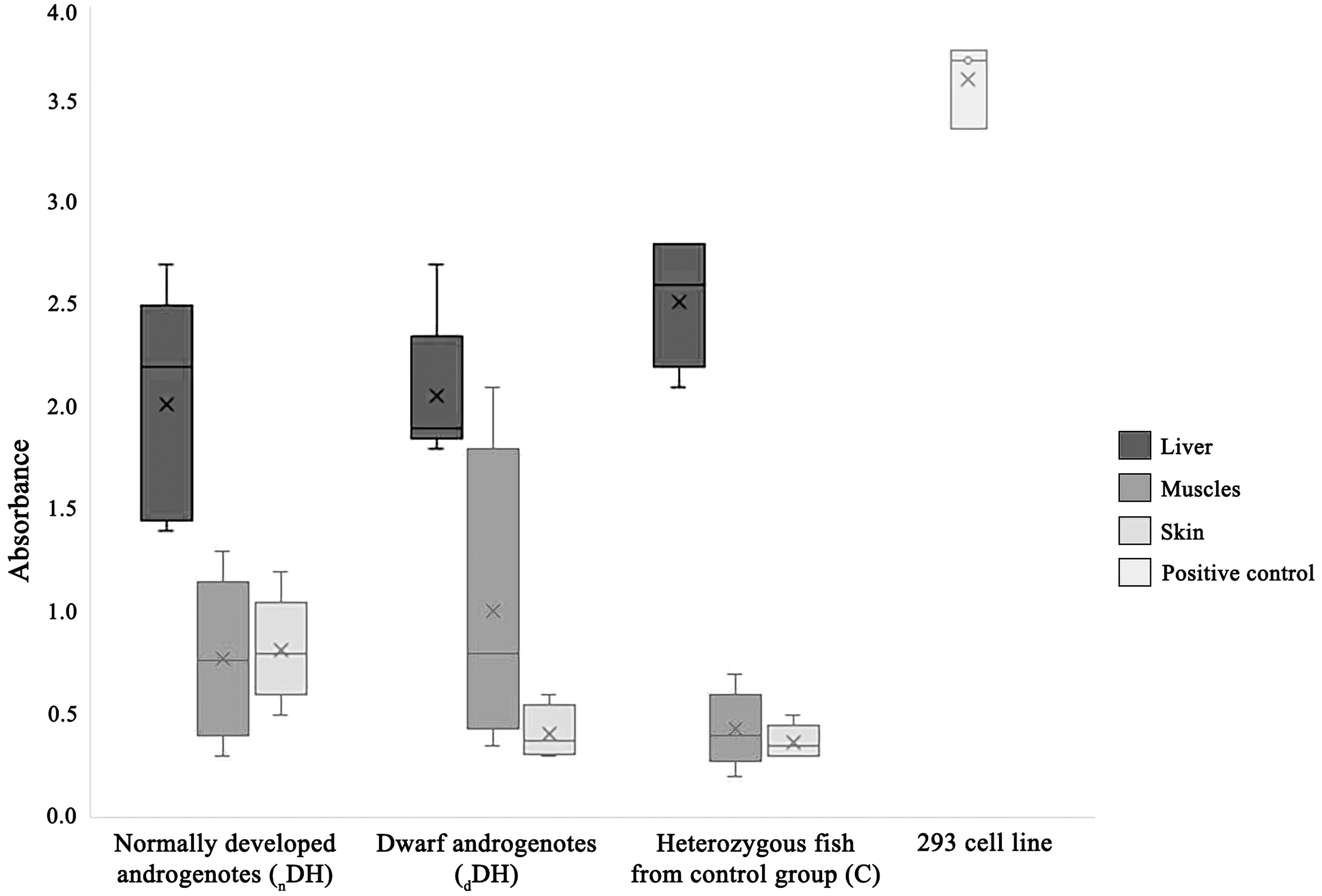
Telomerase activity (expressed in absorbance) in the somatic tissues from the examined rainbow trout. 293 cell line: Human, telomerase-positive embryonic kidney cell line (3.61 ± 0.27).
The activity of telomerase in the muscles of androgenetic trout from nDH and dDH groups was 0.78 ± 0.39 and 1.01 ± 0.76, respectively. The lowest activity of telomerase was observed in the muscles of fish from the control group (0.43 ± 0.19), however, the differences were not significant.
The activity of telomerase in the skin sampled from nDH, dDH, and heterozygous individuals from the control group was as follows: 0.82 ± 0.26, 0.41 ± 0.19, and 0.37 ± 0.08, respectively. The analysis showed significantly (p < 0.05) higher telomerase activity in skin from normally developed androgenetic trout when compared to dwarf specimens and heterozygous individuals from the control group. In the case of one dwarf rainbow trout, no telomerase activity was detected in the skin and in the muscles. The activity of telomerase in 293 cell line was sufficiently high (3.61 ± 0.27) which confirms good quality of performed analysis.
Discussion
In contrast to mammals, increased telomerase activity has been detected in the somatic tissues from varied fish species irrespective of their age.13,16,36 Experiments conducted on several model and non-model fish species show that upregulated telomerase may be related to maintenance of the telomere length, somatic growth, and tissue regeneration.15,17,21,37–40
Here, 1 year old homozygous DH androgenetic rainbow trout with growth retardation and their normally developed homozygous and heterozygous siblings were examined to find out any differences in the telomerase activity among these fishes. Irrespective of the genetic status and fish appearance, the pattern of telomerase activity was similar with the highest abundance found in the liver tissue and the lower activity reported in the muscles and skin. Comparable activity of telomerase in dwarf and normally developed trout are paralleled with results of our previous experiment showing no differences in the telomere length between rainbow trout with growth retardation and those showing typical growth rate. 25 Equal length of telomeres observed in the rainbow trout siblings with disturbed and normal growth is thus explained by comparable activity of telomerase in those fish that compensate for any loss of the telomeric DNA. 26
Although activity of telomerase has been confirmed in the muscles of all fishes studied to date under this regard,13,15,37–39,41 its role in the fish somatic growth is not unambiguous. Some fishes are thought to exhibit indeterminate growth with the fastest growth rate observed during the early stage of life. 42 In adults, the growth rate slows down but never stops. 43 Thus, it was hypothesized first that indeterminate growth in fish might be associated with ubiquitous telomerase activity.13–16,19 In marine medaka (Oryzias melastigma), early growth characterized by the rapid increase of the body mass is accompanied by upregulation of the telomerase activity in the muscles. 19 However, activity of telomerase changed upon maturity in these fishes. In sexually matured marine medaka, activity of telomerase was maintained in females but declined in males and was not related to the body mass. 19
In the European hake (Merluccius merluccius) upregulation of TERT expression observed in the muscles and skin has been paralleled with increasing of the body size. 16 In turn, reduction of the telomerase activity in muscles in adults has been reported in Atlantic cod (Gadus morhua) 16 and rainbow trout 13 but not in zebrafish (Danio rerio). 41 These results suggest that telomerase may play some role during early development and growth in fish; however, different patterns of telomerase activity observed in adults between species indicate indefinite function of telomerase upon maturity. 19 Insignificant differences in the telomerase activity in muscles from rainbow trout with growth deficiency and their normally developed siblings confirmed that telomerase in dwarf trout is not downregulated. On the other hand, significantly reduced abundance of telomerase observed in the skin of dwarf androgenotes may indicate that telomerase is somehow committed to processes related to growth in rainbow trout at least in the skin tissue, but further studies need to be performed to provide more information concerning this phenomenon.
In contrast to mammals, fish are considered as highly regenerative animals with capacity to regrow damaged heart, spinal cord, brain, and liver, among others.14,21,40,44 A direct correlation between regeneration efficiency and telomerase expression observed in various fish species suggests that such upregulation of telomerase is not linked to their longevity or indeterminate growth but to the regeneration ability of injured, damaged, or amputated tissues.14,17,36,41 Thus, the retained telomerase activity prevents telomere attrition during increased cell proliferating rates that accompany organ regeneration process. Liver is the organ that is famous for its capability to fully restore after acute injury for the mass and function. In several fish species, liver that was also confirmed to have a high regeneration capacity 45 is listed as the organ with relatively high activity of telomerase.13,39,46 Similar has been observed in the present research where activity of telomerase was the highest in liver tissue in all groups of the sampled rainbow trout.
Conclusion
Our study indicated that telomerase in rainbow trout with growth deficiency is not downregulated at least in the muscles and in the liver. In contrast, skin exhibited decreased activity of telomerase in the dwarf rainbow trout. Unequivocal results concerning activity of telomerase in the dwarf rainbow does not verify the role of this enzyme in processes related to somatic growth in fish. In turn, increased abundance of telomerase observed in the rainbow trout liver may be related to its high regeneration capacity.
Footnotes
Acknowledgments
The authors thank Rafał Różyński from the Department of Salmonid Research, Inland Fisheries Institute in Olsztyn, Rutki for his technical assistance during the experiment.
Authors' Contribution
K.O. and L.P. designed and coordinated the study. L.P., A.B., and M.K. carried out the laboratory works and data analysis. M.K. performed the graphics preparations. All authors were involved in writing and editing of the article.
Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This study was supported by the National Science Centre (NCN) Poland project number 2017/27/B/NZ9/00113.