
Abstract
Regenerative medicine is an emerging field of research aiming to understand the wound healing mechanisms and to develop new therapeutic strategies. Nanocarriers are used to improve drug bioavailability, solubility, and therapeutic abilities. In this study, we used for the first time curcumin loaded oligo kappa-carrageenan-graft-polycaprolactone (oligoKC-g-PCL) nanomicelles to investigate their regenerative potential using a model of tail amputation in zebrafish eleutheroembryo. First, we showed that curcumin encapsulated oligoKC-g-PCL spherical micelles had a mean size of 92 ± 32 nm and that micelles were successfully loaded with curcumin. These micelles showed a slow and controlled drug release over 72 h. The toxicity of curcumin nanomicelles was then tested on zebrafish eleutheroembryo based on the survival rate after 24 h. At nontoxic concentration, curcumin nanomicelles improved tail regeneration within 3 days postamputation, compared with empty micelles or curcumin alone. Furthermore, we demonstrated that curcumin nanomicelles increased the recruitment of neutrophils and macrophages 6 h postlesion. Finally, our study highlights the efficiency of oligoKC-g-PCL nanomicelles for encapsulation of hydrophobic molecules such as curcumin. Indeed, our study demonstrates that curcumin nanomicelles can modulate inflammatory reactions in vivo and promote regenerative processes. However, further investigations will be required to better understand the mechanisms sustaining regeneration and to develop new therapeutics.
Introduction
Wound healing is a natural process that involves restoration of the damaged cells, tissues, and organs. Depending on the cause of the wound, it can be acute or chronic. Acute wounds include traumatic lesions, while chronic wounds are associated with chronic diseases such as diabetes, vascular, or degenerative diseases. 1 A better understanding of the mechanisms of self-renewal and the cellular basis of wound pathology is needed for the development of new therapeutic strategies to effectively manage the wounds. 2 In that context, regenerative medicine is an emerging field of research which is important for the discovery of new therapeutics that promote tissue repair. 3 Many studies have investigated the healing potential of plants and have reported the benefits of some plant-derived bioactive molecules such as curcumin. Curcumin is a hydrophobic polyphenol molecule present in turmeric isolated from Curcuma longa that is widely used as a curry spice. It is the major curcuminoid of turmeric displaying many therapeutic properties, including anti-inflammatory, antioxidant, antiproliferative, antiangiogenic, antidiabetic, and wound healing.4,5 However, the hydrophobic nature, poor bioavailability, low solubility, as well as rapid metabolization, degradation, and clearance, of curcumin limit its therapeutic benefits. 6 To overcome these limitations and enhance the therapeutic benefits of curcumin, many nanoformulations, including liposomes, polymeric micelles, and polymer-curcumin conjugates, have been developed. 7
Polymeric micelles made of sulfated marine polysaccharides, including alginin, carrageenan, chitosan, and dextran, copolymerized with different polymers are newly emerging in wound healing research and therapeutics.8–10 Marine sulfated polysaccharides have many bioactivities such as antioxidative, antimicrobial, wound healing, anticancer, anti-inflammatory, and antiviral properties. 11 In a recent study, we encapsulated curcumin into carrageenan nanomicelles (Cur-micelles) and showed that it was noncytotoxic (in vitro in EA-hy926 endothelial cells and in vivo in adult zebrafish). 12 As well, the curcumin encapsulation improved the cellular uptake of curcumin and promoted the anti-inflammatory effect of curcumin. 12 In this work, we determined the healing potential of our Cur-micelles in vivo in zebrafish. The use of zebrafish allows to study various pathophysiological processes and has gained the attention of researchers in many fields.13–15 Indeed, zebrafish shares a strong genomic homology, as well as common physiological processes with mammals. 16 Due to their ability to regenerate, zebrafish has emerged as a model to study wound healing and regeneration.17–20
Many zebrafish models have been developed in adult fish and eleutheroembryo to study regenerative processes occurring in the liver, pancreas, spinal cord, heart, brain, retina, and tail.21–24 To this end, we decided to use a simple and fast model of tail amputation in zebrafish eleutheroembryo to study regeneration. 25 Indeed, the zebrafish tail is easily accessible for amputation and has a simple cellular organization. The tail regeneration process involves (1) epidermal cell migration that covers the wound and epidermal apical cap signaling center, (2) cell dedifferentiation into mesenchymal blastema structure, and (3) proliferation of blastema and replacement of amputated tissue. 26 Many signaling molecules, including sonic hedgehog ligands, 27 Wnts, 28 bone morphogenic proteins,29,30 fibroblast growth factors (FGFs), 31 insulin-like growth factors, 32 and retinoic acids, 33 are involved in the fin regeneration process. As most organs start to function within 5 days postfertilization, it is relatively reliable to use eleutheroembryo for large sample size screening and shorter time frames.
In the present study, curcumin-encapsulated oligoK-carrageenan nanomicelles were prepared and characterized by nuclear magnetic resonance (NMR) and dynamic light scattering (DLS) analyses and the drug release studied according to the previously published work. 12 The curcumin nanomicelle toxicity was tested in 3 days postfertilization (dpf) eleutheroembryo for 24 h by monitoring the survival rate. Next, a tail injury on 3-dpf eleutheroembryo was performed to investigate the effect of curcumin nanomicelles on tail regeneration. As inflammation and cell recruitment play a major role in reparation, the recruitment of neutrophils and macrophages at the damaged site was studied using the same model.
Materials and Methods
Preparation of carrageenan micelles
Carrageenan micelles were prepared as previously described.12,34
Carrageenans were extracted from red algae Kappaphycus alvarezii obtained from Ibis Algaculture (Madagascar). The carrageenans extracted were composed of κ-carrageenan (68%) and λ-carrageenan (32%) as analyzed previously by NMR.
The ƙ-carrageenase enzyme, which hydrolyzes the β-(1 → 4) linkage of carrageenan, was used to digest κ-carrageenan into oligocarrageenans. This enzyme was isolated from Pseudoalteromonas carrageenovora (DSMZ, Germany). The bacteria were stimulated with a 0.15% (M/v) ƙ-carrageenan solution.
Enzymatic digestion of ƙ-carrageenan was done using ƙ-carrageenase.
The carrageenan micelles were synthesized in three steps (1) partial acetylation of oligocarrageenan (digested carrageenan) hydroxyl groups, (2) grafting of hydrophobic caprolactone on partially acetylated oligocarrageenan, and (3) deprotection of the partially acetylated hydroxyl groups. 12
Drug loading
Curcumin encapsulation was done using the acetone volatilization method as previously described. 12
Five hundred milligrams of carrageenan micelles were mixed with 50 mg of curcumin and dissolved in 10 mL of acetone. This mixture was slowly agitated overnight at room temperature by dropwise addition of deionized water (250 mL). Acetone was evaporated at 30°C followed by elimination of the nonencapsulated curcumin by a 0.22 μm filter (stericup GP Millipore).
Characterization of carrageenan micelles
NMR analysis
NMR analysis was performed on a 600 MHz Avance III Bruker NMR spectrometer equipped with a 1 H/ 19 F, 13 C, and 15 N cryoprobe. 1D 1 H and 1D 13 C spectra were recorded in 100% D2O or CDCl3 (Eurisotop, France) at room temperature, and tetramethylsilane (Sigma-Aldrich, Germany) was used as a reference as described previously. 1 H NMR spectra were recorded with 128 scans using a sweep width of 10 ppm. 2D spectra were obtained with 32 scans and a sweep width of 10 ppm for 1 H and 120 ppm for 13 C. In all spectra, the carrier was placed at 4.7 ppm for 1 H and 50 ppm for 13 C as performed previously. 12
Determination of particle size
The particle size distribution of carrageenan micelles was analyzed using DLS particle size analyzer (Zetasizer Nano, Malvern Instruments) as previously described. One milliliter of an aqueous solution of amphiphilic micelles prepared in phosphate-buffered saline (PBS) was filtered through a 0.22 μm pore size polytetrafluoroethylene syringe filter to remove any aggregates. The critical micelle concentration (CMC) was determined at concentrations of 1 and 2 mg/mL. CMC was obtained by plotting the logarithm of the intensity of the scattered light as a function of oligocarrageenan-graft-polycaprolactone (PCL) concentration as previously described. 12
Drug release
A total of 20 mg of curcumin or 20 mg of curcumin-loaded micelles were dissolved in 10 mL of PBS and were subjected to dialysis using a Cellu-Sep H1 dialysis membrane (molecular weight cut-off = 2000 Da) immersed in 200 mL of PBS at 37°C. The amount of curcumin released in the PBS was measured by absorbance at 420 nm over 3 days.
Zebrafish husbandry
Zebrafish (Danio rerio) were housed in the facility at the CYROI/DéTROI (La Réunion). Adult fish (AB strain) were maintained under standard conditions of temperature (28.5°C for adult fish and 26.5°C for eleutheroembryo), photoperiod (14-h dark/10-h light cycle), pH (7.4), and conductivity (400 μS). 25 Fish were fed thrice a day with Gemma Micro 300 (Skretting). All the animal experiments were conducted in accordance with the French and European Community Guidelines for the Use of Animals in Research (86/609/EEC and 2010/63/EU).
Note that in this article, the term “eleutheroembryo” refers to a developmental stage from 0 to 120 h postfertilization (hpf). The term “larva” is used when the animal begins to feed autonomously, from ∼5 to 6-dpf.
Zebrafish eleutheroembryo acute toxicity test
To determine nontoxic doses that could be used, the toxicity of micelles, curcumin, and curcumin-encapsulated micelles was evaluated on 3-dpf eleutheroembryo. Newly fertilized eggs were maintained in fish water for normal development at 26.5°C. At 3-dpf, zebrafish eleutheroembryo (10 per group) were treated with different concentrations (5, 10, and 30 μM) of empty micelles, curcumin, and curcumin encapsulated micelles for 24 h. Based on our previous in vitro study showing an anti-inflammatory activity at 15 μM of curcumin-encapsulated micelles, concentrations of curcumin-encapsulated micelles ranging from 5 to 30 μM were tested on zebrafish eleutheroembryo. After 24 h of treatment, eleutheroembryo survival was estimated with a stereomicroscope (Nikon SMZ18) as the number of live animals without morphological malformations (such as pericardiac edema, spinal curvature, or lower developmental rate) divided by the total number of animals.
Tail amputation and treatment
Amputation and treatments were performed on the 3-dpf zebrafish eleutheroembryo as previously described. 25 Briefly, eleutheroembryo was first anesthetized with 0.02% tricaine (MS-222). They were then placed on a microscopic slide, and the caudal fin was amputated just close to the notochord using a blade, under a stereomicroscope. After the amputation, the eleutheroembryo was immediately placed back in E3 medium (Control) or E3 medium in presence of the respective treatments namely micelles, curcumin, and curcumin encapsulated micelles at 10 μM.
Analyses of immune cell recruitment and tail regeneration
The investigation of the recruitment of macrophages (lymphocyte cytosolic protein 1 [Lcp1]-positive cells) and neutrophils (myeloperoxidase [Mpo]-positive cells) was performed on 3-dpf eleutheroembryo exposed to the abovementioned treatments for 6 h after the tail amputation. Briefly, the amputated eleutheroembryos were placed in micelles, free curcumin, and curcumin encapsulated micelles at 10 μM for 6 h, before being fixed in 4% paraformaldehyde overnight at 4°C and processing for immunohistochemistry experiments.
For tail regeneration assay, the amputated eleutheroembryos were placed in the abovementioned treatments for 3 days with daily renewal of the treatments. Pictures were taken using a stereomicroscope under anesthesia, at 0 and 3-days postlesion. The regeneration area was selected and quantified using ImageJ software as previously described. 25
Tissue preparation and immunohistochemistry
For Lcp1 staining, fixed eleutheroembryo was washed four times with 1 × PBS containing 0.1% Tween 20 (PTW) for permeabilization. Then, eleutheroembryo was washed thrice in distilled water for 5 min and further permeabilized with cold acetone (−20°C) for 10 min at room temperature. Eleutheroembryo was next washed with distilled water and PTW. Then, blocking of nonspecific sites was performed using a blocking buffer (PTW, 2% bovine serum albumin and 1% dimethyl sulfoxide) for 1 h at room temperature. Eleutheroembryos were then treated with primary rabbit anti-Lcp1 antibody (GeneTex, reference: GTX124420, dilution: 1/200 in blocking buffer) and incubated overnight at 4°C.
For Mpo staining, fixed eleutheroembryos were dehydrated with 25%, 50%, 75%, and 100% methanol series and stored at −20°C overnight. Eleutheroembryos were next gradually rehydrated and washed four times in PTW for 5 min. After 1 h in blocking buffer, eleutheroembryos were treated with primary rabbit anti-Mpo antibody (Abcam; ref: ab210563, dilution: 1/200 in blocking buffer) and incubated overnight at 4°C.
The next day, after washes with PTW, eleutheroembryos were incubated with fluorescent secondary antibody (Alexa Fluor 594; Thermo Fisher; ref: A11005; dilution: 1/500 in PTW) and 4′,6-diamidino-2-phenylindole cell nuclei counterstaining (Sigma-Aldrich; ref: D9542; 300 ng/mL in PTW) for 2 h at room temperature. Finally, eleutheroembryos were washed thrice with PTW and visualized by fluorescence microscopy (Nikon eclipse 80i).
Statistical analysis
Data were expressed as mean ± standard deviation of three independent experiments. Statistical analysis was performed by one-way analysis of variance (ANOVA) followed by Bonferroni's multiple comparison test and two-way ANOVA for drug release studies using GraphPad Prism 6 (GraphPad Software, Inc., San Diego, CA). Statistical significance was considered for a p-value ˂0.05.
Results
Characterization of carrageenan micelles by NMR analysis
Oligo kappa-carrageenan-grafted-PCL (oligoKC-g-PCL) was synthesized using a previously described method. 12 Briefly, hydroxyl groups of digested carrageenan (oligocarrageenan) (Fig. 1a) were partially acetylated and copolymerized with hydrophobic Ɛ-caprolactone (PCL) on the other remaining hydroxyl groups. Self-assembled amphiphilic carrageenan micelles in an aqueous solution were obtained by removing acetyl groups.

Characterization of carrageenan nanomicelles by NMR analysis.
The 1 H NMR analysis of digested and undigested carrageenan is represented in Figure 1a. Partially acetylated oligoKC-g-PCL was analyzed using 1 H NMR. The peak at 2.13 ppm corresponds to acetyl functional groups (Fig. 1b). The 1 H NMR spectra of partially acetylated oligoKC-g-PCL indicated 60% of acetylation based on integration values of protons at 5.04 and 4.6 ppm (corresponding to anomeric H in oligocarrageenan) and at 2.13 ppm (acetyl groups).
Oligocarrageenan-graft-caprolactone analyzed by 1 H NMR spectrum (Fig. 1c) showed both PCL signals between 1.3 and 4 ppm; oligocarrageenan signals from 3 to 5.5 ppm. The acetyl protons which were detected in the above step at 2.09 ppm were not detected in this spectrum (Fig. 1c) confirming the removal of partial acetyl groups in the last step.
The results of carrageenan micelle (oligoKC-g-PCL) characterization were in agreement with the previous results.
Drug loading and release studies
Hydrophobic curcumin was encapsulated into carrageenan micelles using acetone volatilization method, and drug release studies were done using dialysis method. 12 The amount of drug encapsulated was determined by UV absorption and calibration curve. The drug release studies of curcumin encapsulated nanomicelles (Cur-micelles) were done in 1 × PBS and dialysis method at 37°C (Fig. 2). Curcumin was released over a period of 4 days with 60% released after 72 h.

Kinetic studies of drug release 20 mg of curcumin or curcumin loaded in micelles were dialyzed using a 2000 Da dialysis membrane as described in the methods. Drug released into the PBS medium was measured by absorbance at 420 nm. Data are represented as mean ± SD of three independent experiments n = 3. ***p ˂ 0.001 and **p ˂ 0.01 as curcumin encapsulated in micelle compared to curcumin alone. PBS, phosphate-buffered saline; SD, standard deviation.
The drug release rate and amount of curcumin encapsulated in nanomicelles were similar to the previously developed micelles 12 and indicated a controlled drug release. As demonstrated previously, the drug release rate can vary depending on the interaction between the drug and micelle core.
Particle size determination of carrageenan micelles (oligoKC-g-PCL) and curcumin encapsulated micelles by DLS analysis
Using DLS analysis, the average particle size of empty micelles was determined to be 142 ± 93 nm and curcumin-encapsulated micelles (Cur-micelles) was 92 ± 32 nm as shown in Figure 3a and b, respectively. The size of these micelles and curcumin encapsulated micelles was slightly smaller than previously described, with 142 versus 187 nm for the micelles and 92 versus 177 nm for the loaded micelles.

Particle size distribution of carrageenan nanomicelles determined by DLS analysis. Samples were prepared in PBS at 1 μg/mL. Size distribution of empty micelles
The CMC estimated by DLS was 4.10–5 M. A polydispersity index (PDI) near to zero implies homogeneity of dispersions, and a PDI >0.3 indicates heterogeneity. The PDI of our Cur-micelles nanoparticles was around 0.2. The particle size of curcumin encapsulated micelles is smaller than the empty micelles due to the strong interaction of curcumin with the hydrophobic PCL core of micelles which may cause the shrinkage of the micelles.
Acute toxicity of curcumin nanomicelles in 3-dpf zebrafish eleutheroembryo
To study the toxicity of micelles, free curcumin, and curcumin encapsulated micelles, 3-dpf zebrafish eleutheroembryos were treated with different concentrations of the abovementioned conditions for 24 h. Micelles alone did not show a toxic effect on the survival rate at 5, 10, and 30 μM (Fig. 4). In contrast, a significant mortality was observed for curcumin alone and curcumin-encapsulated micelles at 30 μM. At lower concentrations (5 and 10 μM), we did not observe any death. Based on these results, we decided to use the concentration of 10 μM of empty micelles and curcumin-encapsulated micelles for assessing their biological properties.

Toxicity of curcumin nanomicelles on 3-dpf zebrafish eleutheroembryo. The toxicity was determined by measuring the survival rate of eleutheroembryo after 24 h of exposure with E3 medium (control), empty micelles, curcumin, and Cur-micelles (5, 10, and 30 μM). Survival rate percentage was calculated by counting the number of alive and dead eleutheroembryo after the different treatments for 24 h. Curcumin and curcumin nanomicelles were both toxic at 30 μM concentration. Data are represented as mean ± SD of three independent experiments with 20 eleutheroembryos per condition. ****p ˂ 0.0001 compared to control (one-way ANOVA). ANOVA, analysis of variance; dpf, days postfertilization.
Effect of curcumin nanomicelles on regeneration and immune cell recruitment in a model of tail injury
The effect of curcumin encapsulated micelles on regeneration was first tested. Based on the above results, free curcumin and empty and curcumin-loaded micelles were investigated at a 10 μM concentration for their potential effects on regeneration in a model of tail amputation. At 3-dpf, eleutheroembryos were amputated and treated with E3 medium, micelles, curcumin, and curcumin-encapsulated micelles. Three days after the transection, the tail regeneration was significantly increased with curcumin-encapsulated micelles compared to control, micelles, or curcumin alone (Fig. 5), demonstrating pro-regenerative properties of the encapsulated curcumin in this model.

Curcumin nanomicelles promote the regeneration of the zebrafish tail after amputation.
To analyze if this improved regeneration involved a change in immune cell recruitment, the effect of curcumin-encapsulated micelles was tested on the recruitment of neutrophils and macrophages through Mpo and Lcp1 immunostainings, respectively.
As shown in Figure 6, the number of macrophages (Lcp1-positive cells) was significantly higher in eleutheroembryo treated with curcumin-encapsulated micelles compared to control, empty micelles, or curcumin alone. Similar results were obtained for neutrophils (Mpo-positive cells) (Fig. 7). Indeed, a higher number of neutrophils were observed at the site of injury in the eleutheroembryo treated with curcumin-encapsulated micelles compared to control, micelles, or curcumin alone.
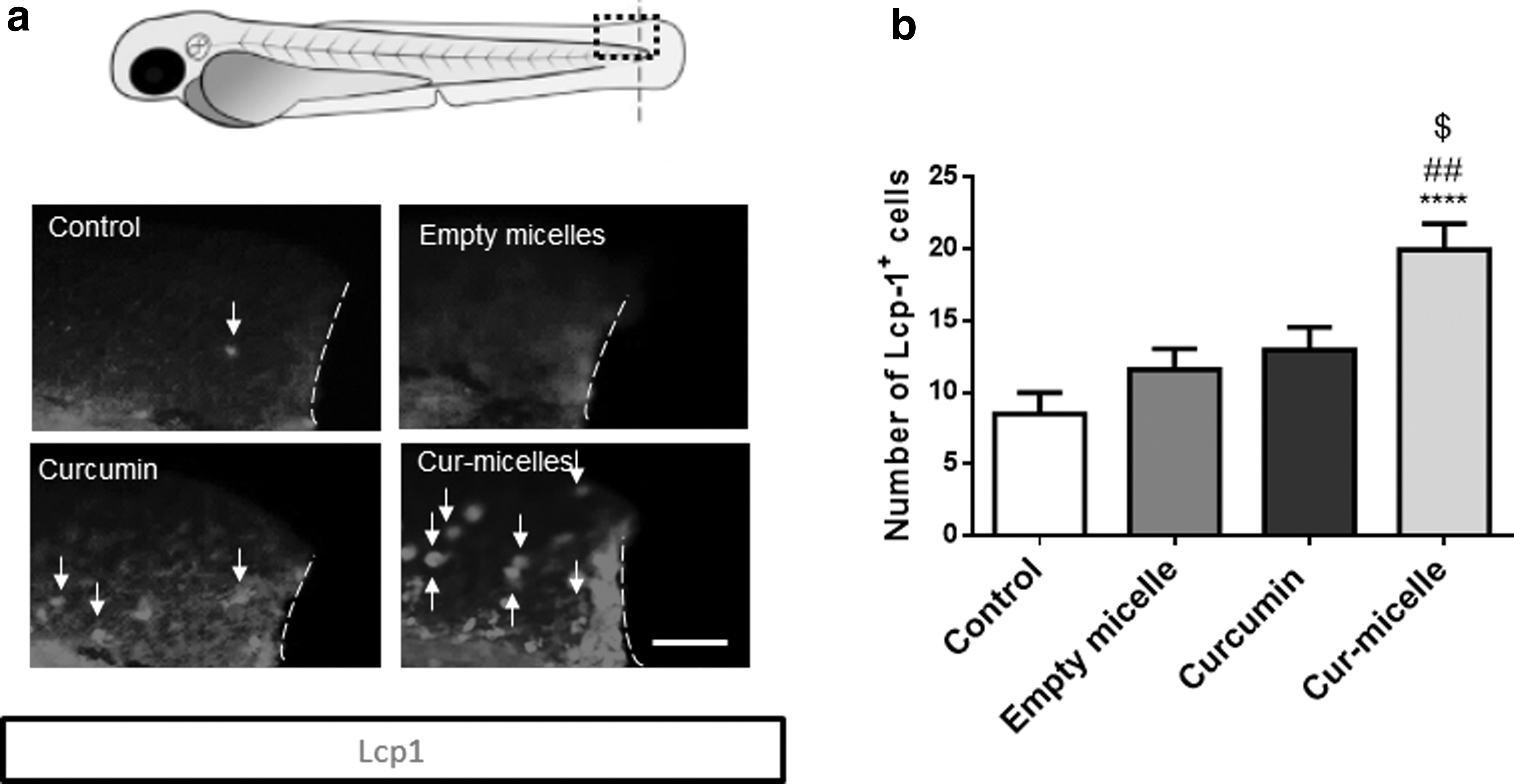
Curcumin nanomicelles increase the recruitment of macrophages at the tail injury site.

Curcumin nanomicelles increase the recruitment of neutrophils at the tail injury site.
Discussion
Studies on polyphenolic molecules, including flavonoids, reveal that they promote wound healing by several mechanisms such as the scavenging of free radicals and reactive oxygen species and suppression of acute inflammation through time-dependent regulation of pro-inflammatory and anti-inflammatory chemokine expression.35,36 Among the polyphenols, curcumin has been studied for its regenerative wound healing properties.37,38 The antimicrobial property of polyphenolic molecules, including curcumin, also contributes to wound healing by avoiding prolonged inflammation through the reduction of the microbial load at the site of injury. 39 However, the therapeutic benefits of curcumin are limited by its poor bioavailability and solubility due to its hydrophobic nature. 40
Many nano-formulations of curcumin-encapsulated nanoparticles (Cur-micelles) have been developed recently to overcome these limitations and enhance its therapeutic properties. 41 The biodistribution of Cur-micelles in zebrafish and mice showed their accumulation in liver, lungs, and brain tissues. 12 In the present work, the characterization of our curcumin-encapsulated nanomicelles showed similar parameters to those found in our previous study, 12 with a mean particle size of 142 ± 93 nm and a drug release of 60% by 72 h. This demonstrates a reproducible and stable production of oligoKC-g-PCL nanomicelles and curcumin-encapsulated nanomicelles.
Although many beneficial effects of curcumin have been shown in wound healing and regeneration in in vitro and in vivo models (mice, rats, and rabbit), 1 the effect of nano-formulated curcumin on zebrafish tail regeneration has not yet been studied or explored. We then investigated the regenerative properties of these Cur-micelles in vivo using zebrafish eleutheroembryos. Our study showed that the Cur-micelles enhanced tail regeneration compared to curcumin or micelle alone. Zebrafish fin amputation causes acute inflammation, including the recruitment of immune cells, which triggers a rapid increase in reactive oxygen species (ROS) production. 42 Within 24 h of tail amputation, ROS-mediated chemokine and cytokine signals from the site of amputation tend to increase the proliferation and migration of epithelial cells to the site of injury and contribute to the formation of new tissue.
Neutrophils, with limited phagocytic capacity, are the first phagocytic cells present at the site of injury. Macrophages reach the site of injury later and play a crucial role in phagocytosis of the tissue debris and resolve the inflammatory responses and remain until the tissue repair. 43 Macrophages also promote fin regeneration by downregulating ROS levels at the site of injury. 42 Indeed, macrophage depletion was linked to decreased tissue regeneration, 43 while neutrophil depletion did not affect the fin regeneration in adult zebrafish. 44
This is in agreement with our results showing that the increased regeneration observed with Cur-micelles treatment is associated with an increased recruitment of neutrophils (Mpo-positive cells) and macrophages (Lcp1-positive cells) at the site of tail injury, as shown previously for other extract rich in polyphenols in the same tail injury model. 25
Caudal fin regeneration also involves cell migration and proliferation of epidermis and mesenchymal cells, formation of new blood vessels, and remodeling of the extracellular matrix. Wound epidermis at the tail injury covers the underlying matured tissue from which highly proliferative mesenchymal cells grow and regenerate the damaged tissue. 14 Many key signaling pathways are essential for wound repair. These include, among others, the Wnt, FGF, Notch, and Hedgehog pathways. 33 Indeed, FGF receptor fgfr 1 is involved in the signaling of epithelial cap, mesenchymal cells, and blastema recruitment needed for proliferation and regeneration of tail tissue.45,46 Sonic hedgehog (shh) signaling may also be involved in fin regeneration. 47 It could be interesting in future studies to investigate the effects of the Cur-micelles on these cells and molecular pathways.
Footnotes
Authors' Contributions
N.D., L.G., and C.L.D.H. designed the experiments. All the authors participated in the analysis of the experiments and/or in the writing of the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from La Reunion University and from FEDER (RE0022527-ZEBRATOX) EU-Region Reunion-French state national counterpart.