
Abstract
Parkinson's disease (PD) is a neurodegenerative disorder that is clinically assessed by motor symptoms associated with the loss of midbrain dopaminergic neurons affecting the quality of life for over 8.5 million people worldwide. The neurotoxin 6-hydroxydopamine (6-OHDA) has been used to chemically induce a PD-like state in zebrafish larvae by several laboratories; however, highly variable concentration, methodology, and reagents have resulted in conflicting results suggesting a need to investigate these issues of reproducibility. We propose a protocol that addresses the differences in methodology and induces changes in 6 days postfertilization (dpf) larvae utilizing a 24-h exposure at 3 dpf with 30 μM 6-OHDA. Despite ∼50% lethality, no morphological or development differences in surviving fish are observed. Definition of our model is defined by downregulation of the expression of th1 by reverse transcriptase-quantitative polymerase chain reaction, a marker for dopaminergic neurons and a reduction in movement. Additionally, we observed a downregulation of pink1 and an upregulation of sod1 and sod2, indicators of mitochondrial dysfunction and response to reactive oxygen species, respectively.
Introduction
Parkinson's disease (PD) is a disorder that affects ∼8.5 million people globally. 1 Clinically, it is diagnosed using characteristic motor symptoms that include resting tremors, rigidity, akinesia, and postural instability. 1 Diagnosis of PD is confirmed by threshold loss of A-9 midbrain dopaminergic (mDA) neurons. 2 Several approaches to utilize zebrafish as a tool for PD have been investigated to help characterize the genetic basis of disease as well as look for treatment options.3–5 The primary methodology of assessment has focused on reduction of molecular markers such as TH (Tyrosine Hydroxylase), SLC6A3 (Solute Carrier Family 6 Member 3), and SLC18A2 (Solute Carrier Family 18 Member A2) to characterize neuron loss and movement deficits.6–8 TH is the rate-limiting enzyme involved in the production of dopamine (DA). 9 SLC6A3 is a sodium-dependent DA transporter important for DA uptake from the synaptic cleft, while the SLC18A2 is critical for packaging DA into synaptic vesicles.
The goal of this article is to compare 6-hydroxydopamine (6-OHDA) protocols (Table 1) and provide an exposure method that allows us to observe behavioral changes in the larvae without larval toxicity or utilizing morphologically altered larvae. We hypothesized that 6-OHDA exposure would alter characteristic reactive oxygen species (ROS), mitochondrial dysfunction, and dopaminergic markers at the transcriptional level.
Available 6-Hydroxydopamine Protocols Differ in Reagents Composition, Endpoints, and Effectiveness
6-OHDA, 6-hydroxydopamine; dpf, days postfertilization.
Materials and Methods
Zebrafish care and maintenance
AB [ZIRC] zebrafish larvae came from in-house breeding or were used directly upon arrival from ZIRC. Fish care and maintenance are performed as described by Westerfield and coworkers. 10 All methodologies were approved by the UT Tyler IACUC (protocol 705).
Zebrafish embryos were collected from randomized bulk mating and placed into 100 × 15 mm petri dishes with 80 embryos per plate in embryo water (60 μg/mL Instant Ocean and 24 ppm of Methylene blue) and kept in a glass-front 28°C incubator. Larvae born on different dates were considered as different cohorts. In some cases, large cohorts of larvae exposed to independently measured 6-OHDA were considered different groups of fish. Larvae were dechorionated at 2 days postfertilization (dpf) and transferred to Netwell plates (Corning six-well plate with Netwell insert, 74 μm polyester mesh 24 mm insert). All solutions were changed daily during the experiment.
6-OHDA assessment
All work was performed with 6-OHDA-Hydrobromide (6-OHDA, Tocris) in the fume hood or with N95 respirators. 6-OHDA stocks were made fresh daily with L-ascorbic acid for stabilization. Experiments to assess how the variation of individual components affected larval were performed by altering the concentration of reagents either individually or in combination: 6-OHDA (1–750 μM), L-ascorbic acid (0.001%–0.2%), methylene blue (absence or presence), HEPES buffer (absence or presence), duration of exposure (24 h exposure or 48 h exposure), and age at exposure (2, 3, and 4 dpf).
6-OHDA preparation for molecular and behavioral experiments
Solid 6-OHDA was kept at −20°C until absolutely needed. To maintain consistency, 6-OHDA 100 μM stocks were prepared by adding 2.5 mg of 6-OHDA directly into a conical tube, and diluting it with 10 mL of the vehicle solution (0.2% L-ascorbic acid in embryo water buffered to pH 7.0 utilizing 40 mM HEPES [12 mL of 1 M HEPES free acid, and 8 ml of 1 M HEPES sodium salt in 480 ml embryo water]). This solution was further diluted 1:10 in vehicle solution by rinsing of the conical tube into a larger container. Additional dilutions were performed in vehicle to maintain concentrations of L-ascorbic acid and HEPES buffer.
6-OHDA exposure for molecular and behavioral experiments
At 2 dpf, larvae were moved to the Netwell plates—12 larvae in 12 mL embryo water per well. The larvae were exposed to 6-OHDA starting at 3 dpf for 24 h, and transferred daily to fresh vehicle solution until analyzed at 6 dpf.
Behavioral assessment
Six dpf larvae were placed in a 24-well plate—one fish per well. The plate was placed on a LED light plate (Biorad) and acclimated for 30 min. Behavior was recorded for 10 min (Phantom Camera, 30 frames per second, sensitivity 100). Videos were manually scored by three blinded individuals using a binary score of one (moved) or zero (no movement). Movement was defined as a turn and/or the larvae traveled a quarter of the way across the well. Percentages were calculated for each plate, and statistical analyses (T-test) performed using Prism V10.0.2 (Graphpad). Further analysis, of the fish that displayed movement, was performed using Fiji (ImageJ). Videos were opened as virtual stacks. Frames displaying movement of the larvae were manually notated. The virtual stacks were further modified by using the Z-project tool with minimum intensity projection and pathlengths measured.
RNA harvesting
RNA was harvested from exposures that contained greater than 10 morphologically normal larvae. Larvae were placed in TRIzol (Thermo Fisher Scientific), homogenized utilizing a single-use polymer pestle, and RNA extracted using an RNA extraction Kit (Zymo Research) using the manufacturer's protocol including the optional DNAase I incubation. RNA concentration and quality were measured using the Nanodrop One (Thermo Fisher Scientific).
Primer design and verification
Primers were designed using NCBI Primer BLAST 11 to meet MIQE guidelines. 12 cDNA was generated with the Sensifast cDNA synthesis kit (Meridian) following manufacture's protocol. Four hundred nanograms of RNA is utilized for each 20 μL reaction; however, the resultant cDNA was diluted 50% to increase subsequent reaction numbers. PCR was performed with the MyTaq Red PCR kit (Meridian) following manufacturer's recommendations. Primers that produced a single amplicon within 10% error of the theoretical size were purified utilizing DNA clean up kit (New England Biolabs) and Sanger sequenced (Eurofins Genomics). Sequence quality was assessed using A Plasmid Editor 13 and NCBI BLAST [Danio rerio (taxid:7955) genome].
For qPCR, cDNA was amplified using the PowerTrack SYBR green with ROX reagent on a Quantistudio5 (Thermo Fisher Scientific). Primer pairs were assessed on up to 5 samples and ran in technical triplicate with a nontemplate control on the same plate. Primer efficiencies were acquired with a serialized 50% dilution (50%, 25%, 12.5%, 6.25%) with efficiencies between 90% and 110% deemed acceptable (Table 2). Melt curves and agarose gel electrophoresis were used to verify reverse transcriptase-quantitative polymerase chain reaction (RT-qPCR) products. Outlier reactions from the technical triplicates were removed based on discrepancies within the triplicate (deviating more than a quarter Cq within the replicate). Analysis of amplification curves used the ΔΔ CT method using hprt1 as the endogenous control; T-tests were used to compare expression levels of exposed versus control for each primer pair (Quauntistudio, Thermo Fisher Scientific; Excel, Microsoft).
Primer Sequence for Endogenous Control and Genes of Interest
Results
We started by replicating procedures and concentrations for 6-OHDA from the available literature (1–750 μM 6-OHDA). In our laboratory, we could not replicate existing protocols that used higher 6-OHDA concentrations (greater than 50 μM) as lethality or morphological changes precluded our analysis (data not shown).3,4,14 Previous works in rats established the use of 0.2% L-ascorbic acid for 6-OHDA solubility in aqueous solutions. In more recent zebrafish protocols, L-ascorbic acid has been used; however, concentrations varied with some exposures using less than the established 0.2% concentration. 15
We performed a dosage curve to determine the effect of L-ascorbic acid alone on larval phenotypes. We determined that 0.001% L-ascorbic acid in embryo water can be utilized without observed lethality. Lethality is correlated with lowering the pH of the embryo water (data not shown), and the appearance of the larvae was similar to that observed when using other drugs that lower the pH of the embryo water. This suggested pH as the causative agent in lethality. A 6-OHDA dosage curve (0 − 250 μM) was performed with 0.001% L-ascorbic acid in embryo water as a vehicle. Higher concentrations demonstrated morphological features consistent with an acidic environment including tail curvature, visible cell death, and blebbing primarily from the yolk sac. Morphological defects and developmental delays were observed at lower concentrations between 50 and 70 μM depending on the cohort. We observed tail curvature, delayed or incomplete air bladder inflation, cardiac edema, irregular eye shape, and smaller eyes (data not shown). Furthermore, behavioral assessments displayed a high variability.
This is consistent with Kalyn and coworkers that concluded 6-OHDA is too toxic without buffering conditions. Vaz and coworkers utilized a HEPES pH buffer, and we experimentally determined that a vehicle of 40 mM HEPES buffer with 0.2% L-ascorbic acid could be utilized to make a vehicle solution at pH 7.0. In addition, we assessed this vehicle in the presence or absence of methylene blue. Methylene blue, a standard regent in our embryo water, is an antioxidant, and as expected, we saw a reduction of toxicity when utilizing methylene blue in our vehicle; therefore, it was maintained in the rest of our solutions.
Morphology and mortality
We performed dosage curves utilizing 6-OHDA in a vehicle consisting of 0.2% L-ascorbic acid in 40 mM HEPES buffered embryo water. We varied the 6-OHDA starting time (2, 3 , or 4 dpf) and duration of exposure (24 or 48 h) and assessed viability as an initial screening tool (Fig. 1). The results of our viability assessments are inconsistent with Feng and coworkers in that exposures starting at 2 dpf were too toxic. We saw lethality dipping below 50% at 10 μM of 6-OHDA (Fig. 1 and Table 1). This could be due to the increase of 6-OHDA availability afforded by the L-ascorbic acid. Consistent with Vaz and coworkers, treatment beginning at 4 dpf was less toxic, and we observed lethality above 50% up to 60 μM of 6-OHDA; however, this is observed at a concentration tenfold less than is reported in Vaz and coworkers (Fig. 1 and Table 1). We observed complete lethality at 750 μM (data not shown), that is, the dosage used in their work.
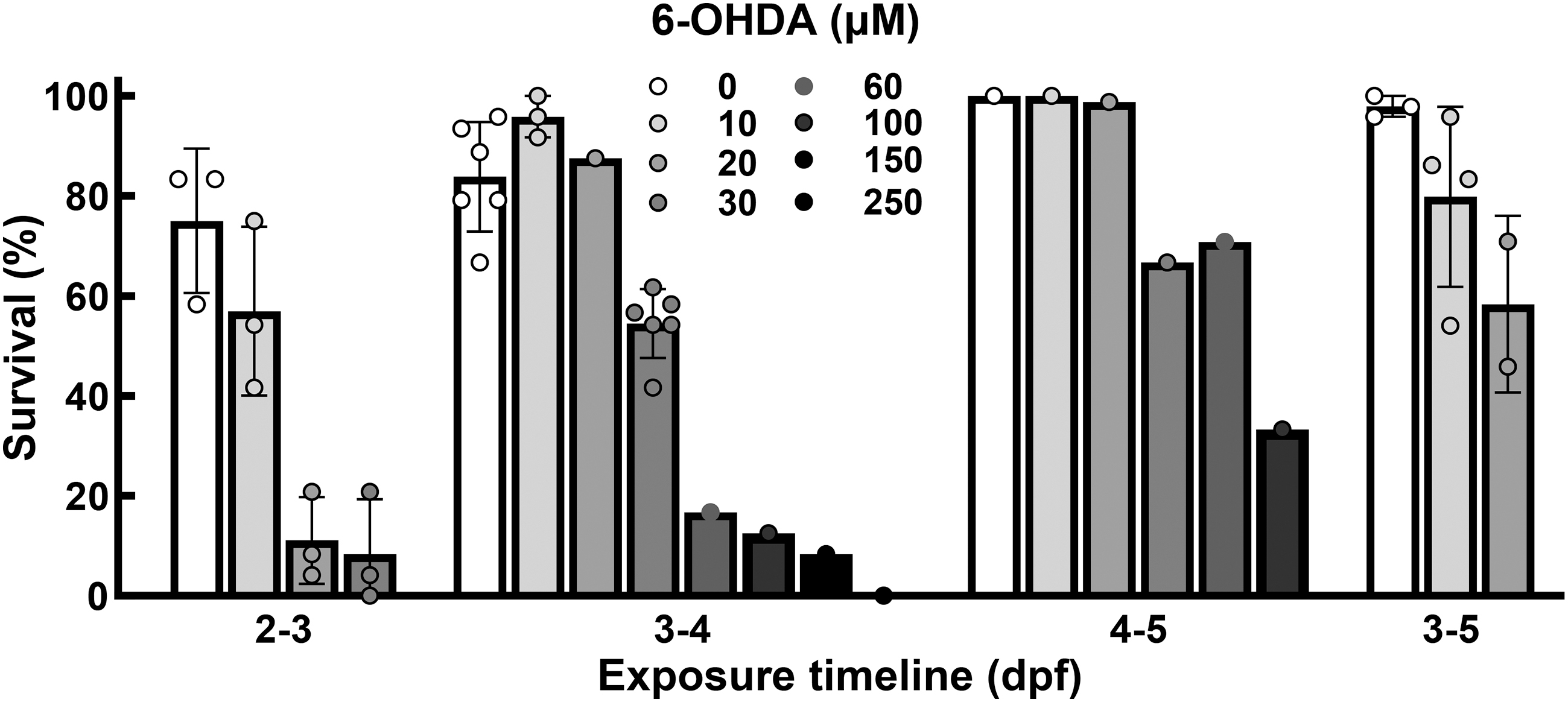
Viability varies based on duration and concentration of 6-OHDA exposure. Assessment of viability for four different protocols. The numbers below each group refer to the exposure age. Each individual data point refers to a new group of fish (Strain AB, n ≥ 24 per point). 6-OHDA, 6-hydroxydopamine.
Exposures lasting 24 h starting at 3 dpf seemed to have the best profile based on practicality, viability, and subsequent analyses. Concentrations above 30 μM 6-OHDA have a lethality greater than 50% in 24-h exposed fish starting at 3 dpf (3–4 dpf) (Fig. 1); therefore, we focused on the 30 μM 6-OHDA for subsequent analyses. Thirty micrometers 6-OHDA caused an average of 54.05% ± 6.41% (n = 504, Fig. 1). No morphological or developmental differences were observed for surviving fish in either exposed or control fish.
RT-qPCR quantification
RT-qPCR served as a second line of screening for exposed larvae. We reasoned that only those larvae that showed quantitative reductions of th1, a primary marker for dopaminergic cells, required further characterization. For our 3 dpf, 24 h 6-OHDA exposed larvae, we saw significant reductions in th1 RNA expression for 30 μM 6-OHDA but not 20 μM 6-OHDA. To confirm the significant reduction, we exposed three separate fish cohorts to 30 μM 6-OHDA or vehicle control at 3 dpf for a 24-h exposure. We also performed a within-cohort independent replicate allowing for four groups of pooled RNA for RT-qPCR analysis. The RNA was collected at 6 dpf—48 h after 6-OHDA exposure. A significant reduction of th1 and pink1 expression (p-value = 0.02 and p = 0.04, respectively, Fig. 2A) and an upregulation of sod1 and sod2 (p = 0.03 and p = 0.02, respectively, Fig. 2B) was observed. We did not observe significant differences in mDA neuron markers slc6a3 and slc18a2 (p = 0.63 and p = 0.13, respectively, Fig. 2C). Additionally, we looked at gad1b, a marker for GABAergic neurons, to assess specificity. We did not observe a significant difference in expression (p > 0.99, Fig. 2C).
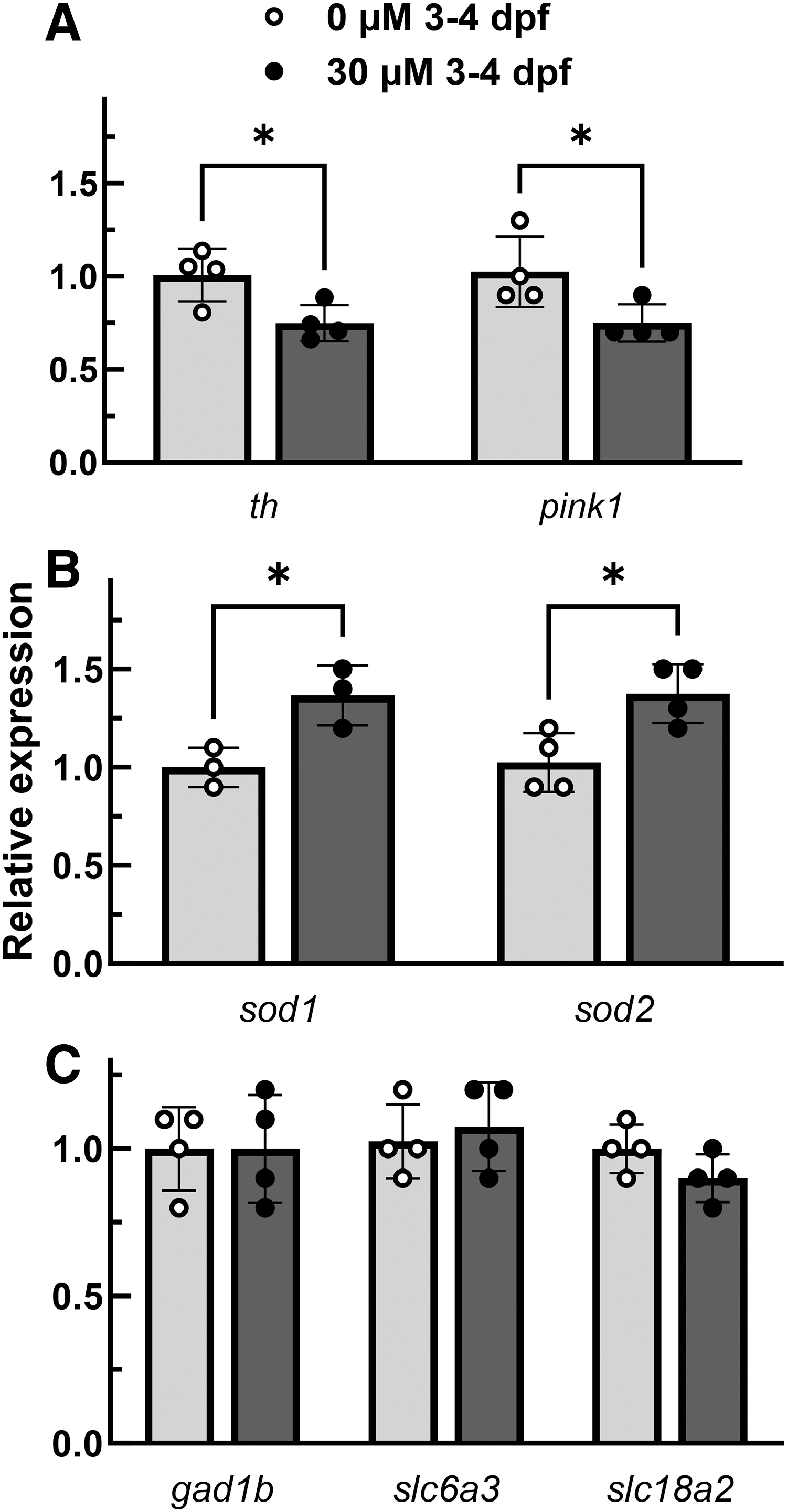
6-OHDA selectively reduces th1 expression and induces a state of mitochondrial dysfunction through ROS introduction. RT-qPCR of four groups exposed to 30 μM 6-OHDA for 24 h starting at day 3 (Strain AB, 24 embryos pooled per point). A significant reduction compared to controls occurred for th1, a marker for dopaminergic cells, and pink1, a marker for mitochondrial dysfunction
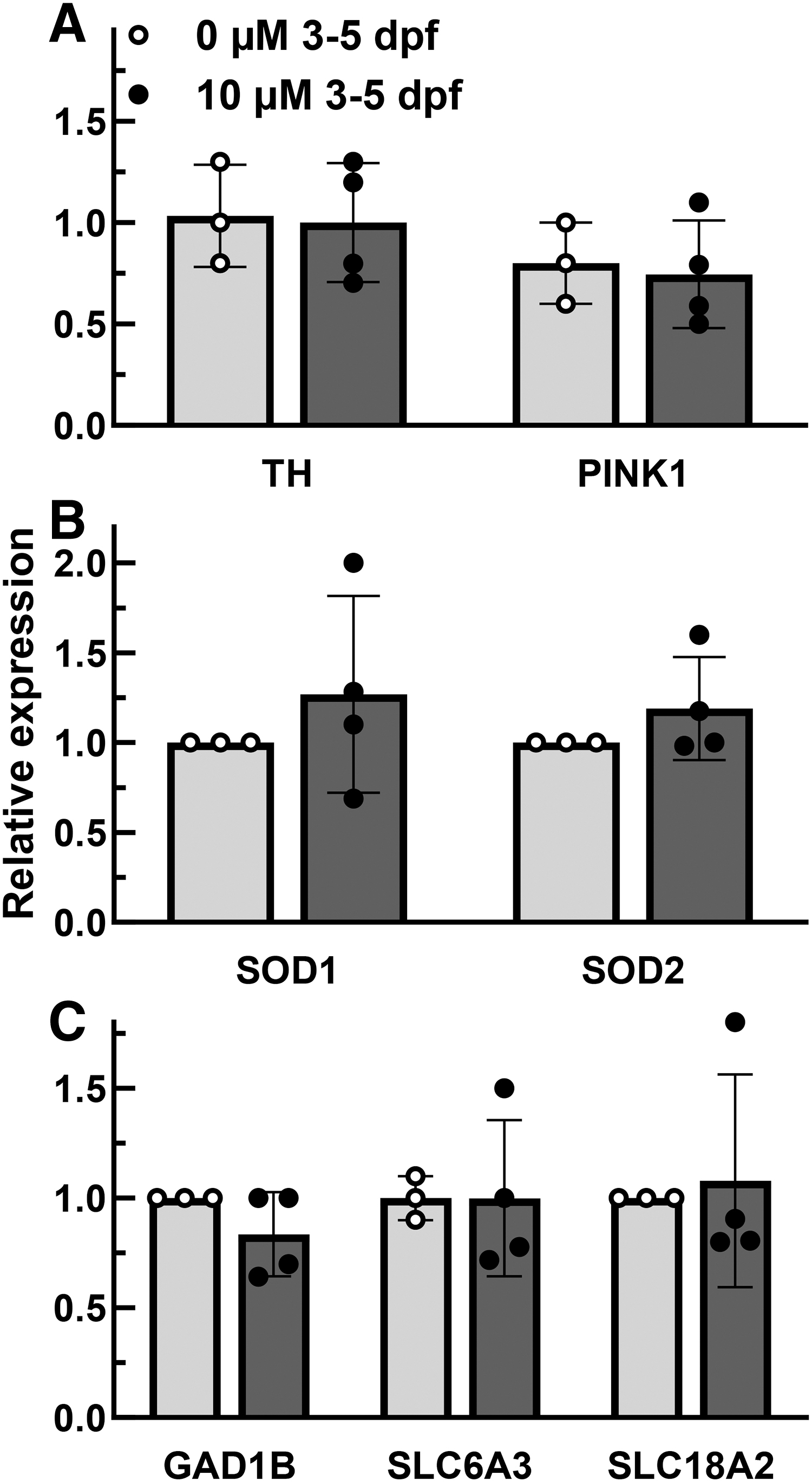
As part of our screening approach, we assessed the other conditions that had viability equivalent or better than the 3 dpf, 24 h 30 μM 6-OHDA exposure. Exposures starting at 4 dpf had a better viability (Fig. 1), so we assessed th1 expression in these cohorts. No difference in th1 expression was observed up to 60 μM 6-OHDA (the highest dose with greater than 50% viability (difference defined by not greater than 0.2-fold for a single event, 12 Fig. 3). The 3 dpf, 48-h 10 μM 6-OHDA exposure viability was improved, but we did not see a difference of th1 or any of the genes of interest (th1, p = 0.88; pink1, p = 0.78; sod1, p = 0.45; sod1, p = 0.32; gad1b, p = 0.21; slc6a3, p = 0.99; slc18a2, p = 0.79; Fig. 4).
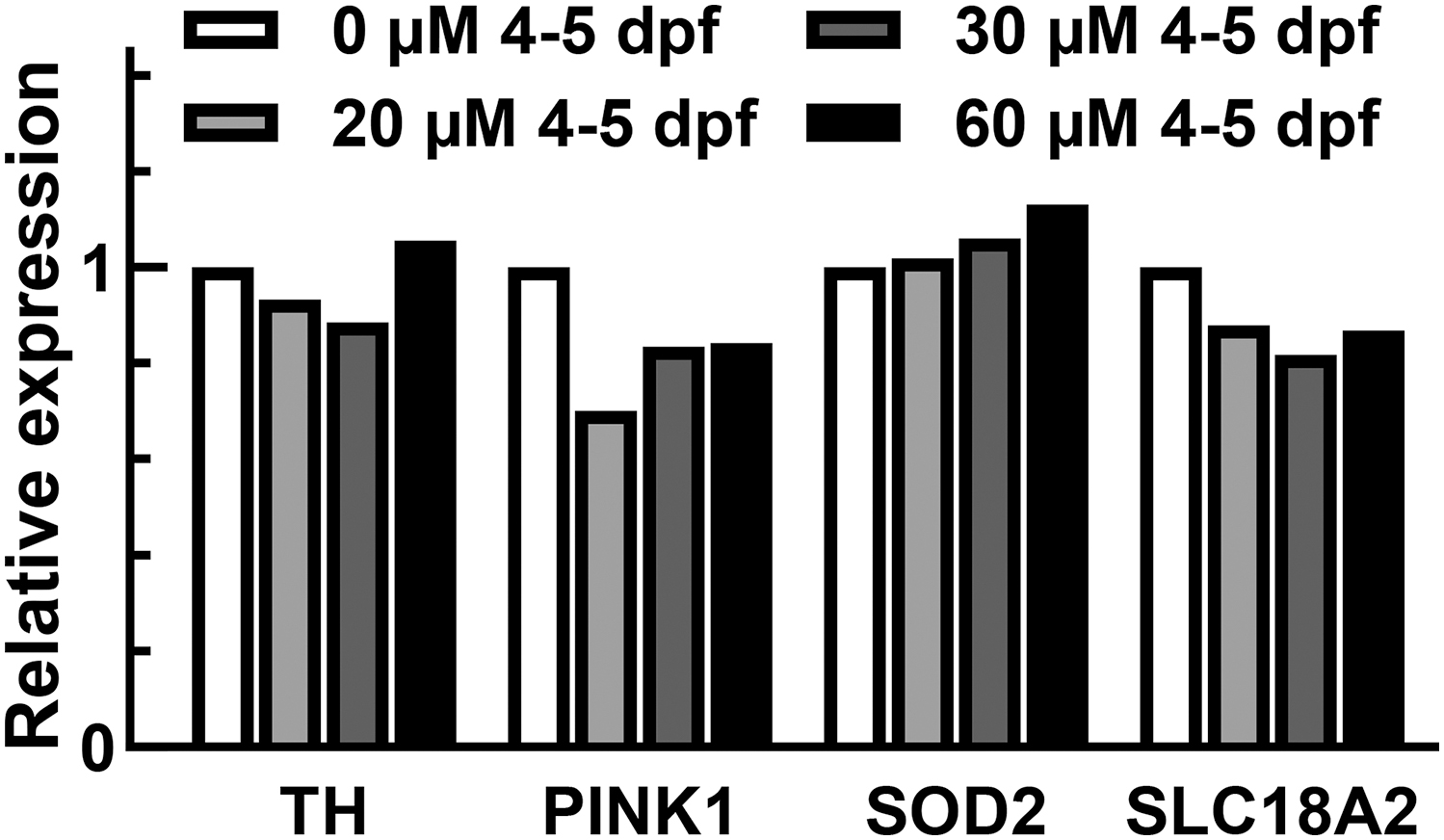
Fish exposed to 6-OHDA starting at 4 dpf do not show a reduction in th1. Vaz and coworkers had utilized a protocol with 24 h of treatment starting at 4 dpf. To assess if we could see expression changes using a similar timepoint, a RT-qPCR screen was performed in a single biological group. At doses that met viability and morphological criteria, 6-OHDA exposure did not induce a change greater than 0.2-fold (Strain AB, normalized to hprt1 and control, 24 fish pooled per sample). dpf, days postfertilization.
Extending exposure time of 10 μM 6-OHDA to 48 h cannot produce a PD-like state. To determine if a longer exposure with a lower dose might alter expression, we assessed a 48-h treatment at 10 μM 6-OHDA. Each point represents pooled RNA harvested from at least 15 fish from 3 different cohorts. (Strain AB, normalized to hprt1 and control). No significant expression differences were identified compared to control. PD, Parkinson's disease.
Behavioral assessment
Clinically, Parkinson's is defined by behavioral classifiers, and other zebrafish groups have demonstrated that locomotor changes are present following 6-OHDA exposure; therefore, we assessed the behavior of morphologically normal larvae that demonstrated a reduction in th1 expression. Blinded assessment of 6 dpf larvae exposed to 30 μM 6-OHDA for 24 h beginning at 3 dpf demonstrated a significant reduction in the number of animals that moved over a 10-min period compared to controls (Fig. 5, p = 0.04, n = 35). Subsequent analysis was performed on the morphologically normal animals that did move (21 controls and 13 exposed), and no significant differences were observed for distance (p = 0.9), time moved (p = 0.8), velocity (p = 0.8), or latency to first movement (p = 0.76).
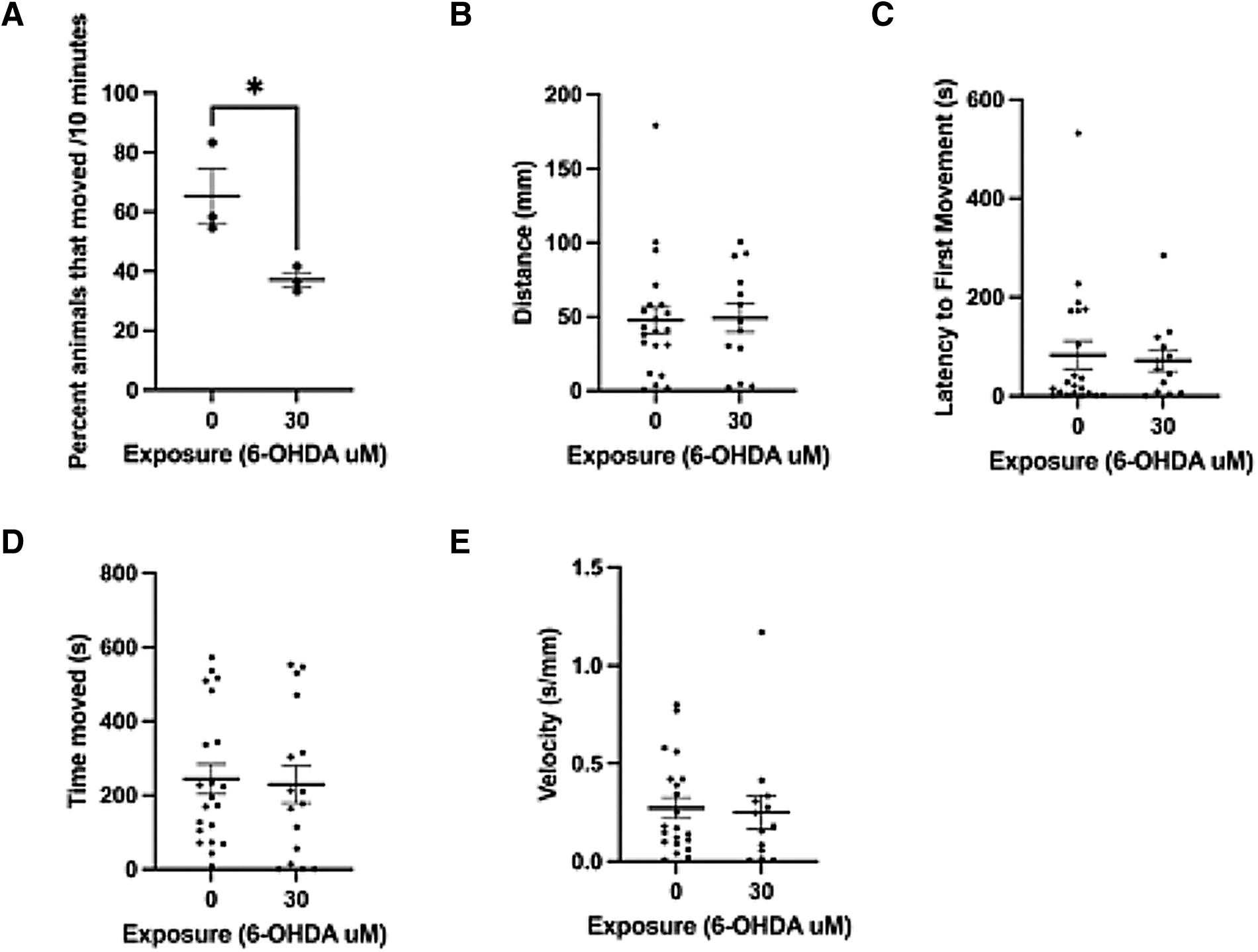
6-OHDA exposure decreases larval movement. A detailed analysis was performed of the movement of 6 dpf larval zebrafish following a 24-h treatment starting at 3 dpf with 30 μM 6-OHDA. Blinded analysis of whether an animal moved over 10 min was first performed
Discussion
Previous attempts to utilize 6-OHDA in zebrafish had shortcomings. In 2014, Feng and coworkers published the initial characterization of this technique. It focused on analysis with 250 μM 6-OHDA; however, lacked the use of L-ascorbic acid that is critical for maintaining 6-OHDA activity in aqueous environments. This protocol has been repeated by others 16 ; however, in our hands, we observed high lethality and morphological issues precluding the use of this protocol in our laboratory. We reviewed other protocols and identified protocols that ranged from Kalyn and coworkers that stated toxicity issues precluded 6-OHDA use at 1 μM 6-OHDA to Vaz and coworkers that utilized 750 μM for large-scale drug screening (Table 1).3,14 The protocols where L-ascorbic acid concentrations varied 3 or was absent 4 would cause a significant difference on variability, potential side effects, and required concentrations. Similar to Vaz and coworkers, we utilized HEPES-buffered embryo water to avoid water acidification that can be a result of L-ascorbic acid use. The HEPES buffer maintains the 0.2% L-ascorbic acid containing embryo water at a pH of 7. Even with our best vehicle (containing 40 mM HEPES buffer, 0.2% L-ascorbic acid, and methylene blue), we could not obtain results that used high concentrations of 6-OHDA similar to other protocols4,14 due to lack of viability. In addition, the morphological defects that we observed at concentrations greater than 70 μM 6-OHDA in the surviving embryos would make behavioral analysis difficult. In other protocols, these animals have been used for analyses including behavioral analysis 16 ; however, we question how to disentangle the movement defects due to morphological differences from those directly attributed to DA neurodegeneration. We excluded larvae with morphological deformations from our behavioral analyses, and still observed a reduction in animals that moved. Furthermore, our methodology was quick and focused on a movement versus no movement assessment. It eliminated the need for special recording and behavioral analysis approaches making it faster for screening and more accessible to laboratories reducing initial investment. 17
Our protocol utilized RT-qPCR to identify a quantitative reduction of th1 rather than initially focusing on behavioral approaches. This was chosen for two reasons: (1) RT-qPCR provided a rapid analytical method with less variability than behavior—speeding up our analysis, and (2) by looking directly at th1 reduction, we could avoid characterization of nonspecific or toxic side effects that could contribute to behavioral deficits. The reduction of th1 was reproducible with our final protocol.
The markers slc6a3 and slc18a2 have been utilized in conjunction with th1 to triangulate the condition of mDA neurons. We did not see a similar reduction in these markers. There are several reasons that could contribute to this difference. The first is RNA quality leading to variability. For the slc18a2 marker, we performed a leave one out experiment, and without the oldest slc18a2 sample, we do see a significant reduction (data not shown). The second reason may be that the DA neurons may be injured leading to a reduction in th1 compared to controls; however, the neurons are not completely degenerated. Lastly, we utilized whole pooled larvae for RNA collection. The specific effects are potentially diminished due to their expression in non-mDA cells. Lastly, we propose that this effect is specific as it did not change expression of gad1b indicating that it is not impacting other neuronal populations. Whether the 6-OHDA has induced specific mDA neurodegeneration or induced a disease state, the net result of having less th1 would be a reduced DA availability, suggesting a specific phenotype for assessing potential treatments. Future work will be focused on confirmation with protein assays.
We expanded our analysis by assessing additional outputs of pink1 and sod2 that suggest the 6-OHDA is affecting mitochondria and upregulating the superoxide dismutase system to compensate.5,18,19 This is consistent with the research in other 6-OHDA models with regards to mechanism. While this may be considered an obvious outcome, we feel it is important to demonstrate that 6-OHDA was working as expected. This work provides a starting point to investigate whether mDA neurons either have a reduced ability to respond to ROS or more likely have an equivalent ability to respond, but due to additional ROS production inherent to their function have a reduced threshold before pathogenesis.20,21
Lastly, our protocol utilized an acute exposure followed by multiple days in vehicle. This allows time for neurodegenerative processes to progress, or compensatory cellular processes to occur. This opens up two potential avenues of research. First, utilizing an expanded molecular fingerprint, the window between 3 dpf (initiation of exposure) and 6 dpf to characterize cellular changes associated with 6-OHDA neurodegeneration. This could lead to a better understanding of the processes of cell death associated with increased ROS. 22 Second, this provides the opportunity for assessment of novel compounds designed for the treatment of diseases associated with mDA loss, including PD. The combination of the molecular fingerprint with an easily assessed behavioral phenotype can be used to assess neuroprotective compounds. We acknowledge that there are still limitations with the 6-OHDA model, as it only models one aspect of PD—the DA neuron loss; however, combined with other models for example, the synuclein aggregation model, 23 it will give us a better understanding of what prospective compounds are doing at the molecular level.
Footnotes
Acknowledgments
We would like to thank Dr. Mattew Greenwold (UT Tyler) for his advice on RT-qPCR analysis and Dr. Randy Back (UT Tyler) for the use of his recording setup. AB strain Zebrafish provided by the Zebrafish International Resource Center (ZIRC) at the University of Oregon This work was supported by the NIH R16 (1R16NS129676-01) and the Texas Academy of Science (Student Research Award Competition M.S.) Material was presented previously as a Master of Science Dissertation—“Modeling a Parkinson's Disease-Like State in Zebrafish Larvae Utilizing 6-Hydroxydopamine,” Adrian Romero, M.S., University of Texas at Tyler, December 2023, ![]() .
.
Authors' Contributions
A.R.: Conceptualization, Methodology, Formal Analysis, Investigation, Writing—original draft, Writing—review and editing, Funding Acquisition; A.S.: Investigation; J.D.J.: Investigation; K.L.: Investigation; M.S.E-.H.: Methodology, Supervision, A.K.H.: Conceptualization, Methodology, Formal Analysis, Writing—original draft, Writing—review and editing, Supervision, Project Administration, Funding Acquisition; B.R.B.; Conceptualization, Methodology, Formal Analysis, Investigation, Writing—original draft, Writing—review and editing, Supervision, Project Administration, Funding Acquisition.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work is funded by the National Institute of Neurological Disorders and Stroke: R16NS129676 (PI-Hamouda), Texas Academy of Sciences (PI-Romero).